

Protein Phosphatases MoPtc5, MoPtc1, and MoPtc2 Contribute to the Vegetative Growth, Stress Adaptation, and Virulence of Magnaporthe oryzae

 , , ,
, , ,  , ,
, ,  , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Genes Knockout and Complementation Assays
2.2. Fungal Strains, Media and Culture Conditions
2.3. Pathogenicity, Host Penetration, and Analysis of Appressoria Turgor Pressure
2.4. RNA Extraction, Quantitative Real-Time PCR (qRT-PCR), and RT-PCR Assays
2.5. Southern Blotting Assay
2.6. Extraction of Proteins and Western Blotting Assays
2.7. Microscopic Examination Assays
2.8. Statistical Analysis
3. Results
3.1. Generation of ∆Moptc5∆Moptc1 and ∆Moptc5∆Moptc2 Deletion Mutants
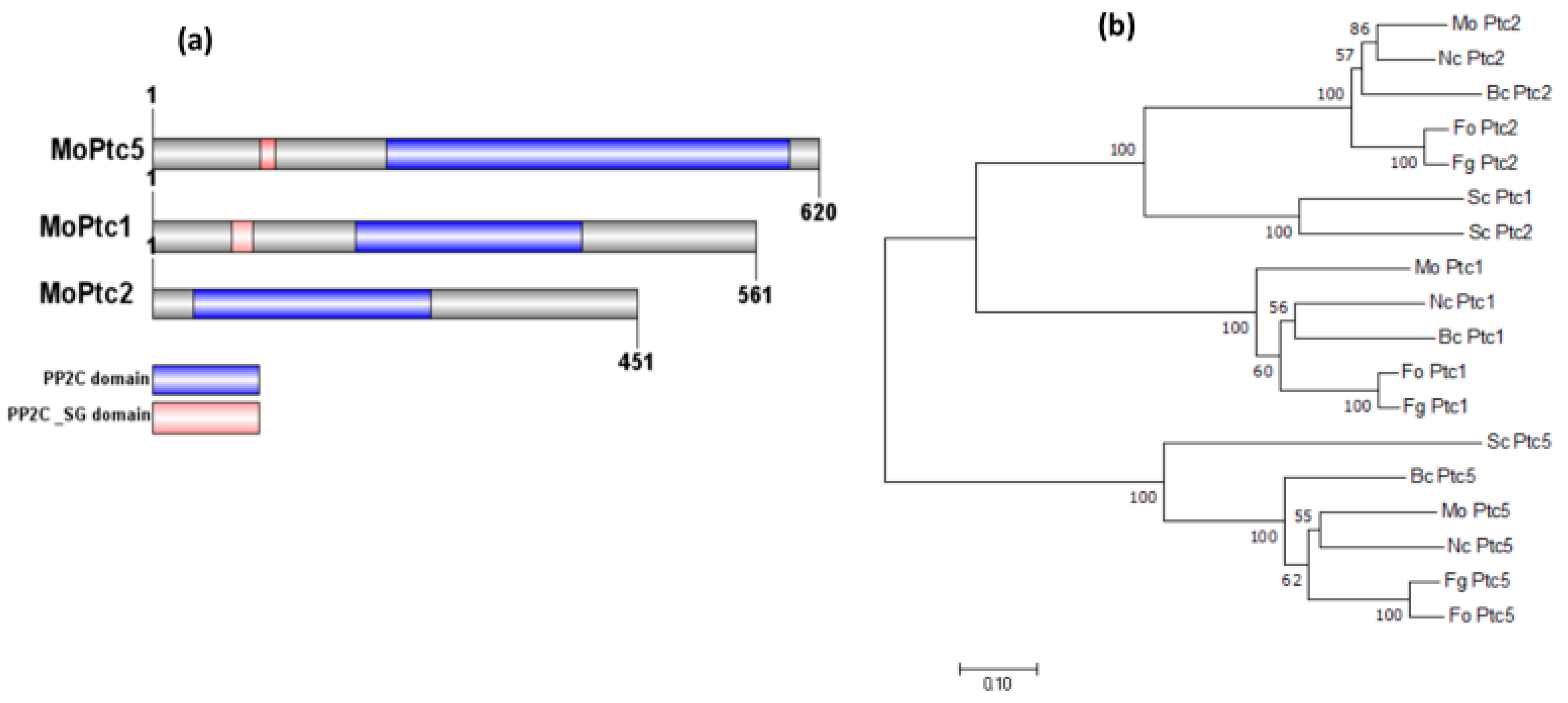
3.2. Phylogenetic Analyses and Identification of MoPtc1, MoPtc2, and MoPtc5
3.3. ∆Moptc5∆Moptc1 and ∆Moptc5∆Moptc2 Strongly Affect the Hyphal Growth and Sporulation of M. oryzae
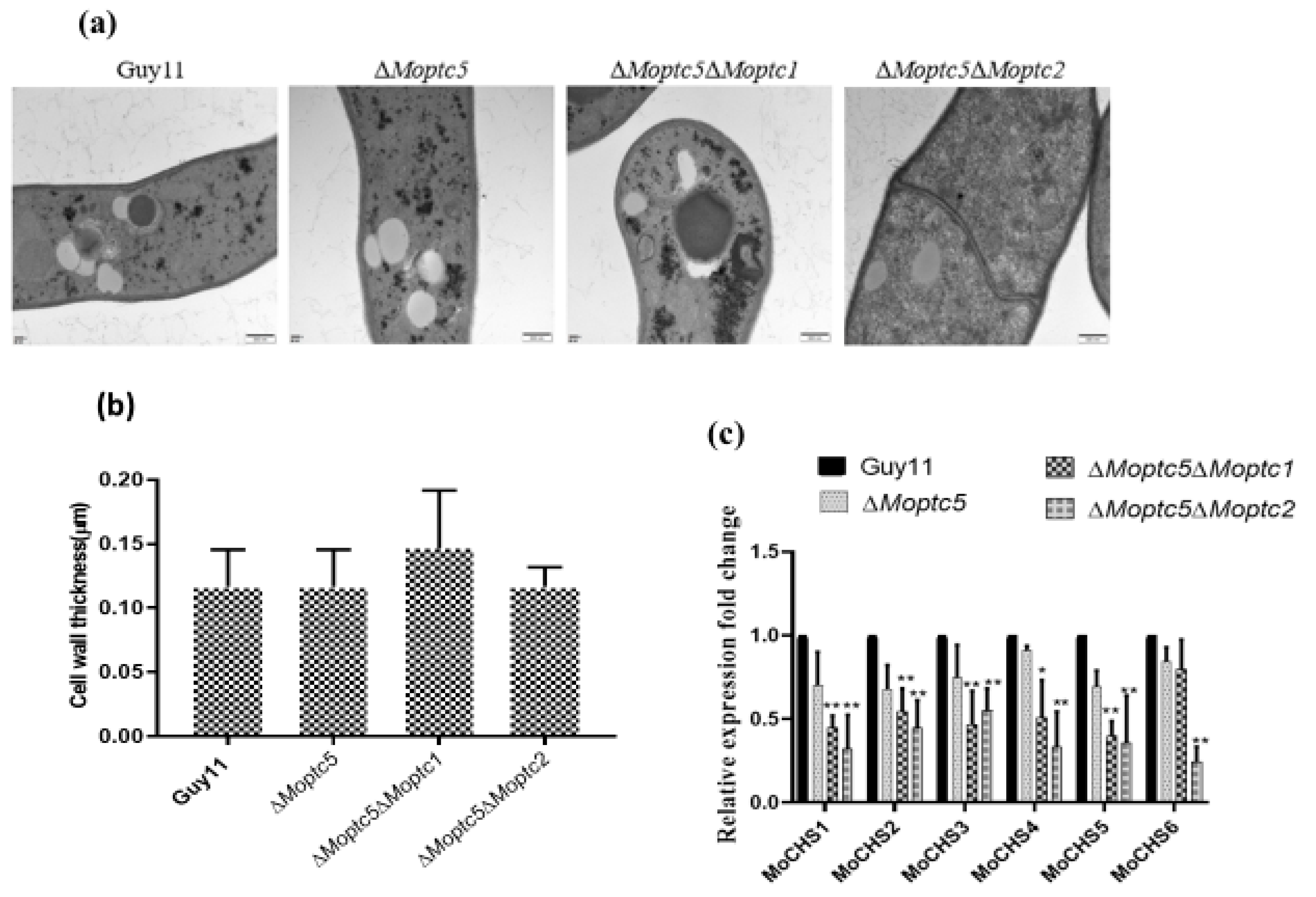
3.4. MoPTC5, MoPTC1, and MoPTC2 in Cell Wall Maintenance and Mps1 Phosphorylation Level
3.5. Double Knockout of MoPTC5 with MoPTC1 and MoPTC2 Did Not Influence the Cell Wall Thickness of Rice Blast Fungus
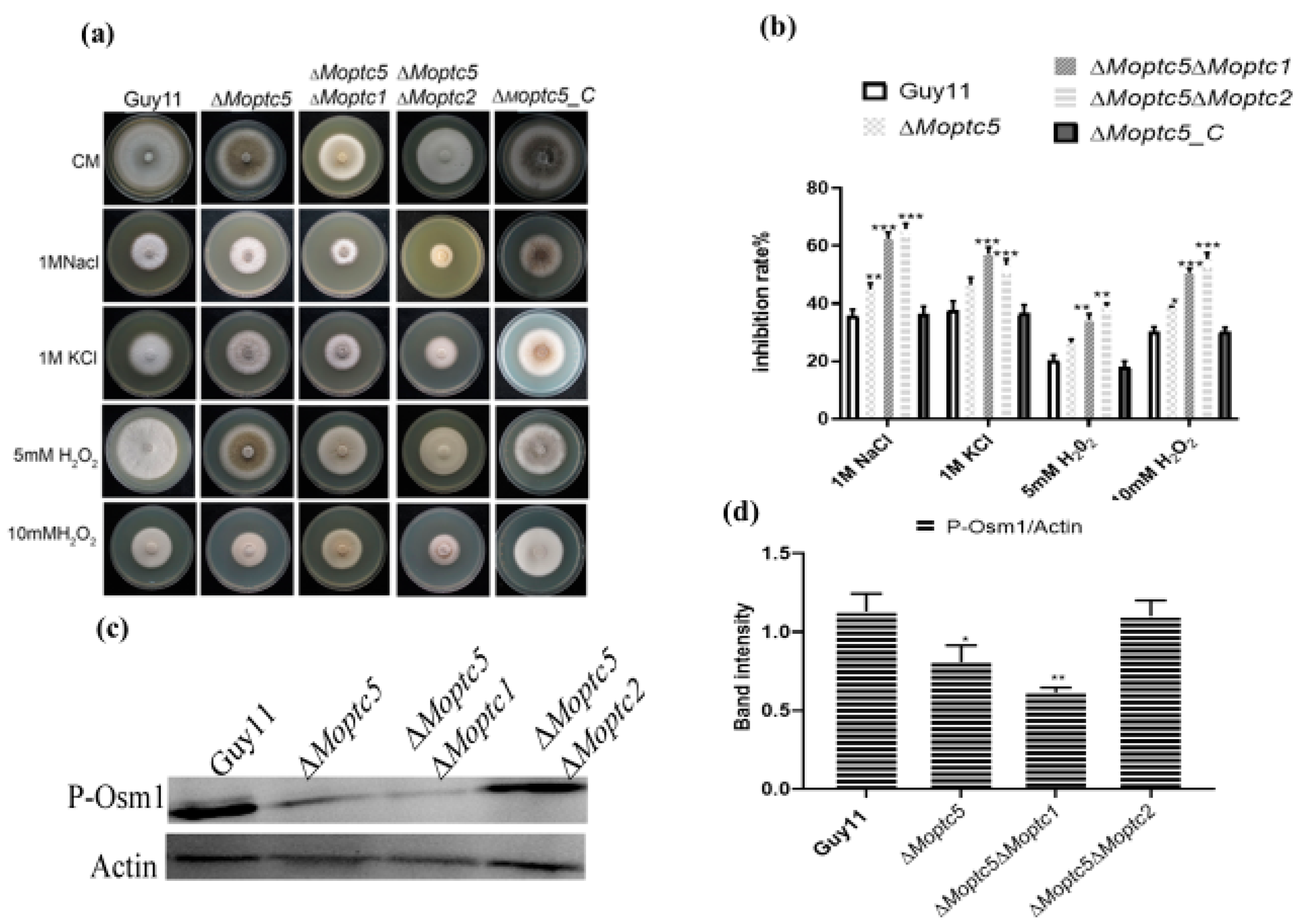
3.6. Moptc5, Along with Moptc1 and Moptc2, Plays Important Roles in Osmotic, Oxidative Stress, and MoOsm1 Phosphorylation in M. oryzae
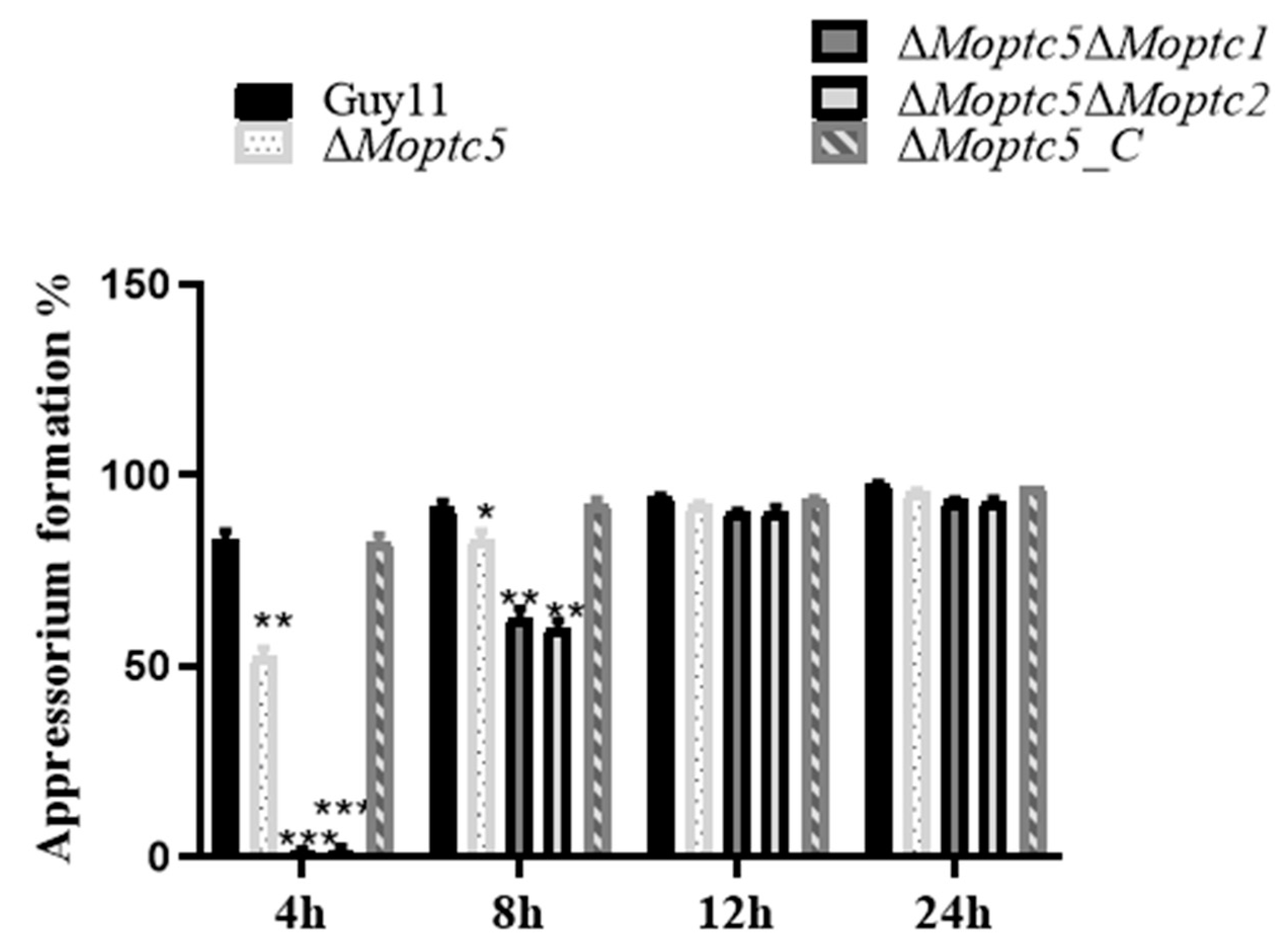
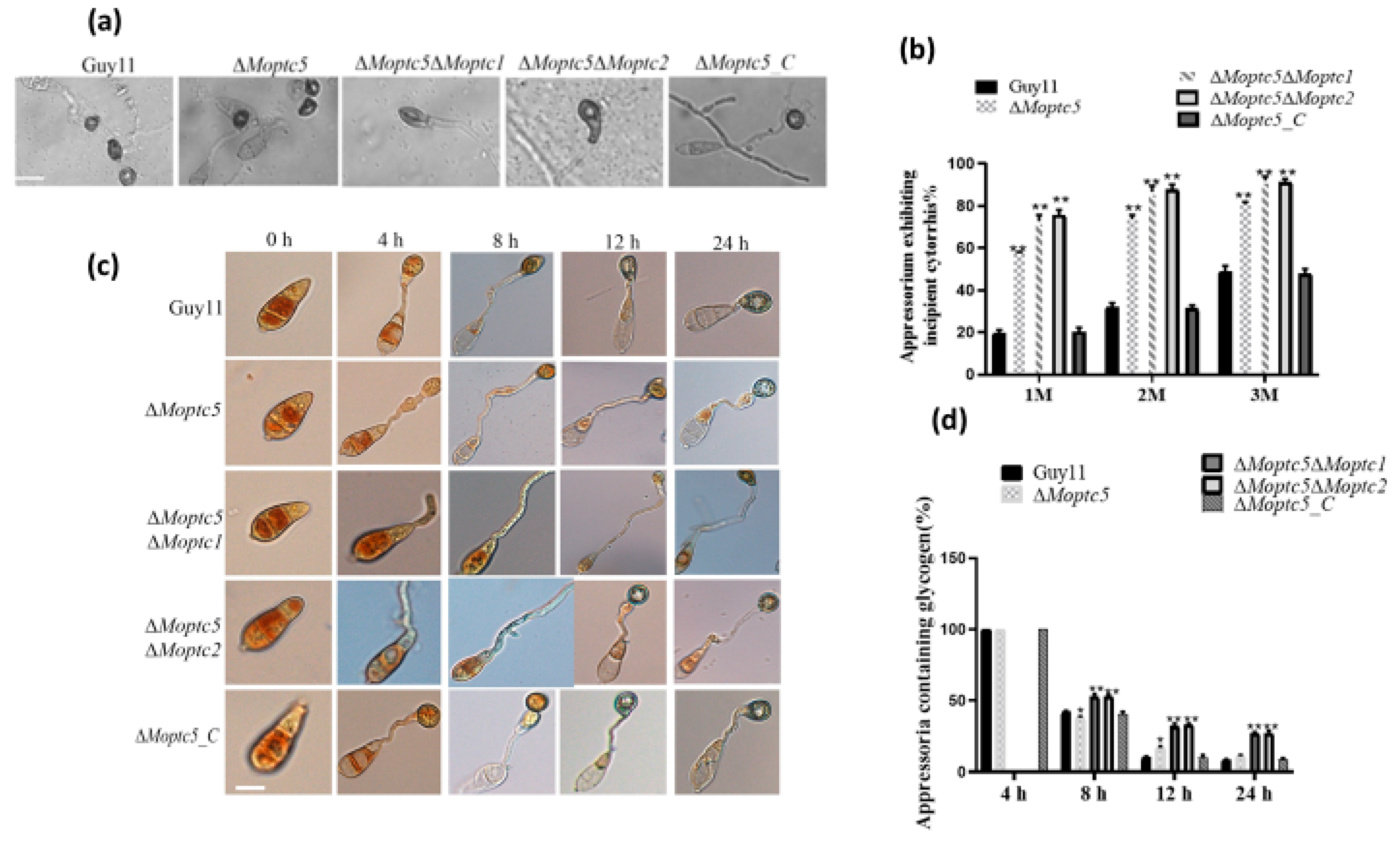
3.7. ∆Moptc5∆Moptc1 and ∆Moptc5∆Moptc2 Mutants Are Remarkably Delayed in Appressoria Formation
3.8. MoPTC5, MoPTC1, and MoPTC2 Synergistically Influence Appressoria Turgor Generation and the Utilization of Glycogen in M. oryzae
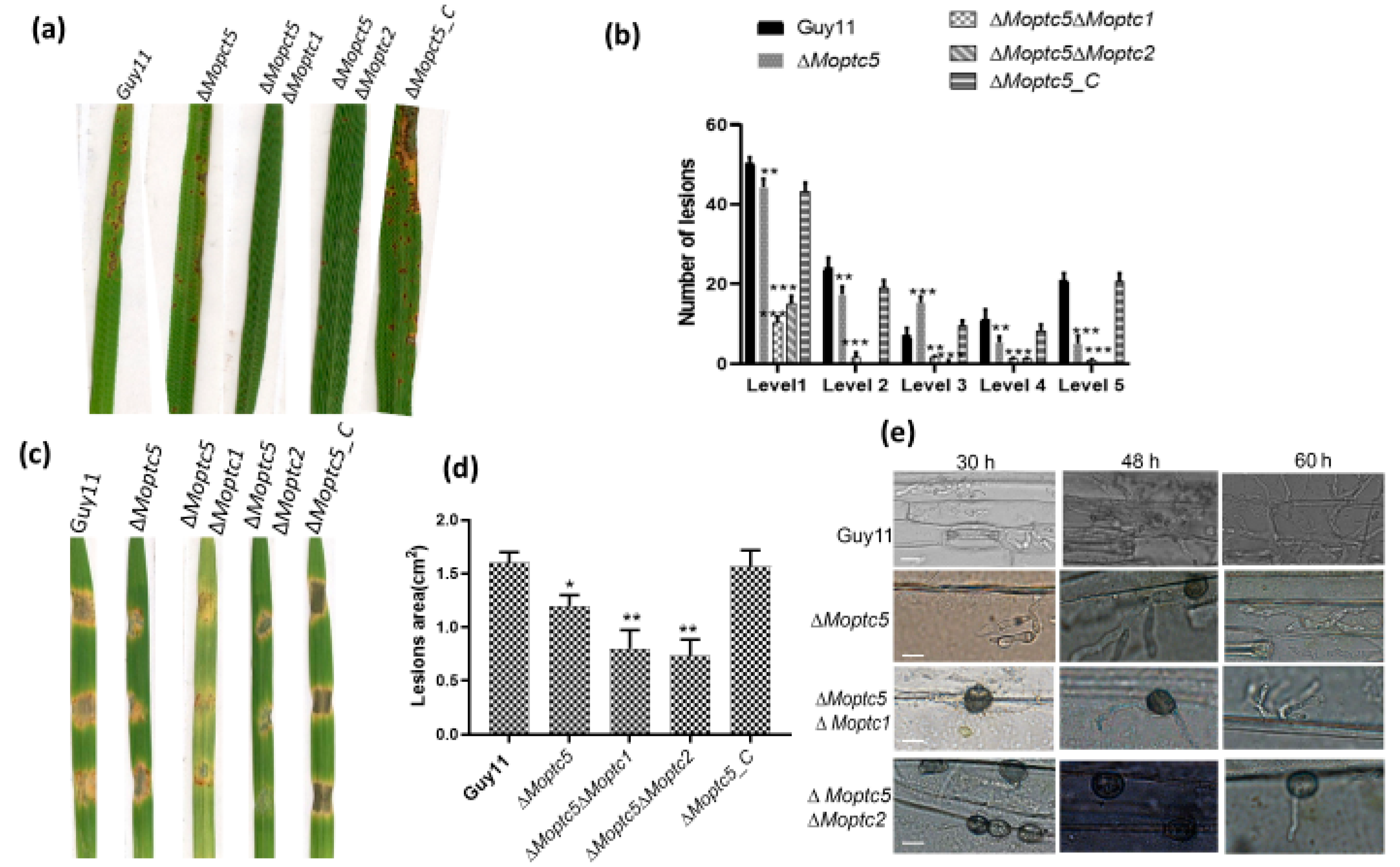
3.9. ∆Moptc5∆Moptc1 and ∆Moptc5∆Moptc2 Strains Weakened Virulence to Infect Rice Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fernandez, J.; Orth, K. Rise of a Cereal Killer: The Biology of Magnaporthe oryzae Biotrophic Growth. Trends Microbiol. 2018, 26, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Goh, J.; Han, S.S.; Roh, J.H.; Kim, Y.; Heu, S.; Shim, H.K.; Jeong, D.G.; Kang, I.J.; Yang, J.W. Comparative Pathogenicity and Host Ranges of Magnaporthe oryzae and Related Species. Plant Pathol. 2020, 36, 305–313. [Google Scholar] [CrossRef]
- Li, J.; Lu, L.; Wang, Q.; Shi, Z.; Li, C.; Guo, Z. Genome Re-Sequencing Reveals the Host-Specific Origin of Genetic Variation in Magnaporthe Species. Front. Genet. 2022, 13, 861727. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, N.; Ganesan, P.; Mughal, N.M.; Yadav, I.S.; Kumar, A. Genome assisted molecular typing and pathotyping of rice blast pathogen, Magnaporthe oryzae, reveals a genetically homogenous population with high virulence diversity. Fungal Biol. 2021, 125, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Wengler, M.R.; Talbot, N.J. Mechanisms of regulated cell death during plant infection by the rice blast fungus Magnaporthe oryzae. Cell Death Differ. 2025, 10, 1038. [Google Scholar] [CrossRef]
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two distinct secretion systems facilitate tissue invasion by the rice blast fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996. [Google Scholar] [CrossRef]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef]
- Gupta, Y.K.; Dagdas, Y.F.; Martinez-Rocha, A.-L.; Kershaw, M.J.; Littlejohn, G.R.; Ryder, L.S.; Sklenar, J.; Menke, F.; Talbot, N.J. Septin-dependent assembly of the exocyst is essential for plant infection by Magnaporthe oryzae. Plant Cell 2015, 27, 3277–3289. [Google Scholar] [CrossRef]
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Rice blast: A disease with implications for global food security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef]
- Yamaguchi, I. Overview on the chemical control of rice blast disease. In Rice Blast: Interaction with Rice and Control, Proceedings of the 3rd International Rice Blast Conference, Ibaraka, Japan, January 2004; Springer: Berlin/Heiderberg, Germany, 2004; pp. 1–13. [Google Scholar]
- Simkhada, K.; Thapa, R. Rice blast, a major threat to the rice production and its various management techniques. Turk. J. Agric.-Food Sci. Technol 2022, 10, 147–157. [Google Scholar] [CrossRef]
- Sahu, P.K.; Sao, R.; Choudhary, D.K.; Thada, A.; Kumar, V.; Mondal, S.; Das, B.K.; Jankuloski, L.; Sharma, D. Advancement in the breeding, biotechnological and genomic tools towards development of durable genetic resistance against the rice blast disease. Plants 2022, 11, 2386. [Google Scholar] [CrossRef]
- Ariño, J.; Velázquez, D.; Casamayor, A. Ser/Thr protein phosphatases in fungi: Structure, regulation and function. Microb Cell 2019, 6, 217. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Sugimoto, H.; Inoue, S.I.; Takahashi, Y.; Hayashi, M.; Hayashi, Y.; Mizutani, M.; Ogawa, T.; Kinoshita, D.; Ando, E.; et al. Type 2C protein phosphatase clade D family members dephosphorylate guard cell plasma membrane H+-ATPase. Plant Physiol. 2022, 188, 2228–2240. [Google Scholar] [CrossRef]
- Ghosh, A.; Servin, J.A.; Park, G.; Borkovich, K.A. Global analysis of serine/threonine and tyrosine protein phosphatase catalytic subunit genes in Neurospora crassa reveals interplay between phosphatases and the p38 mitogen-activated protein kinase. Genes Genome Genet. 2014, 4, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Ariño, J.; Casamayor, A.; González, A. Type 2C protein phosphatases in fungi. Eukaryot. Cell 2011, 10, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Lemos, P.; Ruiz-Roldán, C.; Hera, C. Role of the phosphatase Ptc1 in stress responses mediated by CWI and HOG pathways in Fusarium oxysporum. Fungal Genet. Biol. 2018, 118, 10–20. [Google Scholar] [CrossRef]
- Tatjer, L.; González, A.; Serra-Cardona, A.; Barceló, A.; Casamayor, A.; Ariño, J. The Saccharomyces cerevisiae Ptc1 protein phosphatase attenuates G2-M cell cycle blockage caused by activation of the cell wall integrity pathway. Mol. Microbiol. 2016, 101, 671–687. [Google Scholar] [CrossRef]
- Peng, Y.J.; Ding, J.L.; Feng, M.G.; Ying, S.H. Glc8, a regulator of protein phosphatase type 1, mediates oxidation tolerance, asexual development and virulence in Beauveria bassiana, a filamentous entomopathogenic fungus. Curr. Genet. 2019, 65, 283–291. [Google Scholar] [CrossRef]
- Rodriguez-Iglesias, A.; Schmoll, M. Protein phosphatases regulate growth, development, cellulases and secondary metabolism in Trichoderma reesei. Sci. Rep. 2019, 9, 10995. [Google Scholar] [CrossRef]
- Jenkinson, F.; Zegerman, P. Roles of phosphatases in eukaryotic DNA replication initiation control. DNA Repair 2022, 118, 103384. [Google Scholar] [CrossRef]
- Fasullo, M. Checkpoint Control of DNA Repair in Yeast. In Saccharomyces; IntechOpen: London, UK, 2021. [Google Scholar]
- Cai, Y.; Liu, X.; Shen, L.; Wang, N.; He, Y.; Zhang, H.; Wang, P.; Zhang, Z. Homeostasis of cell wall integrity pathway phosphorylation is required for the growth and pathogenicity of Magnaporthe oryzae. Mol. Plant Pathol. 2022, 23, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Anjago, W.M.; Biregeya, J.; Shi, M.; Chen, Y.; Wang, Y.; Wang, Z.; Hong, Y.; Chen, M. The Calcium Chloride Responsive Type 2C Protein Phosphatases Play Synergistic Roles in Regulating MAPK Pathways in Magnaporthe oryzae. J. Fungi 2022, 8, 1287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, M.; Yang, G.; Zhang, B.; Cao, X.; Yuan, J.; Ge, F.; Wang, S. PP2C phosphatases Ptc1 and Ptc2 dephosphorylate PGK1 to regulate autophagy and aflatoxin synthesis in the pathogenic fungus Aspergillus flavus. mBio 2023, 14, e00977-23. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, J.; Li, J.; Jiang, L. Mithochondrial type 2C protein phosphatases CaPtc5p, CaPtc6p, and CaPtc7p play vital roles in cellular responses to antifungal drugs and cadmium in Candida albicans. FEMS Yeast Res. 2012, 12, 897–906. [Google Scholar] [CrossRef]
- Sharmin, D.; Sasano, Y.; Sugiyama, M.; Harashima, S. Effects of deletion of different PP2C protein phosphatase genes on stress responses in Saccharomyces cerevisiae. Yeast 2014, 31, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Biregeya, J.; Anjago, W.M.; Pan, S.; Zhang, R.; Yang, Z.; Chen, M.; Felix, A.; Xu, H.; Lin, Y.; Nkurikiyimfura, O. Type 2C Protein Phosphatases MoPtc5 and MoPtc7 Are Crucial for Multiple Stress Tolerance, Conidiogenesis and Pathogenesis of Magnaporthe oryzae. J. Fungi 2022, 9, 1. [Google Scholar] [CrossRef]
- Kim, J.-A.; Hicks, W.M.; Li, J.; Tay, S.Y.; Haber, J.E. Protein phosphatases pph3, ptc2, and ptc3 play redundant roles in DNA double-strand break repair by homologous recombination. Mol. Cell. Biol. 2011, 31, 507–516. [Google Scholar] [CrossRef]
- Shearer, C.; Crane, J.; Chen, W. Freshwater ascomycetes: Ophioceras species. Mycologia 1999, 91, 145–156. [Google Scholar] [CrossRef]
- Huhndorf, S.; Miller, A.N. A new species of Camarops and phylogenetic analysis of related taxa in the Boliniaceae. N. Am. Fungi 2008, 3, 231–239. [Google Scholar] [CrossRef]
- Matar, K.A.O.; Chen, X.; Chen, D.; Anjago, W.M.; Norvienyeku, J.; Lin, Y.; Chen, M.; Wang, Z.; Ebbole, D.J.; Lu, G.-d. WD40-repeat protein MoCreC is essential for carbon repression and is involved in conidiation, growth and pathogenicity of Magnaporthe oryzae. Curr. Genet. 2017, 63, 685–696. [Google Scholar] [CrossRef]
- Zhou, R.Q.; Jin, J.J.; Li, Q.M.; Su, Z.Z.; Yu, X.J.; Tang, Y.; Luo, S.M.; He, Y.; Li, X.L. Early Detection of Magnaporthe oryzae-Infected Barley Leaves and Lesion Visualization Based on Hyperspectral Imaging. Front. Plant Sci. 2018, 9, 1962. [Google Scholar] [CrossRef]
- Lin, L.; Cao, J.; Du, A.; An, Q.; Chen, X.; Yuan, S.; Batool, W.; Shabbir, A.; Zhang, D.; Wang, Z.; et al. eIF3k Domain-Containing Protein Regulates Conidiogenesis, Appressorium Turgor, Virulence, Stress Tolerance, and Physiological and Pathogenic Development of Magnaporthe oryzae. Front. Plant Sci. 2021, 12, 748120. [Google Scholar]
- Huhndorf, S.M.; Greif, M.; Mugambi, G.K.; Miller, A.N. Two new genera in the Magnaporthaceae, a new addition to Ceratosphaeria and two new species of Lentomitella. Mycologia 2008, 100, 940–955. [Google Scholar] [CrossRef] [PubMed]
- Siddiquee, S.; Rovina, K.; Naher, L.; Rodrigues, K.F.; Uzzaman, M.A. Phylogenetic relationships of Termitomyces aurantiacus inferred from internal transcribed spacers DNA sequences. Adv. Biosci. Biotechnol. 2015, 6, 358. [Google Scholar] [CrossRef]
- Molina, A.; Jordá, L.; Torres, M.; Martín-Dacal, M.; Berlanga, D.J.; Fernández-Calvo, P.; Gómez-Rubio, E.; Martín-Santamaría, S. Plant cell wall-mediated disease resistance: Current understanding and future perspectives. Mol. Plant 2024, 17, 699–724. [Google Scholar] [CrossRef]
- Cruz-Mireles, N.; Osés-Ruiz, M.; Derbyshire, P.; Jégousse, C.; Ryder, L.S.; Bautista, M.J.A.; Eseola, A.; Sklenar, J.; Tang, B.; Yan, X.; et al. The phosphorylation landscape of infection-related development by the rice blast fungus. Cell 2024, 187, 2557–2573.e18. [Google Scholar] [CrossRef]
- Li, C.; Xu, D.; Hu, M.; Zhang, Q.; Xia, Y.; Jin, K. MaNCP1, a C2H2 Zinc Finger Protein, Governs the Conidiation Pattern Shift through Regulating the Reductive Pathway for Nitric Oxide Synthesis in the Filamentous Fungus Metarhizium acridum. Microbiol. Spectr. 2022, 10, e0053822. [Google Scholar] [CrossRef]
- Jung, B.; Kim, S.; Lee, J. Microcyle conidiation in filamentous fungi. Mycobiology 2014, 42, 1–5. [Google Scholar] [CrossRef]
- Kim, K.S.; Lee, Y.-H. Gene expression profiling during conidiation in the rice blast pathogen Magnaporthe oryzae. PLoS ONE 2012, 7, e43202. [Google Scholar] [CrossRef]
- Pogorelko, G.; Lionetti, V.; Bellincampi, D.; Zabotina, O. Cell wall integrity: Targeted post-synthetic modifications to reveal its role in plant growth and defense against pathogens. Plant Signal. Behav. 2013, 8, e25435. [Google Scholar] [CrossRef]
- Pusztahelyi, T. Chitin and chitin-related compounds in plant–fungal interactions. Mycology 2018, 9, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Goughenour, K.D.; Whalin, J.; Slot, J.C.; Rappleye, C.A. Diversification of Fungal Chitinases and Their Functional Differentiation in Histoplasma capsulatum. Mol. Biol. Evol. 2021, 38, 1. [Google Scholar] [CrossRef] [PubMed]
- Özer Uyar, G.E.; Uyar, b. effect of nacl and kcl salts on growth and lactic acid production of Rhizopus oryzae. GIDA J. Food 2016, 41, 299. [Google Scholar]
- Wang, Y.; Guan, Y.; Lin, W.; Yan, H.; Neng, J.; Sun, P. Quantitative Proteomic Profiling of Fungal Growth, Development, and Ochratoxin A Production in Aspergillus ochraceus on High- and Low-NaCl Cultures. Toxins 2021, 13, 51. [Google Scholar] [CrossRef]
- Montibus, M.; Pinson-Gadais, L.; Richard-Forget, F.; Barreau, C.; Ponts, N. Coupling of transcriptional response to oxidative stress and secondary metabolism regulation in filamentous fungi. Crit. Rev. Microbiol. 2015, 41, 295–308. [Google Scholar] [CrossRef]
- Mapuranga, J.; Zhang, N.; Zhang, L.; Chang, J.; Yang, W. Infection strategies and pathogenicity of biotrophic plant fungal pathogens. Front. Microbiol. 2022, 13, 799396. [Google Scholar] [CrossRef]
- Jiang, J.; Yun, Y.; Yang, Q.; Shim, W.-B.; Wang, Z.; Ma, Z. A type 2C protein phosphatase FgPtc3 is involved in cell wall integrity, lipid metabolism, and virulence in Fusarium graminearum. PLoS ONE 2011, 6, e25311. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, P.; Sun, Q.; Li, R.; Qin, Z.; Sha, G.; Zhou, Y.; Bi, R.; Zhang, H.; Zheng, L. The MoPah1 phosphatidate phosphatase is involved in lipid metabolism, development, and pathogenesis in Magnaporthe oryzae. Mol. Plant Pathol. 2022, 23, 720–732. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, J.; Huang, Z.; Li, Y.; Li, H.; Huang, P.; Cai, Y.; Wang, J.; Liu, X.; Lin, F.C.; et al. The endosomal-vacuolar transport system acts as a docking platform for the Pmk1 MAP kinase signaling pathway in Magnaporthe oryzae. New Phytol. 2025, 245, 722–747. [Google Scholar] [CrossRef]
- Aron, O.; Wang, M.; Lin, L.; Batool, W.; Lin, B.; Shabbir, A.; Wang, Z.; Tang, W. MoGLN2 Is Important for Vegetative Growth, Conidiogenesis, Maintenance of Cell Wall Integrity and Pathogenesis of Magnaporthe oryzae. J. Fungi 2021, 7, 463. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, Q.; Li, Y.; Huang, P.; Liao, J.; Wang, J.; Li, H.; Cai, Y.; Wang, J.; Liu, X.; et al. A multilayered regulatory network mediated by protein phosphatase 4 controls carbon catabolite repression and de-repression in Magnaporthe oryzae. Commun. Biol. 2025, 8, 130. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biregeya, J.; Otieno, F.J.; Chen, M.; Mabeche, A.W.; Felix, A.; Aimable, N.; Abubakar, Y.S.; Aron, O.; Lu, G.; Wang, Z.; et al. Protein Phosphatases MoPtc5, MoPtc1, and MoPtc2 Contribute to the Vegetative Growth, Stress Adaptation, and Virulence of Magnaporthe oryzae. J. Fungi 2025, 11, 231. https://doi.org/10.3390/jof11030231
Biregeya J, Otieno FJ, Chen M, Mabeche AW, Felix A, Aimable N, Abubakar YS, Aron O, Lu G, Wang Z, et al. Protein Phosphatases MoPtc5, MoPtc1, and MoPtc2 Contribute to the Vegetative Growth, Stress Adaptation, and Virulence of Magnaporthe oryzae. Journal of Fungi. 2025; 11(3):231. https://doi.org/10.3390/jof11030231
Chicago/Turabian StyleBiregeya, Jules, Frankline Jagero Otieno, Meilian Chen, Anjago Wilfred Mabeche, Abah Felix, Nsanzinshuti Aimable, Yakubu Saddeeq Abubakar, Osakina Aron, Guodong Lu, Zonghua Wang, and et al. 2025. "Protein Phosphatases MoPtc5, MoPtc1, and MoPtc2 Contribute to the Vegetative Growth, Stress Adaptation, and Virulence of Magnaporthe oryzae" Journal of Fungi 11, no. 3: 231. https://doi.org/10.3390/jof11030231
APA StyleBiregeya, J., Otieno, F. J., Chen, M., Mabeche, A. W., Felix, A., Aimable, N., Abubakar, Y. S., Aron, O., Lu, G., Wang, Z., Hong, Y., & Tang, W. (2025). Protein Phosphatases MoPtc5, MoPtc1, and MoPtc2 Contribute to the Vegetative Growth, Stress Adaptation, and Virulence of Magnaporthe oryzae. Journal of Fungi, 11(3), 231. https://doi.org/10.3390/jof11030231