
Procymidone Application Contributes to Multidrug Resistance of Botrytis cinerea

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungicides
2.2. In Vitro Selection of Procymidone-Resistant B. cinerea
2.3. Resistance Level and Stability of Procymidone-Resistant B. cinerea Mutants
2.4. Cross-Resistance
2.5. Mycelial Growth Rate
2.6. Mycelial Growth under Different Temperatures
2.7. Conidial Production of B. cinerea
2.8. Virulence of Procymidone-Resistant Mutants for Host Infection
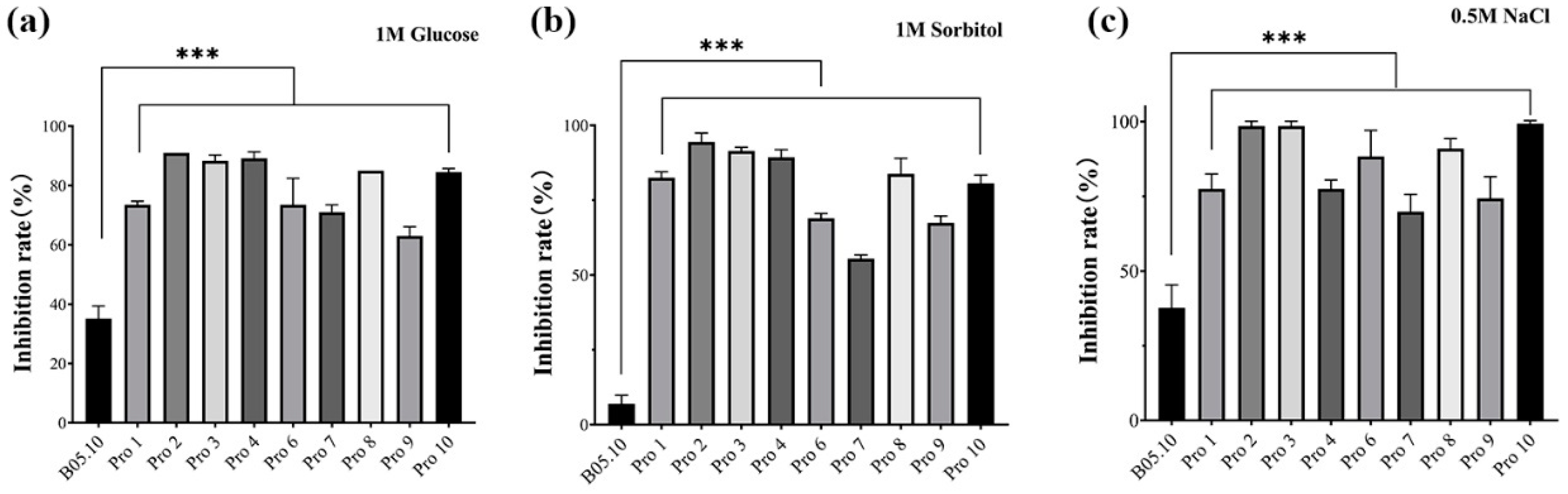
2.9. Sensitivity of B. cinerea Strains to Osmotic Stress
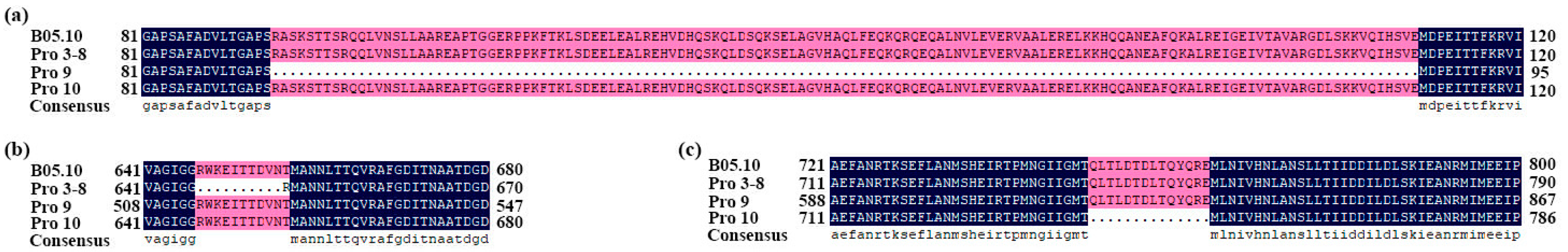
2.10. Sequence Analysis of the BcBos1 Gene and Other Target Genes between Procymidone-Resistant Mutants and Their Wild-Type Parental Strains
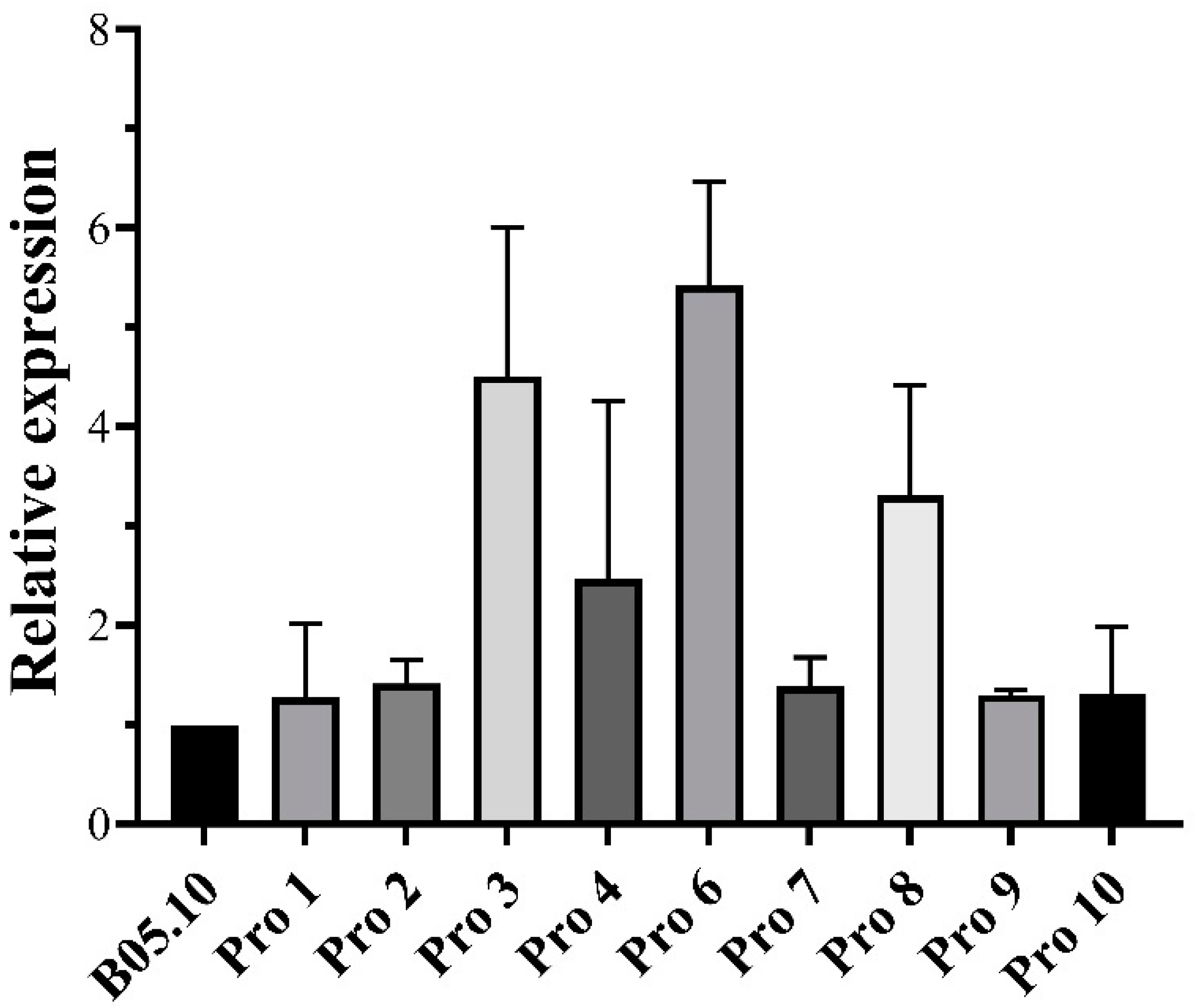
2.11. Comparison of the Expression Level of BcBos1 between Procymidone-Resistant Mutants and Their Wild-Type Parental Strains
2.12. Molecular Docking of Procymidone at B. cinerea BcBos1 Site
2.13. qRT-PCR of ABC Transporter Genes
2.14. Statistical Analyses
3. Results
3.1. Procymidone-Resistant Mutants of B. cinerea
3.2. Cross-Resistance of Procymidone-Resistant Mutants
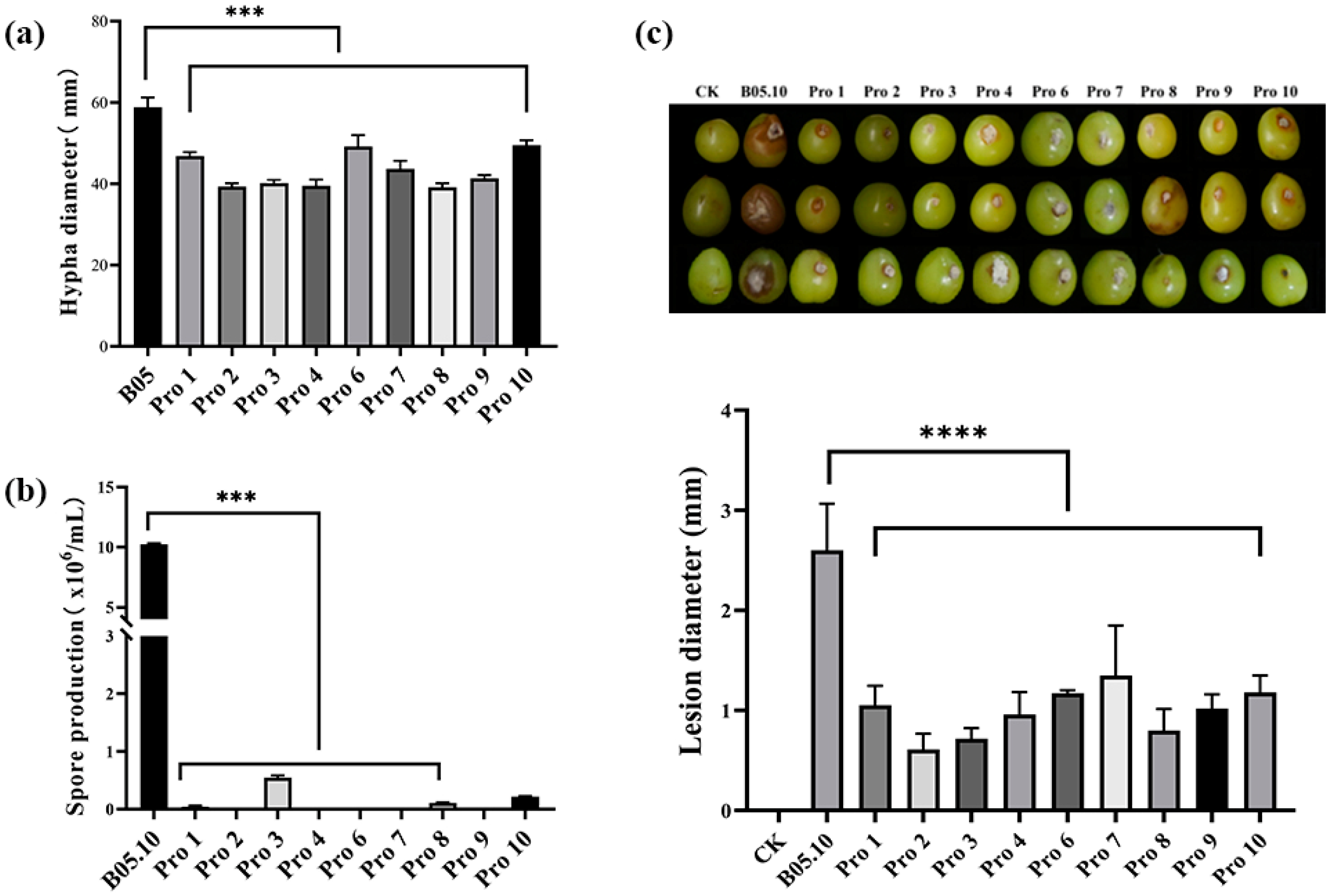
3.3. Fitness of B. cinerea Mutants
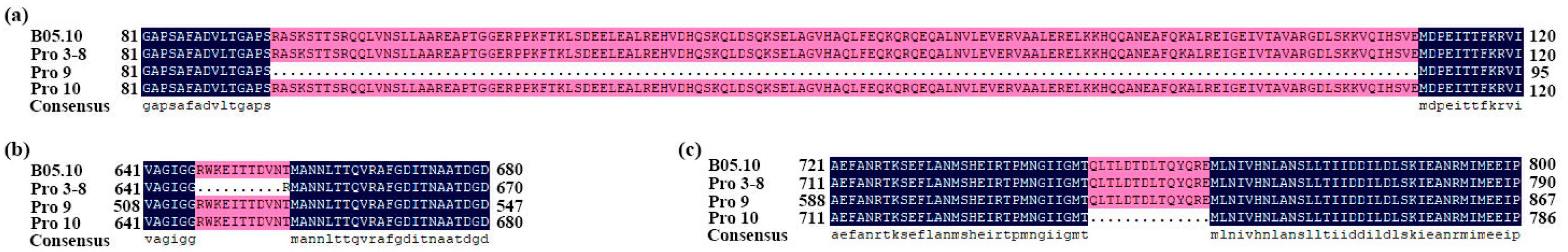
3.4. Sequence Variation in the Target Genes
3.5. BcBos1 Gene Expression Using qRT-PCR
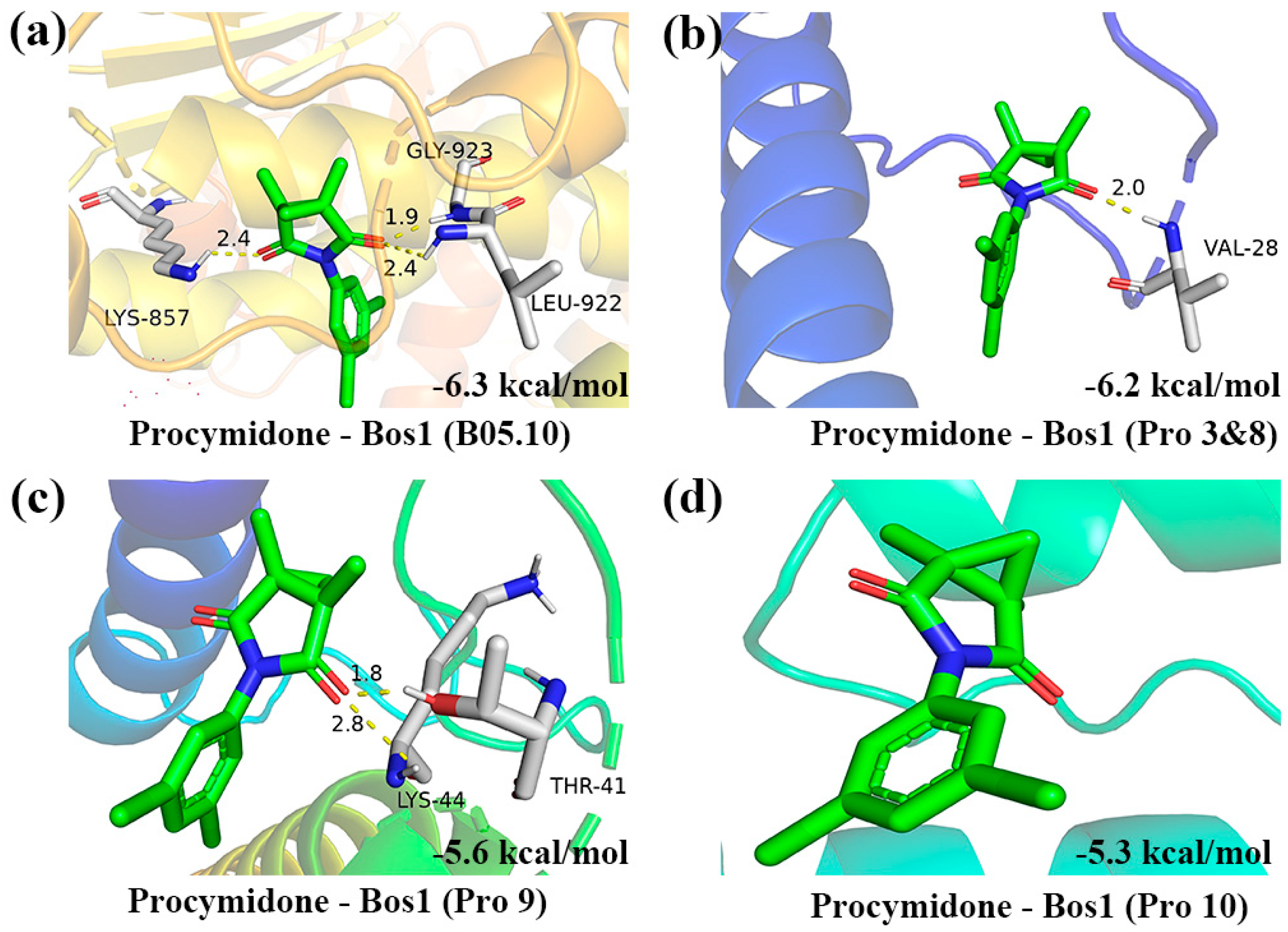
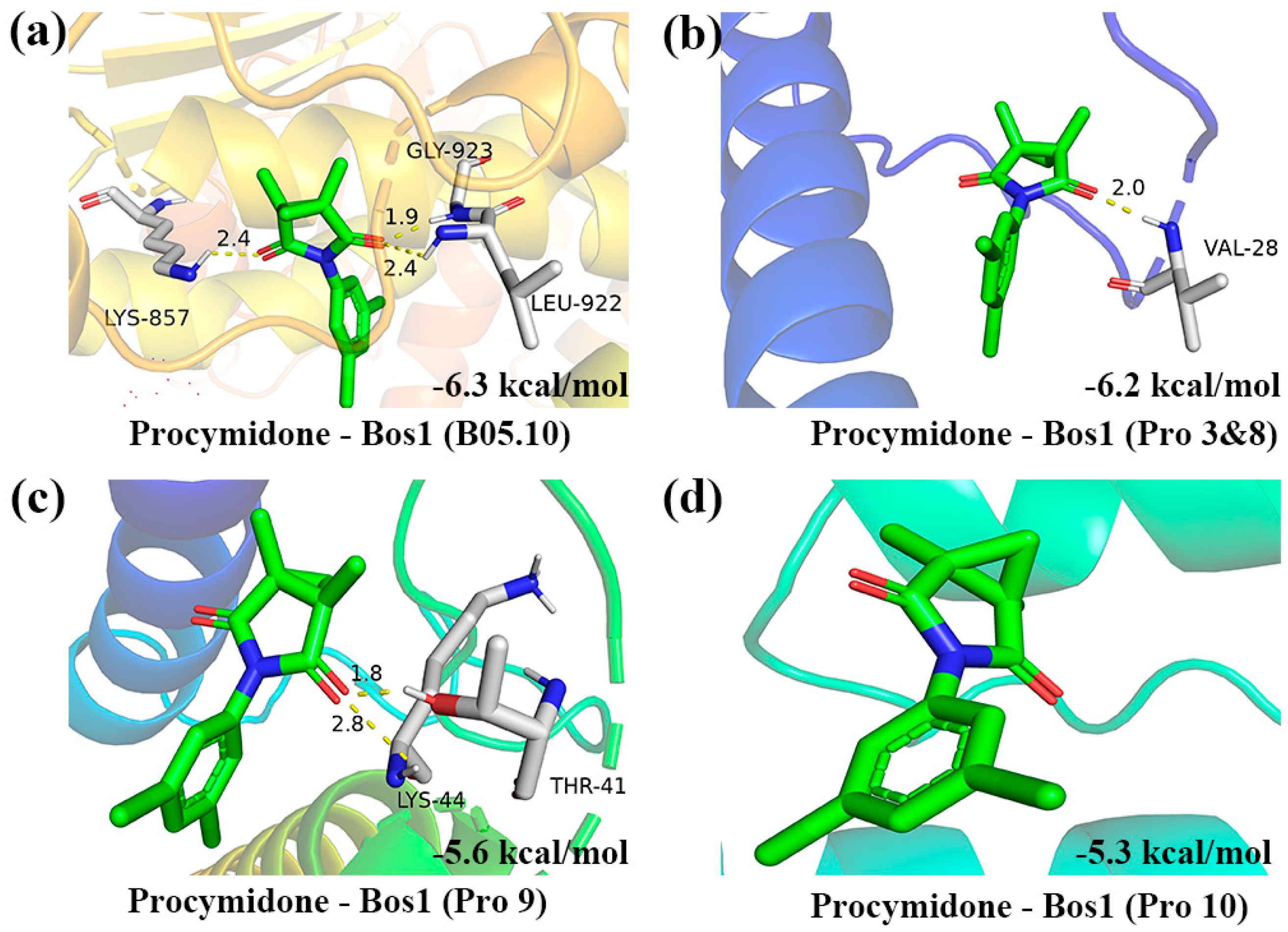
3.6. Molecular Docking
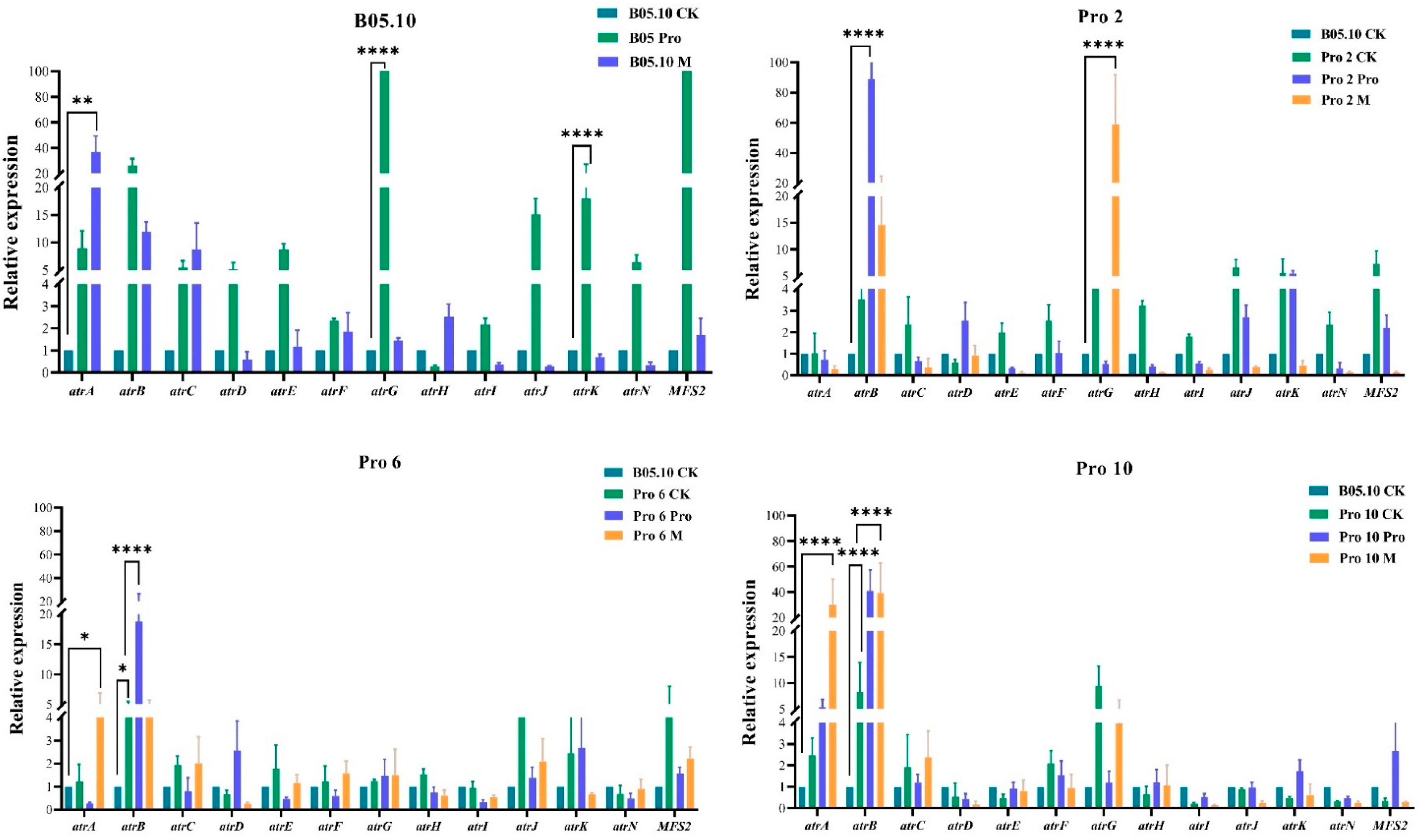
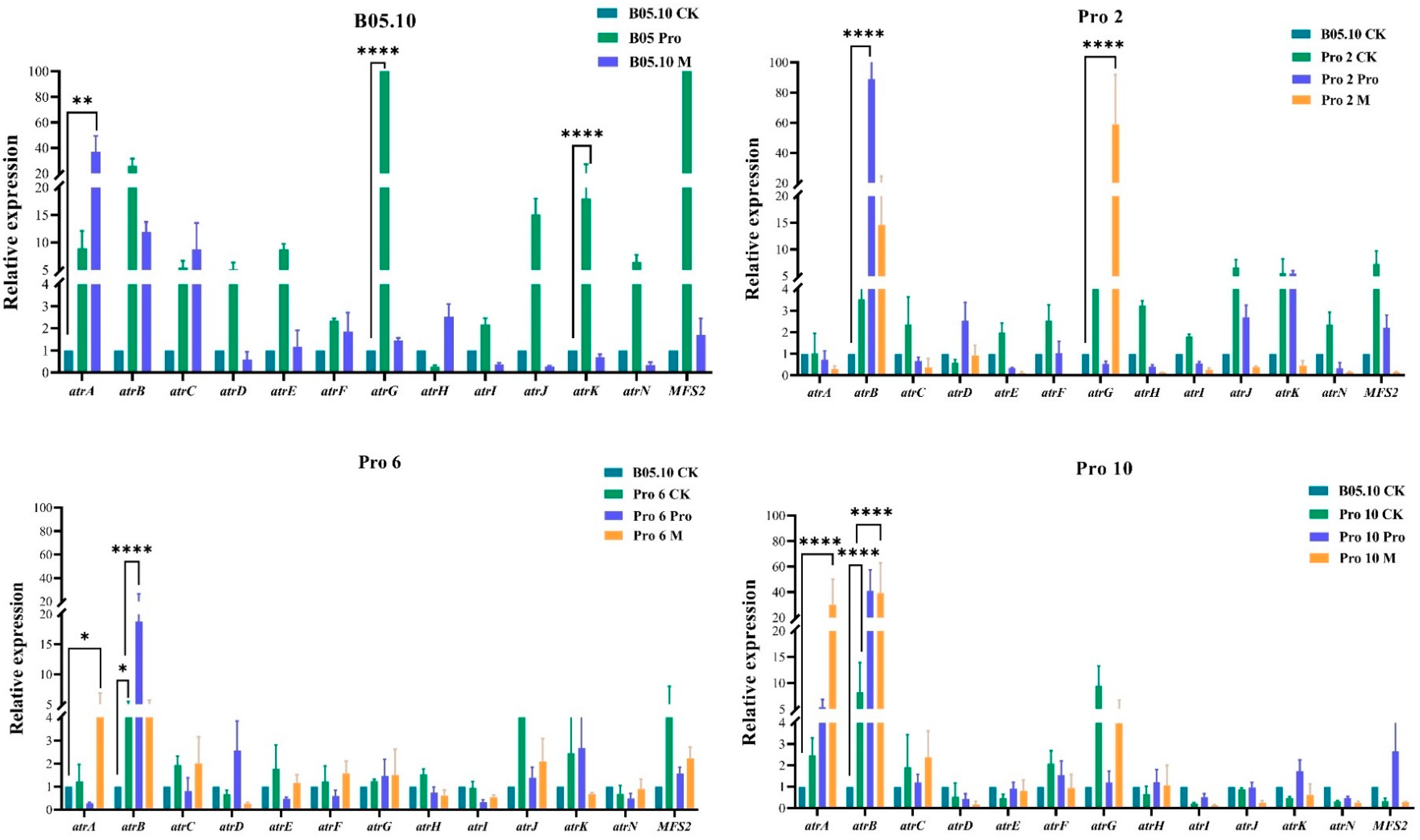
3.7. Expression of ABC and MFS Transporter Genes Using qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fraile, A.; Alonso, A.; Sagasta, E.M. Some characteristics of Botrytis cinerea isolates tolerant to procymidone. Plant Pathol. 1986, 35, 82–85. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Zhu, P.; Wang, Y.; Na, Y.; Guo, H.; Cai, Y.; Nie, H.; Jiang, Y.; Xu, L. A single nucleotide mutation in adenylate cyclase affects vegetative growth, sclerotial formation and virulence of Botrytis cinerea. Int. J. Mol. Sci. 2020, 21, 2912. [Google Scholar] [CrossRef]
- Zhang, C.; Imran, M.; Liu, M.; Li, Z.; Gao, H.; Duan, H.; Zhou, S.; Liu, X. Two point mutations on CYP51 combined with induced expression of the target gene appeared to mediate pyrisoxazole resistance in Botrytis cinerea. Front. Microbiol. 2020, 11, 1396. [Google Scholar] [CrossRef]
- Panayotakou, M.; Malathrakis, N.E. Resistance of Botrytis cinerea to dicarboximide fungicides in protected crops. Ann. Appl. Biol. 1983, 102, 293–299. [Google Scholar] [CrossRef]
- Leroux, P.; Gredt, M.; Leroch, M.; Walker, A. Exploring mechanisms of resistance to respiratory inhibitors in field strains of Botrytis cinerea, the causal agent of gray mold. Appl. Environ. Microbiol. 2010, 76, 6615–6630. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Zwiers, L.H.; De Waard, M.A. Secretion of natural and synthetic toxic compounds from filamentous fungi by membrane transporters of the ATP-binding cassette and major facilitator superfamily. Eur. J. Plant Pathol. 2002, 108, 719–734. [Google Scholar] [CrossRef]
- Pane, C.; Rekab, D.; Firrao, G.; Ruocco, M.; Scala, F. A novel gene coding for an ABC transporter in Botrytis cinerea (Botryotinia fuckeliana) is involved in resistance to H2O2. J. Plant Pathol. 2008, 90, 453–462. [Google Scholar]
- Rupp, S.; Weber, R.W.S.; Rieger, D.; Detzel, P.; Hahn, M. Spread of Botrytis cinerea strains with multiple fungicide resistance in german horticulture. Front. Microbiol. 2017, 7, 228887. [Google Scholar] [CrossRef]
- Schoonbeek, H.; Del Sorbo, G.; De Waard, M.A. The ABC transporter BcatrB affects the sensitivity of Botrytis cinerea to the phytoalexin resveratrol and the fungicide fenpiclonil. Mol. Plant-Microbe Interact. 2001, 14, 562–571. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Srivastava, A.D.; Dafa-Berger, A.; Rotem, N.; Barda, O.; Levy, M. MFS transporter from Botrytis cinerea provides tolerance to glucosinolate-breakdown products and is required for pathogenicity. Nat. Commun. 2019, 10, 2886. [Google Scholar] [CrossRef]
- Pappas, A.C.; Cooke, B.K.; Jordan, V.W.L. Insensitivity of Botrytis cinerea to iprodione, procymidone and vinclozolin and their uptake by the fungus. Plant Pathol. 1979, 28, 71–76. [Google Scholar] [CrossRef]
- Adnan, M.; Hamada, M.S.; Hahn, M.; Li, G.; Luo, C. Fungicide resistance of Botrytis cinerea from strawberry to procymidone and zoxamide in Hubei, China. Phytopathol. Res. 2019, 1, 17. [Google Scholar] [CrossRef]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.J.; Pradier, J.M.; Leroux, P.; De Waard, M.A.; et al. Fungicide-driven evolution and molecular basis of multidrug resistance in field populations of the grey mould fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, Z.; Chen, L.; He, L.; Lu, H.; Ren, Y.; Mu, W.; Liu, F. Baseline sensitivity of Botrytis cinerea to the succinate dehydrogenase inhibitor isopyrazam and efficacy of this fungicide. Plant Dis. 2016, 100, 1314–1320. [Google Scholar] [CrossRef]
- Cheng, X.K.; Man, X.J.; Wang, Z.T.; Liang, L.; Zhang, F.; Wang, Z.W.; Liu, P.F.; Lei, B.; Hao, J.J.; Liu, X.L. Fungicide SYP-14288 inducing multidrug resistance in Rhizoctonia solani. Plant Dis. 2020, 104, 2563–2570. [Google Scholar] [CrossRef]
- Watanabe, M.; Lee, K.; Goto, K.; Kumagai, S.; Sugita-Konishi, Y.; Hara-Kudo, Y. Rapid and effective DNA extraction method with bead grinding for a large amount of fungal DNA. J. Food Prot. 2010, 73, 1077–1084. [Google Scholar] [CrossRef]
- Wu, Z.; Gao, T.; Liang, Z.; Hao, J.; Liu, P.; Liu, X. Dynamic changes in plant secondary metabolites induced by Botrytis cinerea infection. Metabolites 2023, 13, 654. [Google Scholar] [CrossRef]
- Wang, W.; Fang, Y.; Imran, M.; Hu, Z.; Zhang, S.; Huang, Z.; Liu, X. Characterization of the field fludioxonil resistance and its molecular basis in Botrytis cinerea from shanghai province in China. Microorganisms 2021, 9, 266. [Google Scholar] [CrossRef]
- Leroux, P.; Fritz, R.; Debieu, D.; Albertini, C.; Lanen, C.; Bach, J.; Gredt, M.; Chapeland, F. Mechanisms of resistance to fungicides in field strains of Botrytis cinerea. Pest Manag. Sci. 2002, 58, 876–888. [Google Scholar] [CrossRef]
- Shao, W.; Zhao, Y.; Ma, Z. Advances in understanding fungicide resistance in Botrytis cinerea in China. Phytopathology 2021, 111, 455–463. [Google Scholar] [CrossRef]
- Li, X.; Fernández-Ortuño, D.; Grabke, A.; Schnabel, G. Resistance to fludioxonil in Botrytis cinerea isolates from blackberry and strawberry. Phytopathology 2014, 104, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Peng, J.; Wang, X.; Zhang, W.; Zhou, Y.; Wang, H.; Li, X.; Yan, J.; Duan, L. Transcriptomic analysis of resistant and wild-type Botrytis cinerea isolates revealed fludioxonil-resistance mechanisms. Int. J. Mol. Sci. 2023, 24, 988. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Ding, L.; Michailides, T.J.; Li, H.; Ma, Z. Molecular characterization of field azoxystrobin-resistant isolates of Botrytis cinerea. Pestic. Biochem. Physiol. 2009, 93, 72–76. [Google Scholar] [CrossRef]
- Hayashi, K.; Schoonbeek, H.; Sugiura, H.; De Waard, M.A. Multidrug resistance in Botrytis cinerea associated with decreased accumulation of the azole fungicide oxpoconazole and increased transcription of the ABC transporter gene BcatrD. Pestic. Biochem. Physiol. 2001, 70, 168–179. [Google Scholar] [CrossRef]
- Viaud, M.; Fillinger, S.; Liu, W.; Polepalli, J.S.; Le Pecheur, P.; Kunduru, A.R.; Leroux, P.; Legendre, L. A class III histidine kinase acts as a novel virulence factor in Botrytis cinerea. Mol. Plant Microbe 2006, 19, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yan, L.; Luo, Y.; Michailides, T.J. Sequence variation in the two-component histidine kinase gene of Botrytis cinerea associated with resistance to dicarboximide fungicides. Pestic. Biochem. Physiol. 2007, 88, 300–306. [Google Scholar] [CrossRef]
- Liu, S.; Che, Z.; Chen, G. Multiple-fungicide resistance to carbendazim, diethofencarb, procymidone, and pyrimethanil in field isolates of Botrytis cinerea from tomato in Henan province, China. Crop Prot. 2016, 84, 56–61. [Google Scholar] [CrossRef]
- Segmüller, N.; Ellendorf, U.; Tudzynski, B.; Tudzynski, P. Bcsak1, a stress-activated mitogen-activated protein kinase, is involved in vegetative differentiation and pathogenicity in Botrytis cinerea. Eukaryot. Cell 2007, 6, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Liu, N.; Yang, Y.; Yang, Q.; Chen, C.; Gao, Q. The sensor proteins BcSho1 and BcSln1 are involved in, though not essential to, vegetative differentiation, pathogenicity and osmotic stress tolerance in Botrytis cinerea. Front. Microbiol. 2019, 10, 426350. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.K.; Sun, H.H.; Ying, S.H.; Feng, M.G. Characterization of the Hog1 MAPK pathway in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2017, 19, 1808–1821. [Google Scholar] [CrossRef]
- Yamashita, K.; Shiozawa, A.; Watanabe, S.; Fukumori, F.; Kimura, M.; Fujimura, M. Atf-1 transcription factor regulates the expression of ccg-1 and cat-1 genes in response to fludioxonil under OS-2 MAP kinase in Neurospora crassa. Fungal Genet. Biol. 2008, 45, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Madrid, M.; Soto, T.; Franco, A.; Paredes, V.; Vicente, J.; Hidalgo, E.; Gacto, M.; Cansado, J. A cooperative role for Atf1 and Pap1 in the detoxification of the oxidative stress induced by glucose deprivation in Schizosaccharomyces pombe. J. Biol. Chem. 2004, 279, 41594–41602. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Mulford, K.E.; Fassler, J.S. Oxidative stress function of the Saccharomyces cerevisiae Skn7 receiver domain. Eukaryot. Cell 2009, 8, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zhang, X.; Liu, H.; Xu, J.; Sheppard, D.C. Mitogen-activated protein kinase signaling in plant pathogenic fungi. PLoS Pathog. 2018, 14, e1006875. [Google Scholar] [CrossRef] [PubMed]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryot. Cell 2003, 2, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Leroux, P.; Fillinger, S. The hog1-like map kinase Sak1 of Botrytis cinerea is negatively regulated by the upstream histidine kinase Bos1 and is not involved in dicarboximide- and phenylpyrrole-resistance. Fungal Genet. Biol. 2008, 45, 1062–1074. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, D.; Yin, Y.; Cao, Y.; Ma, Z. The response regulator BcSkn7 is required for vegetative differentiation and adaptation to oxidative and osmotic stresses in Botrytis cinerea. Mol. Plant Pathol. 2015, 16, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yang, Q.; Sundin, G.W.; Li, H.; Ma, Z. The mitogen-activated protein kinase kinase BOS5 is involved in regulating vegetative differentiation and virulence in Botrytis cinerea. Fungal Genet. Biol. 2010, 47, 753–760. [Google Scholar] [CrossRef]
- Yan, L.; Yang, Q.; Jiang, J.; Michailides, T.J.; Ma, Z. Involvement of a putative response regulator Brrg-1 in the regulation of sporulation, sensitivity to fungicides, and osmotic stress in Botrytis cinerea. Appl. Microbiol. Biotechnol. 2011, 90, 215–226. [Google Scholar] [CrossRef]
- Wang, H.; Gai, Y.; Zhao, Y.; Wang, M.; Ma, Z. The calcium-calcineurin and high-osmolarity glycerol pathways co-regulate tebuconazole sensitivity and pathogenicity in Fusarium graminearum. Pestic. Biochem. Physiol. 2023, 190, 10531. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences |
|---|---|
| BcActin | F: TCCAAGCGTGGTATTCTTACCC |
| R: TGGTGCTACACGAAGTTCGTTG | |
| BcEF | F: TCCTTCAAGTACGCATGGGT |
| R: GTACCAGCGGCAATGATGAG | |
| BcatrA | F: CTGGACGAGGCTACTTCTGG |
| R: TGTACTCCACGGTCACCAAA | |
| BcatrB | R: AGAGAGGGGTTGCGAATTCA |
| R: AGAGAGGGGTTGCGAATTCA | |
| BcatrC | F: TTTGGAATCCAGAAGCAACC |
| R: TTCTTCGTGGCCTTTGTTCT | |
| BcatrD | F: TCCAGGAGCCAGCAATACAA |
| R: AACCCTGCGGCAAATGAATT | |
| BcatrE | F: ACAATCATCTGCGGGAAAAC |
| R: GAATCTGTGCAACGAAAGCA | |
| BcatrF | F: AGGGGCAAGACACTTTTGTG |
| R: GCCCTTGCAAAATCCTCTTT | |
| BcatrG | F: AGATACTCGGCGTTGCTTGT |
| R: TTTGGCAAAAAGGACGAAAG | |
| BcatrH | F: GGACAACGTGCAAAGATTCA |
| R: TGCCTGTTCCGTGTGTGTAT | |
| BcatrI | F: TTAGATGCCGAATCCGAAAC |
| R: AATTTCGAAAAGGCCGAGTT | |
| BcatrJ | F: CGCTTATCAAGAGCACACCA |
| R: GCCATTCAAATGTGGGAATC | |
| BcatrK | F: CCGCTTTGATGGAGAACGAG |
| R: GTGATGTAGTCGCCACCAAC | |
| BcatrN | F: ACTCCATCCCCAATCGAAA |
| R: AGTGATTGGGCAACTGACA | |
| Bcmfs2 | F: CATGGCTGTCTCATTCGGTG |
| R: GGAATGAAGATGGCGGTTCC | |
| BcBos1 | F: GATGTGGGTGTGGATGGTAAGATGG |
| R: TGCATCATGATTCTCATTTATTCTCAT |
| Strain | EC50 (μg/mL) | ||||||
|---|---|---|---|---|---|---|---|
| Procymidone | Fludioxonil | Azoxystrobin | Fluazinam | Boscalid | Difenoconazole | Pyrimethanil | |
| B05.10 | 0.13 | 0.03 | 0.31 | 0.01 | 1.17 | 1.41 | 0.08 |
| Pro 1 | >100 | >100 | 1.10 | 0.08 | 0.25 | 0.91 | 0.24 |
| Pro 2 | >100 | >100 | 1.79 | 0.16 | 0.30 | 10.54 | 1.07 |
| Pro 3 | >100 | >100 | 0.84 | 0.10 | 0.67 | 3.06 | 0.23 |
| Pro 4 | >100 | >100 | 23.02 | 0.26 | 5.03 | 21.71 | 0.43 |
| Pro 6 | >100 | >100 | 36.97 | 0.23 | 6.30 | 4.94 | 0.32 |
| Pro 7 | >100 | >100 | 3.79 | 0.11 | 4.99 | 1.31 | 0.74 |
| Pro 8 | >100 | >100 | <0.01 | 0.18 | 7.26 | 0.23 | 0.16 |
| Pro 9 | >100 | >100 | 0.21 | 0.10 | 1.93 | 4.31 | 1.21 |
| Pro 10 | >100 | >100 | 7.30 | 0.02 | 6.27 | 0.71 | 0.83 |
| Isolate | Mycelial Growth (mm) | ||||
|---|---|---|---|---|---|
| 4 °C | 14 °C | 18 °C | 25 °C | 30 °C | |
| B05.10 | 17.00 ± 1.73 d * | 39.00 ± 1.00 cde | 42.50 ± 3.27 cd | 58.50 ± 6.68 ab | 43.00 ± 4.12 a |
| Pro 1 | 25.00 ± 3.00 abc | 40.00 ± 3.74 cde | 40.75 ± 3.83 cd | 46.50 ± 1.65 abc | 41.50 ± 4.55 a |
| Pro 2 | 28.00 ± 2.00 a | 48.50 ± 0.86 b | 52.75 ± 0.82 ab | 49.00 ± 1.00 abc | 21.00 ± 1.00 d |
| Pro 3 | 21.50 ± 1.65 bcd | 45.50 ± 2.59 bc | 43.75 ± 0.43 bc | 56.50 ± 6.53 ab | 39.50 ± 0.86 ab |
| Pro 4 | 24.00 ± 1.41 abc | 45.00 ± 3.60 bcd | 44.50 ± 5.12 bc | 50.50 ± 2.17 abc | 29.00 ± 1.73 c |
| Pro 6 | 26.50 ± 1.65 ab | 47.00 ± 2.23 b | 47.50 ± 2.59 abc | 56.00 ± 6.48 ab | 33.00 ± 3.31 bc |
| Pro 7 | 23.00 ± 1.00 abc | 39.00 ± 1.00 cde | 42.00 ± 1.41 cd | 47.00 ± 1.73 abc | 34.00 ± 1.41 bc |
| Pro 8 | 20.50 ± 2.59 cd | 38.50 ± 2.59 de | 39.00 ± 4.12 cd | 44.50 ± 10.52 bc | 28.00 ± 2.00 cd |
| Pro 9 | 26.50 ± 0.86 ab | 34.00 ± 1.41 e | 33.50 ± 1.65 d | 39.00 ± 5.19 c | 30.00 ± 2.00 c |
| Pro 10 | 22.00 ± 1.41 bcd | 57.50 ± 2.59 a | 55.00 ± 6.08 a | 59.50 ± 0.86 a | 6.75 ± 0.43 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Yu, C.; Bi, Q.; Zhang, J.; Hao, J.; Liu, P.; Liu, X. Procymidone Application Contributes to Multidrug Resistance of Botrytis cinerea. J. Fungi 2024, 10, 261. https://doi.org/10.3390/jof10040261
Wu Z, Yu C, Bi Q, Zhang J, Hao J, Liu P, Liu X. Procymidone Application Contributes to Multidrug Resistance of Botrytis cinerea. Journal of Fungi. 2024; 10(4):261. https://doi.org/10.3390/jof10040261
Chicago/Turabian StyleWu, Zhaochen, Chuxian Yu, Qiuyan Bi, Junting Zhang, Jianjun Hao, Pengfei Liu, and Xili Liu. 2024. "Procymidone Application Contributes to Multidrug Resistance of Botrytis cinerea" Journal of Fungi 10, no. 4: 261. https://doi.org/10.3390/jof10040261
APA StyleWu, Z., Yu, C., Bi, Q., Zhang, J., Hao, J., Liu, P., & Liu, X. (2024). Procymidone Application Contributes to Multidrug Resistance of Botrytis cinerea. Journal of Fungi, 10(4), 261. https://doi.org/10.3390/jof10040261