Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment and Dietary Treatments
2.2. Measurements and Sample Collection
2.3. DNA Extraction, 16S rRNA Gene Amplification and Illumina MiSeq Sequencing
2.4. Bioinformatics and Statistical Analyses
3. Results
3.1. Production Traits of Birds
3.2. Determination the Composition of the Competitive Exclusion Product (Broilact®)
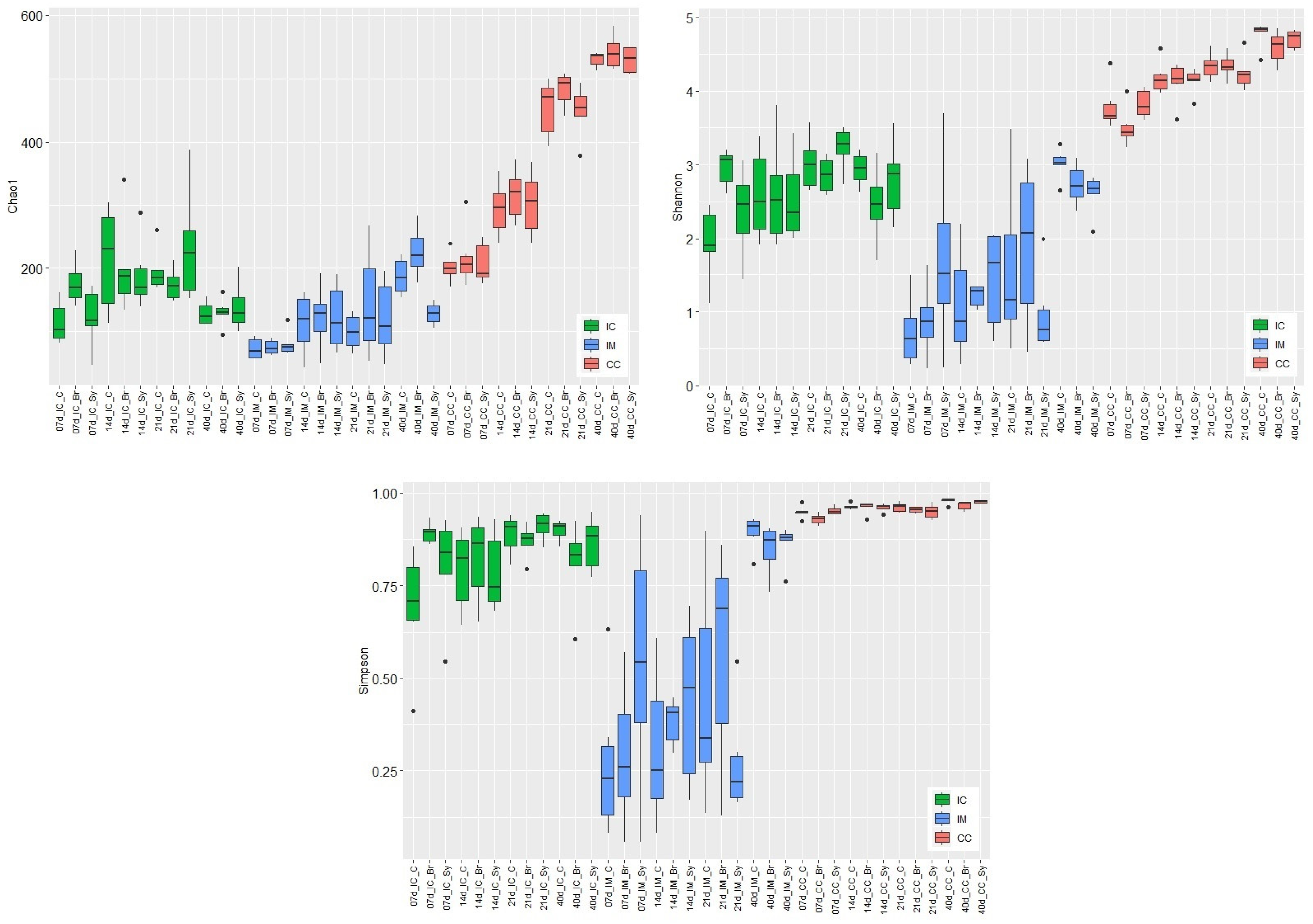
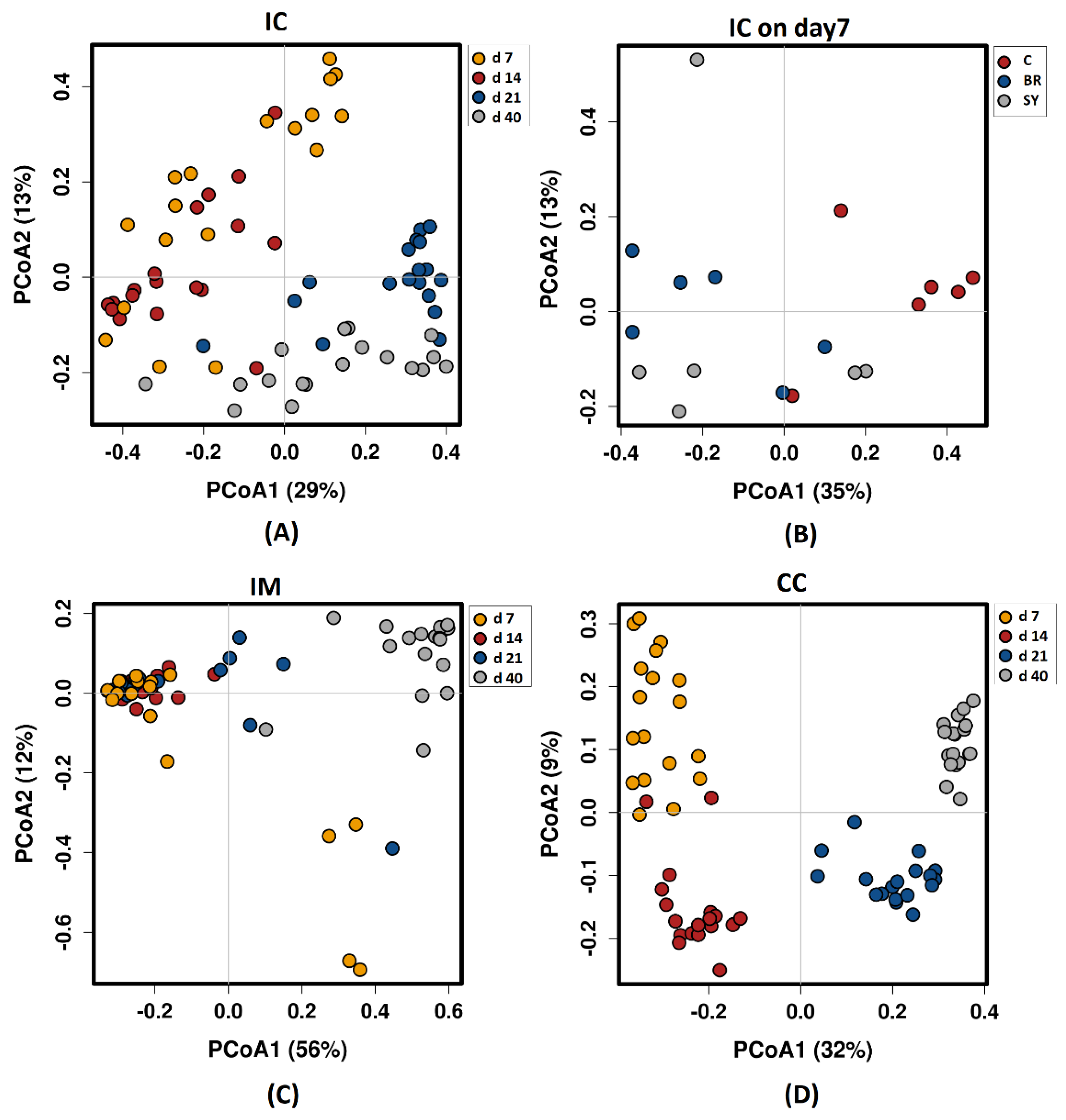
3.3. Alpha and Beta Diversity
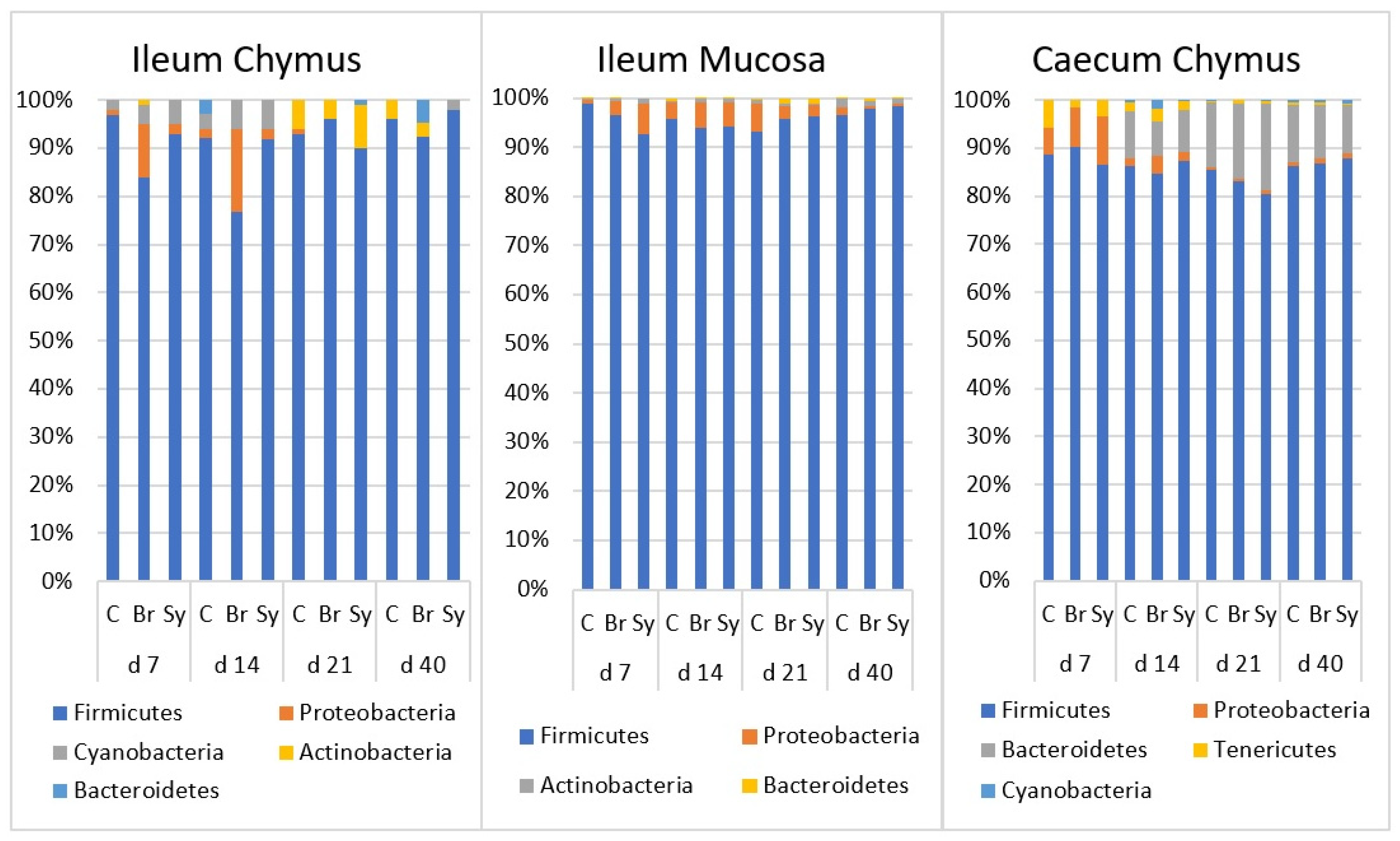
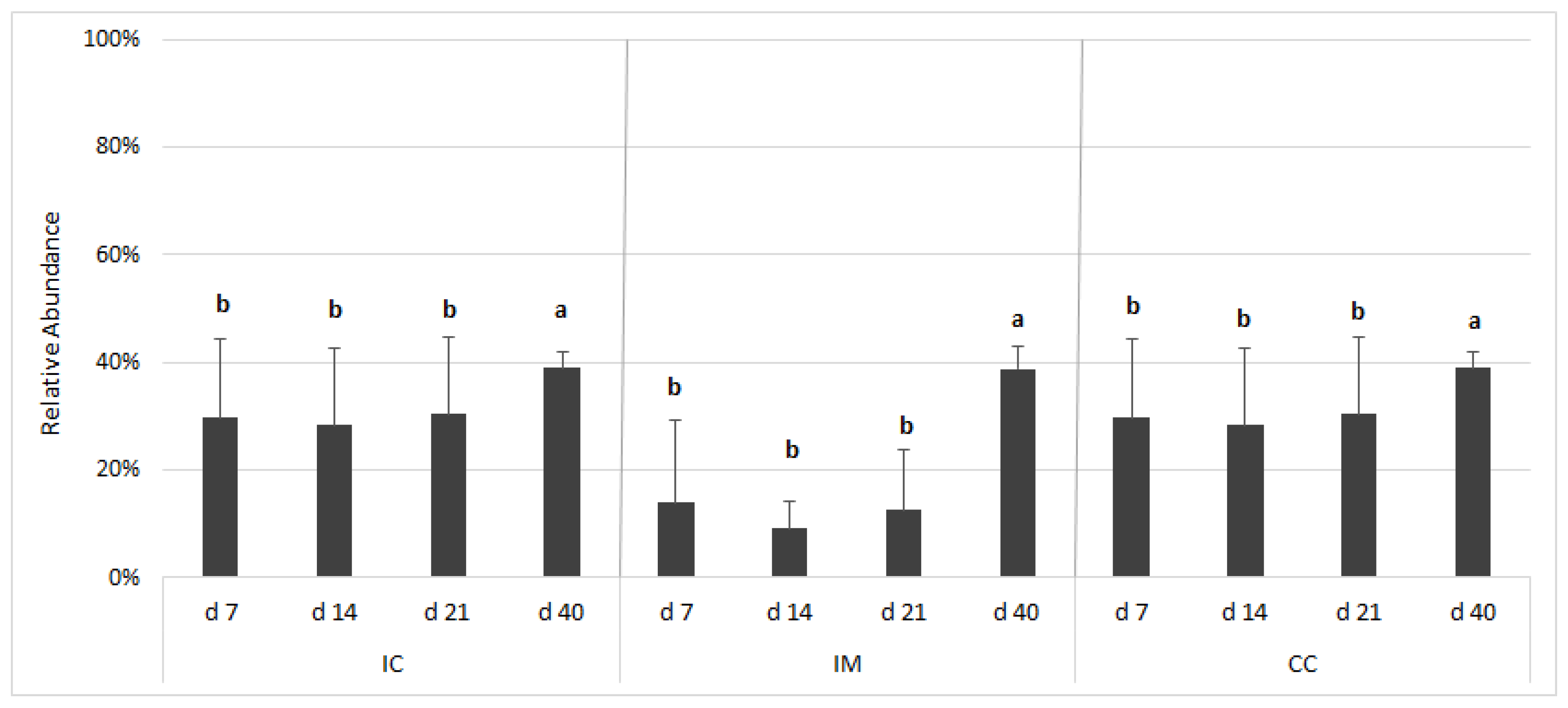
3.4. Taxonomic Composition at Phylum Level and Age-Related Changes in Gut Microbiota
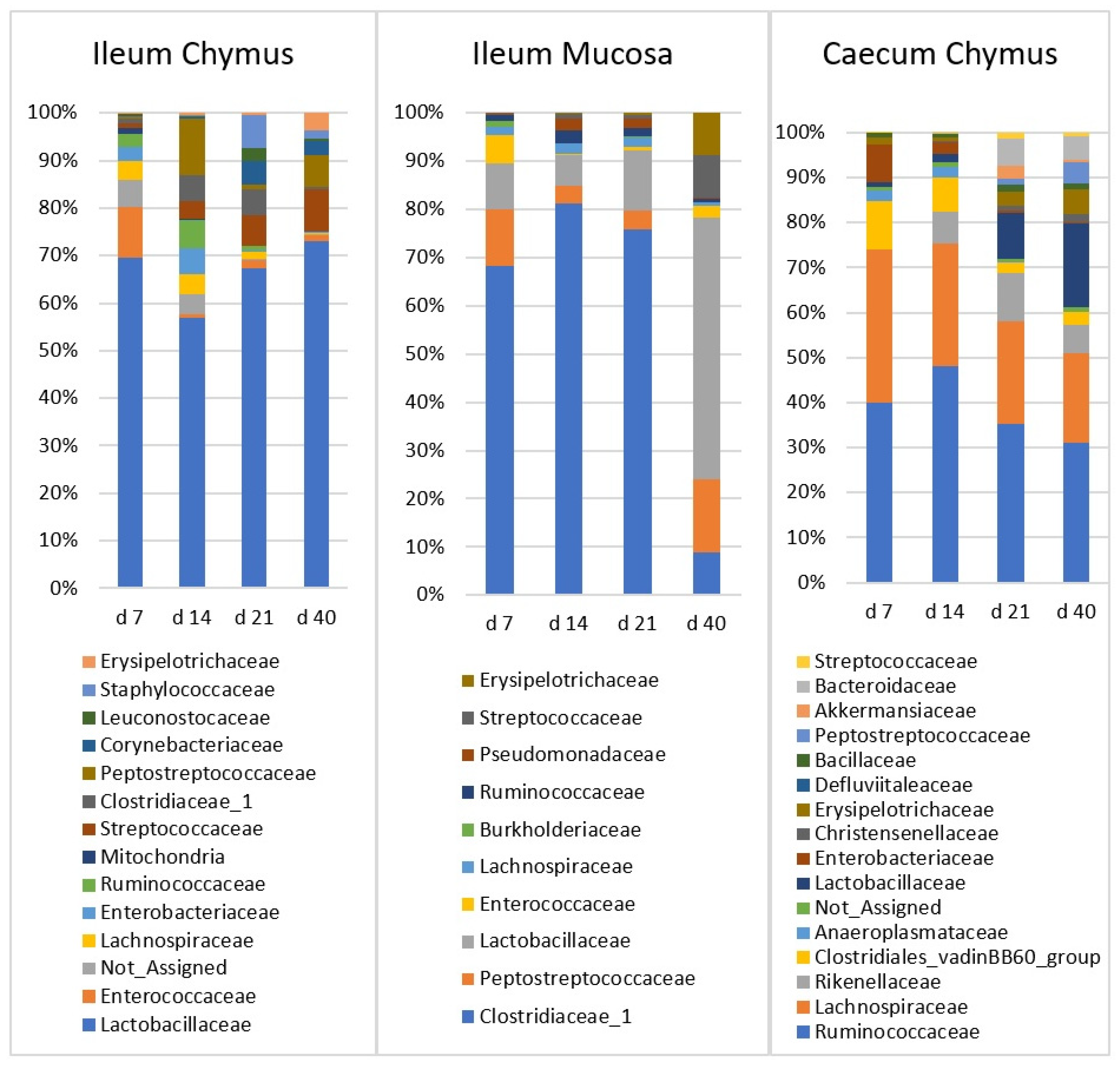
3.5. Taxonomic Composition at Family Level and Age-Related Changes in Gut Microbiota
3.6. Dietary Treatment-Related Changes in Gut Microbiota Composition
3.7. Bugbase
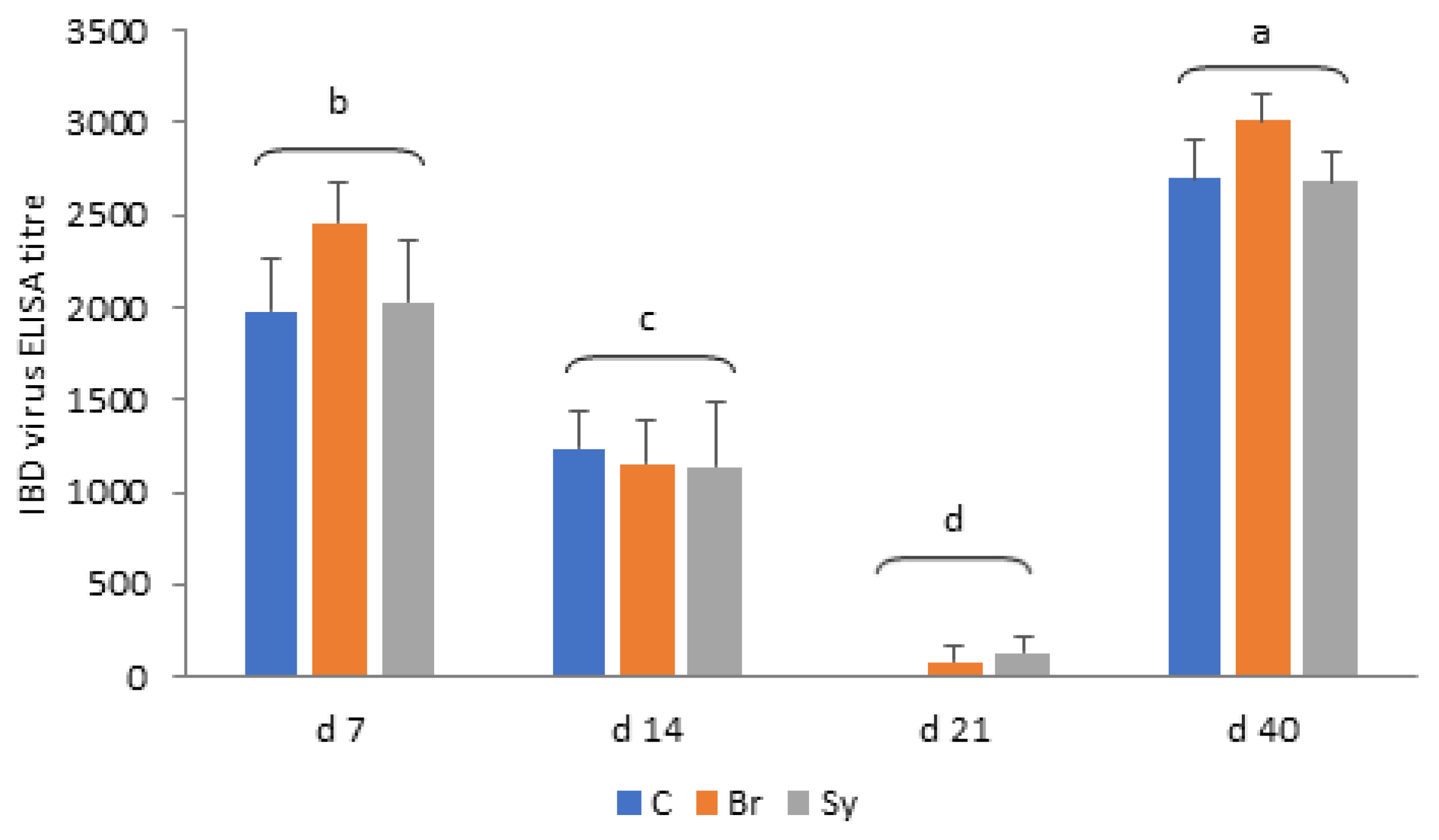
3.8. Infectious Bursal Disease Antibody Titres
3.9. Correlation between the IBD Antibody Titre Values and Microbiota
4. Discussion
4.1. Performance Parameters
4.2. Diversity of Gut Microbiota
4.3. Effects of Age and Dietary Treatments on Composition of Intestinal Microbiota
4.3.1. Ileal Chymus (IC) Microbiota
4.3.2. Ileal Mucus (IM) Microbiota
4.3.3. Caecum Chymus (CC) Microbiota
4.4. Bugbase
4.5. Relationship between the Microbiota Composition and IBD Antibody Titre Values
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Apajalahti, J.; Kettunen, A.; Graham, H. Characteristics of the gastrointestinal microbial communities, with special reference to the chicken. Worlds. Poult. Sci. J. 2004, 60, 223–232. [Google Scholar] [CrossRef]
- Thompson, C.L.; Mikaelyan, A.; Brune, A. Immune-modulating gut symbionts are not “candidatus Arthromitus”. Mucos. Immunol. 2013, 6, 200–201. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Popova, T. Effect of probiotics in poultry for improving meat quality. Curr. Opin. Food Sci. 2017, 14, 72–77. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.S. Benefits of probiotics and/or prebiotics for antibiotic-reduced poultry. Poult. Sci. 2018, 97, 3807–3815. [Google Scholar] [CrossRef]
- Pivnick, H.; Nurmi, E. The Nurmi concepts and its role in the control of Salmonella in poultry. In Developments in Food Microbiology; Davis, R., Ed.; Applied Science Publishers: Basel, Switzerland, 1982; Volume 1, pp. 41–70. [Google Scholar]
- Nurmi, E.; Rantala, M. New aspects of Salmonella infection in broiler production. Nature 1973, 241, 210–211. [Google Scholar] [CrossRef] [PubMed]
- Pedroso, A.A.; Batal, A.B.; Lee, M.D. Effect of in ovo administration of an adult-derived microbiota on establishment of the intestinal microbiome in chickens. Am. J. Vet. Res. 2016, 77, 514–526. [Google Scholar] [CrossRef]
- Meijerink, N.; Kers, J.G.; Velkers, F.C.; van Haarlem, D.A.; Lamot, D.M.; de Oliveira, J.E.; Smidt, H.; Stegeman, J.A.; Rutten, V.P.M.G.; Jansen, C.A. Early Life Inoculation with Adult-Derived Microbiota Accelerates Maturation of Intestinal Microbiota and Enhances NK Cell Activation in Broiler Chickens. Front. Vet. Sci. 2020, 7, 584561. [Google Scholar] [CrossRef]
- Liu, X.; Yan, H.; Lv, L.; Xu, Q.; Yin, C.; Zhang, K.; Wang, P.; Hu, J. Growth performance and meat quality of broiler chickens supplemented with bacillus licheniformis in drinking water. Asian Aust. J. Anim. Sci. 2012, 25, 682–689. [Google Scholar] [CrossRef]
- Kazemi, S.A.; Ahmadi, H.; Karimi Torshizi, M.A. Evaluating two multistrain probiotics on growth performance, intestinal morphology, lipid oxidation and ileal microflora in chickens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Biggs, P.; Parsons, C.M.; Fahey, G.C. The effects of several oligosaccharides on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poult. Sci. 2007, 86, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Böhm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–55. [Google Scholar] [CrossRef]
- Śliżewska, K.; Markowiak-Kopeć, P.; Żbikowski, A.; Szeleszczuk, P. The effect of synbiotic preparations on the intestinal microbiota and her metabolism in broiler chickens. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Brisbin, J.T.; Gong, J.; Parvizi, P.; Sharif, S. Effects of lactobacilli on cytokine expression by chicken spleen and cecal tonsil cells. Clin. Vaccine Immunol. 2010, 17, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Nurmi, E.V.; Schneitz, J.E.; Makela, P.H. Process for the Production of a Bacterial Preparation for the Prophylaxis of Intestinal Disturbances in Poultry. U.S. Patent 4,689,226, 25 August 1987. [Google Scholar]
- Aviagen. Ross Brioler Management Handbook; Aviagen Group, Cummings Research Park, 920 Explorer Boulevard NW: Huntsville, AL, USA, 2018. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.; Caporaso, G.; Blekhman, R.; Knight, R.; et al. BugBase predicts organism-level microbiome phenotypes. bioRxiv 2017, 133462. [Google Scholar] [CrossRef]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, A.A.; Menten, J.F.M.; Lambais, M.R. The structure of bacterial community in the intestines of newly hatched chicks. J. Appl. Poult. Res. 2005, 14, 232–237. [Google Scholar] [CrossRef]
- Donaldson, E.E.; Stanley, D.; Hughes, R.J.; Moore, R.J. The time-course of broiler intestinal microbiota development after administration of cecal contents to incubating eggs. PeerJ 2017, 2017, e3587. [Google Scholar] [CrossRef] [Green Version]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilal, T.; Özpinar, H.; Kutay, C.; Eseceli, H.; Abos, I. The effects of Broilact® on performance and feed digestibility of broilers t Der Effekt von Broilact® auf die Lei stung und Futterverdaulichkeit bei Mastküken. Arch. Geflügelk. 2000, 64, 134–138. [Google Scholar]
- Kefali, S.; Kaygisiz, F.; Toker, N.Y. Effect of probiotics on feed consumption, live weight gain and production cost in broilers. Indian Vet. J. 2007, 267–269. [Google Scholar]
- Schneitz, C. Competitive Exclusion in Poultry-30 Years of Research; Elsevier BV: Amsterdam, The Netherland, 2005; Volume 16. [Google Scholar]
- Reis, M.P.; Fassani, E.J.; Garcia, A.A.P.; Rodrigues, P.B.; Bertechini, A.G.; Barrett, N.; Persia, M.E.; Schmidt, C.J. Effect of Bacillus subtilis (DSM 17299) on performance, digestibility, intestine morphology, and pH in broiler chickens. J. Appl. Poult. Res. 2017, 26, 573–583. [Google Scholar] [CrossRef]
- Mutuş, R.; Kocabaǧli, N.; Alp, M.; Acar, N.; Eren, M.; Gezen, Ş.Ş. The effect of dietary probiotic supplementation on tibial bone characteristics and strength in broilers. Poult. Sci. 2006, 85, 1621–1625. [Google Scholar] [CrossRef]
- Yousefi, M.; Karkoodi, K. Effect of probiotic thepax® and Saccharomyces cerevisiae supplementation on performance and egg quality of laying hens. Int. J. Poult. Sci. 2007, 6, 52–54. [Google Scholar] [CrossRef]
- Oladokun, S.; Koehler, A.; MacIsaac, J.; Ibeagha-Awemu, E.M.; Adewole, D.I. Bacillus subtilis delivery route: Effect on growth performance, intestinal morphology, cecal short-chain fatty acid concentration, and cecal microbiota in broiler chickens. Poult. Sci. 2021, 100, 100809. [Google Scholar] [CrossRef] [PubMed]
- Rafique, K.; Rahman, A.; Mahmood, M. Effect of dietary supplementation of different levels of saccharomyces cerevisiae on growth performance and hematology in broiler. Indian J. Anim. Res. 2020, 54, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef]
- Glendinning, L.; Watson, K.A.; Watson, M. Development of the duodenal, ileal, jejunal and caecal microbiota in chickens. Anim. Microbiome 2019, 1, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, P.; Fothergill, J.; Bernardeau, M.; Wigley, P. Development of the caecal microbiota in three broiler breeds. Front. Vet. Sci. 2019, 6, 201. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.J.; Baetens, J.M.; De Baets, B. Ecological diversity: Measuring the unmeasurable. Mathematics 2018, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Kim, G.B.; Cha, C.J. Spatial heterogeneity and stability of bacterial community in the gastrointestinal tracts of broiler chickens. Poult. Sci. 2014, 93, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Borda-Molina, D.; Vital, M.; Sommerfeld, V.; Rodehutscord, M.; Camarinha-Silva, A. Insights into Broilers’ Gut Microbiota Fed with Phosphorus, Calcium, and Phytase Supplemented Diets. Front. Microbiol. 2016, 7, 2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjitkar, S.; Lawley, B.; Tannock, G.; Engberg, R.M. Bacterial succession in the broiler gastrointestinal tract. Appl. Environ. Microbiol. 2016, 82, 2399–2410. [Google Scholar] [CrossRef] [Green Version]
- Richards-Rios, P.; Fothergill, J.; Bernardeau, M.; Wigley, P. Development of the Ileal Microbiota in Three Broiler Breeds. Front. Vet. Sci. 2020, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Kollarcikova, M.; Kubasova, T.; Karasova, D.; Crhanova, M.; Cejkova, D.; Sisak, F.; Rychlik, I. Use of 16S rRNA gene sequencing for prediction of new opportunistic pathogens in chicken ileal and cecal microbiota. Poult. Sci. 2019, 98, 2347–2353. [Google Scholar] [CrossRef]
- Cisek, A.A.; Binek, M. Chicken intestinal microbiota function with a special emphasis on the role of probiotic bacteria. Pol. J. Vet. Sci. 2014, 17, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Barnes, E.M.; Mead, G.C.; Barnum, D.A.; Harry, E.G. The intestinal flora of the chicken in the period 2 to 6 weeks of age, with particular reference to the anaerobic bacteria. Br. Poult. Sci. 1972, 13, 311–326. [Google Scholar] [CrossRef]
- Rivera-Chávez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microb. 2016, 19, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, W.A.; Mann, E.; Dzieciol, M.; Hess, C.; Schmitz-Esser, S.; Wagner, M.; Hess, M. Age-Related Differences in the Luminal and Mucosa-Associated Gut Microbiome of Broiler Chickens and Shifts Associated with Campylobacter jejuni Infection. Front. Cell. Infect. Microbiol. 2016, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Lilburn, M.; Yu, Z. The Bacteriomes of Ileal Mucosa and Cecal Content of Broiler Chickens and Turkeys as Revealed by Metagenomic Analysis. Int. J. Microbiol. 2016, 2016, 4320412. [Google Scholar] [CrossRef] [Green Version]
- Knarreborg, A.; Brockmann, E.; Høybye, K.; Knap, I.; Lund, B.; Milora, N.; Leser, T.D. Bacillus subtilis (DSM17299) modulates the ileal microbial communities and improves growth performance in broilers | Request PDF. Int. J. Prob. Preb. 2008, 3, 83–88. [Google Scholar]
- Wang, X.; Farnell, Y.Z.; Peebles, E.D.; Kiess, A.S.; Wamsley, K.G.S.; Zhai, W. Effects of prebiotics, probiotics, and their combination on growth performance, small intestine morphology, and resident Lactobacillus of male broilers. Poult. Sci. 2016, 95, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Long, S.; Mahfuz, S.; Wu, D.; Wang, X.; Wei, X.; Piao, X. Effects of probiotics as antibiotics substitutes on growth performance, serum biochemical parameters, intestinal morphology, and barrier function of broilers. Animals 2019, 9, 985. [Google Scholar] [CrossRef] [Green Version]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What makes it tick as successful probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef]
- Klaasen, H.L.B.M.; Koopman, J.P.; Poelma, F.G.J.; Beynen, A.C. Intestinal, segmented, filamentous bacteria. FEMS Microbiol. Lett. 1992, 88, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, A.C.; Hagan, C.E.; Davis, D.J.; Franklin, C.L. Segmented filamentous bacteria: Commensal microbes with potential effects on research. Comp. Med. 2014, 64, 90–98. [Google Scholar]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Yang, H.; Zhang, L.; Su, Y.; Shi, D.; Xiao, H.; Tian, Y. High-throughput sequencing technology to reveal the composition and function of cecal microbiota in Dagu chicken. BMC Microbiol. 2016, 16, 259. [Google Scholar] [CrossRef] [Green Version]
- Oakley, B.B.; Buhr, R.J.; Ritz, C.W.; Kiepper, B.H.; Berrang, M.E.; Seal, B.S.; Cox, N.A. Successional changes in the chicken cecal microbiome during 42 days of growth are independent of organic acid feed additives. BMC Vet. Res. 2014, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocejo, M.; Oporto, B.; Hurtado, A. 16S rRNA amplicon sequencing characterization of caecal microbiome composition of broilers and free-range slow-growing chickens throughout their productive lifespan. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballou, A.L.; Ali, R.A.; Mendoza, M.A.; Ellis, J.C.; Hassan, H.M.; Croom, W.J.; Koci, M.D. Development of the chick microbiome: How early exposure influences future microbial diversity. Front. Vet. Sci. 2016, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.R.; Briggs, W.; Duff, A.; Chasser, K.; Murugesan, R.; Pender, C.; Ramirez, S.; Valenzuela, L.; Bielke, L.R. Comparative effectiveness of probiotic-based formulations on cecal microbiota modulation in broilers. PLoS ONE 2020, 15, e0225871. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Alanis-Lopez, C.; Marroquin-Cardona, A.G.; Kawas, J.R. Composition and Potential Function of Fecal Bacterial Microbiota from Six Bird Species. Birds 2021, 2, 42–59. [Google Scholar] [CrossRef]
- Hamal, K.R.; Burgess, S.C.; Pevzner, I.Y.; Erf, G.F. Maternal antibody transfer from dams to their egg yolks, egg whites, and chicks in meat lines of chickens. Poult. Sci. 2006, 85, 1364–1372. [Google Scholar] [CrossRef]
- Härtle, S.; Magor, K.E.; Göbel, T.W.; Davison, F.; Kaspers, B. Structure and Evolution of Avian Immunoglobulins. In Avian Immunology, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 103–120. ISBN 9780123969651. [Google Scholar]
- Talebi, A.; Pourbakhsh, S.A.; Dorostkar, K. Effects of vaccination routes against IB on performance and immune responses of broiler chickens. Int. J. Poult. Sci. 2005, 4, 795–798. [Google Scholar] [CrossRef] [Green Version]
- Kabir, S.M.L. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef]
- Haghighi, H.R.; Gong, J.; Gyles, C.L.; Hayes, M.A.; Sanei, B.; Parvizi, P.; Gisavi, H.; Chambers, J.R.; Sharif, S. Modulation of antibody-mediated immune response by probiotics in chickens. Clin. Diagn. Lab. Immunol. 2005, 12, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Apata, D.F. Growth performance, nutrient digestibility and immune response of broiler chicks fed diets supplemented with a culture of Lactobacillus bulgaricus. J. Sci. Food Agric. 2008, 88, 1253–1258. [Google Scholar] [CrossRef]
- Hedayati, M.; Manafi, M. Evaluation of Anherbal Compound, a Commercial Probiotic, and an Antibiotic Growth Promoter on the Performance, Intestinal Bacterial Population, Antibody Titers, and Morphology of the Jejunum and Ileum of broilers. Brazilian J. Poult. Sci. 2018, 20, 305–316. [Google Scholar] [CrossRef]
- Janzon, A.; Goodrich, J.K.; Koren, O.; Waters, J.L.; Ley, R.E. Interactions between the Gut Microbiome and Mucosal Immunoglobulins A, M, and G in the Developing Infant Gut. mSystems 2019, 4, e00612-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [CrossRef] [Green Version]
- Methner, U.; Steinbach, G. Wirksamkeit maternaler Salmonellaantikörper gegen eine orale testinfektion von küken mit Salmonella Enteritidis. Berl. Munch. Tierarztl. Wochenschr. 1997, 110, 373–377. [Google Scholar] [PubMed]
- Hornok, S.; Bitay, Z.; Széll, Z.; Varga, I. Assessment of maternal immunity to Cryptosporidium baileyi in chickens. Vet. Parasitol. 1998, 79, 203–212. [Google Scholar] [CrossRef]
- Sahin, O.; Zhang, Q.; Meitzler, J.C.; Harr, B.S.; Morishita, T.Y.; Mohan, R. Prevalence, Antigenic Specificity, and Bactericidal Activity of Poultry Anti-Campylobacter Maternal Antibodies. Appl. Environ. Microbiol. 2001, 67, 3951–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Zhang, Y.; Xiao, K.; Jiang, F.; Wang, H.; Tang, D.; Liu, D.; Liu, B.; Liu, Y.; He, X.; et al. The chicken gut metagenome and the modulatory effects of plant-derived benzylisoquinoline alkaloids 06 Biological Sciences 0605 Microbiology. Microbiome 2018, 6, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Treatments 1 | Body Weight (g/bird) | Daily Weight Gain (g/bird) | ||||||
|---|---|---|---|---|---|---|---|---|
| d 0 | d 7 | d 21 | d 40 | Starter | Grower | Finisher | Total | |
| C | 43 | 260 | 1057 | 2397 | 217 | 797 | 1340 | 2354 |
| Br | 43 | 256 | 1065 | 2398 | 213 | 808 | 1333 | 2355 |
| Sy | 43 | 260 | 1073 | 2441 | 217 | 812 | 1369 | 2398 |
| Pooled SEM | 0.09 | 2.15 | 6.65 | 14.21 | 2.16 | 7.17 | 12.53 | 15.42 |
| p-Values | 0.52 | 0.56 | 0.89 | 0.39 | 0.75 | 0.69 | 0.50 | 0.44 |
| Dietary Treatments 1 | Feed Intake (g/bird) | Feed Conversion Ratio (g/g) | ||||||
|---|---|---|---|---|---|---|---|---|
| Starter | Grower | Finisher | Total | Starter | Grower | Finisher | Total | |
| C | 301 | 1420 | 2298 | 4019 | 1.30 | 1.58 | 1.64 | 1.51 |
| Br | 303 | 1451 | 2283 | 4037 | 1.34 | 1.59 | 1.61 | 1.51 |
| Sy | 290 | 1443 | 2293 | 4026 | 1.26 | 1.57 | 1.59 | 1.48 |
| Pooled SEM | 2.87 | 11.52 | 42.03 | 48.47 | 0.01 | 0.01 | 0.03 | 0.02 |
| p-Values | 0.11 | 0.55 | 0.99 | 0.99 | 0.20 | 0.90 | 0.83 | 0.81 |
| Family | Ileal Chymus | FDR p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Dietary Treatment | Age of Birds | Mean (Dietary Treatment) | |||||||
| d 7 | d 14 | d 21 | d 40 | Dietary Treatment | Age | Int. | |||
| Lactobacillaceae | C | 79.43 A | 59.01 | 63.68 | 60.68 | 65.70 | 0.710 | 0.030 | |
| Br | 49.84 B | 45.53 | 69.02 | 76.02 | 60.10 | ||||
| Sy | 75.20 AB | 56.34 | 51.58 | 67.25 | 62.59 | ||||
| Mean (Age) | 68.16 | 53.62 | 61.43 | 67.99 | 0.130 | ||||
| Enterococcaceae | C | 1.74 B | 0.16 | 1.42 | 1.14 | 1.12 | 0.510 | 0.011 | |
| Br | 18.17 A | 0.32 | 1.88 | 0.92 | 5.32 | ||||
| Sy | 13.76 AB | 1.69 | 2.19 | 1.76 | 4.85 | ||||
| Mean (Age) | 11.22 a | 0.72 b | 1.83 b | 1.27 b | 0.001 | ||||
| Lachnospiraceae | C | 6.88 | 6.68 | 1.02 | 0.15 | 3.68 | 0.640 | 0.530 | |
| Br | 7.55 | 6.28 | 0.70 | 0.19 | 3.68 | ||||
| Sy | 0.49 | 1.94 | 2.99 | 0.35 | 1.44 | ||||
| Mean (Age) | 4.97 | 4.96 | 1.57 | 0.23 | 0.100 | ||||
| Enterobacteriaceae | C | 0.80 | 1.76 | 0.63 A | 0.18 | 0.85 | 0.320 | 0.079 | |
| Br | 9.45 | 16.68 | 0.08 B | 0.17 | 6.60 | ||||
| Sy | 0.07 | 1.62 | 0.14 AB | 0.41 | 0.56 | ||||
| Mean (Age) | 3.44 | 6.69 | 0.28 | 0.25 | 0.065 | ||||
| Ruminococcaceae | C | 4.81 | 9.04 | 1.17 | 0.07 | 3.77 | 0.830 | 0.950 | |
| Br | 3.51 | 4.77 | 0.42 | 0.10 | 2.20 | ||||
| Sy | 1.31 | 5.85 | 2.23 | 0.14 | 2.38 | ||||
| Mean (Age) | 3.21 ab | 6.55 a | 1.27 b | 0.10 b | 0.025 | ||||
| Mitochondria | C | 0.12 | 0.08 | 0.02 | 0.00 | 0.05 | 0.600 | 0.450 | |
| Br | 0.28 | 0.27 | 0.02 | 0.01 | 0.14 | ||||
| Sy | 0.51 | 0.25 | 0.01 | 0.00 | 0.19 | ||||
| Mean (Age) | 0.30 a | 0.20 ab | 0.01 b | 0.00 b | 0.002 | ||||
| Streptococcaceae | C | 1.71 | 4.66 | 8.93 | 12.88 | 7.04 | 0.600 | 0.820 | |
| Br | 1.26 | 4.46 | 5.93 | 6.84 | 4.62 | ||||
| Sy | 0.16 | 4.71 | 6.50 | 7.79 | 4.79 | ||||
| Mean (Age) | 1.04 b | 4.61 b | 7.12 a | 9.17 a | 0.001 | ||||
| Clostridiaceae_1 | C | 0.65 | 0.19 | 6.20 | 0.44 | 1.87 | 0.600 | 0.420 | |
| Br | 0.75 | 1.35 | 5.83 | 0.11 | 2.01 | ||||
| Sy | 0.65 | 11.47 | 6.45 | 1.26 | 4.96 | ||||
| Mean (Age) | 0.68 | 4.34 | 6.16 | 0.61 | 0.075 | ||||
| Peptostreptococcaceae | C | 1.08 | 10.57 | 0.31 | 8.78 | 5.18 | 0.860 | 0.950 | |
| Br | 1.08 | 11.97 | 0.96 | 6.64 | 5.16 | ||||
| Sy | 0.04 | 7.22 | 2.17 | 6.87 | 4.07 | ||||
| Mean (Age) | 0.73 b | 9.92 a | 1.15 b | 7.43 a | 0.001 | ||||
| Corynebacteriaceae | C | 0.05 | 0.12 | 5.63 | 3.22 | 2.25 | 0.600 | 0.770 | |
| Br | 0.56 | 0.07 | 3.49 | 2.01 | 1.53 | ||||
| Sy | 0.09 | 0.25 | 8.08 | 4.49 | 3.23 | ||||
| Mean (Age) | 0.23 b | 0.15 b | 5.73 a | 3.24 ab | 0.001 | ||||
| Leuconostocaceae | C | 0.05 | 0.30 | 2.63 | 0.75 | 0.93 | 0.600 | 0.230 | |
| Br | 0.40 | 0.28 | 1.69 | 0.48 | 0.71 | ||||
| Sy | 0.02 | 0.21 | 4.53 | 0.72 | 1.37 | ||||
| Mean (Age) | 0.16 b | 0.26 b | 2.95 a | 0.65 b | 0.001 | ||||
| Staphylococcaceae | C | 0.04 | 0.81 | 5.72 | 2.41 | 2.24 | 0.860 | 0.880 | |
| Br | 0.24 | 0.19 | 8.02 | 0.99 | 2.36 | ||||
| Sy | 0.08 | 0.56 | 7.85 | 2.57 | 2.76 | ||||
| Mean (Age) | 0.12 b | 0.52 b | 7.20 a | 1.99 b | 0.001 | ||||
| Erysipelotrichaceae | C | 0.04 | 0.21 | 0.19 | 6.38 | 1.71 | 0.770 | 0.360 | |
| Br | 0.21 | 0.13 | 0.14 | 3.38 | 0.97 | ||||
| Sy | 0.04 | 0.98 | 0.68 | 2.04 | 0.93 | ||||
| Mean (Age) | 0.10 b | 0.44 b | 0.34 b | 3.93 a | 0.001 | ||||
| Bacillaceae | C | 0.01 B | 0.03 | 0.02 | 0.002 B | 0.01 B | 0.007 | 0.370 | |
| Br | 0.02 B | 0.05 | 0.01 | 0.002 B | 0.02B | ||||
| Sy | 0.14 A | 0.2 | 0.28 | 0.04 A | 0.16A | ||||
| Mean (Age) | 0.06 | 0.09 | 0.10 | 0.02 | 0.230 | ||||
| Family | Ileal Mucosa | FDR p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Dietary Treatment | Age of Birds | Mean (Dietary Treatment) | |||||||
| d 7 | d 14 | d 21 | d 40 | Dietary Treatment | Day | Int. | |||
| Clostridiaceae_1 | C | 83.77 | 82.22 | 70.38 | 4.26 | 60.16 | 0.590 | 0.022 | |
| Br | 72.28 | 78.10 | 51.62 | 4.29 | 51.57 | ||||
| Sy | 42.28 | 72.84 | 84.99 | 12.22 | 53.08 | ||||
| Mean (Age) | 66.11 a | 77.72 a | 69.00 a | 6.92 b | 0.001 | ||||
| Peptostreptococcaceae | C | 1.06 | 2.43 | 1.93 | 17.75 | 5.79 | 0.730 | 0.590 | |
| Br | 11.09 | 4.26 | 10.64 | 10.90 | 9.22 | ||||
| Sy | 12.38 | 1.62 | 1.98 | 18.14 | 8.53 | ||||
| Mean (Age) | 8.18 ab | 2.77 b | 4.85 ab | 15.60 a | 0.091 | ||||
| Lactobacillaceae | C | 6.48 | 4.00 | 11.91 AB | 41.35 | 15.94 | 0.590 | 0.025 | |
| Br | 6.00 | 5.93 | 22.64 A | 64.41 | 24.74 | ||||
| Sy | 16.25 | 9.96 | 3.90 B | 47.03 | 19.28 | ||||
| Mean (Age) | 9.58 b | 6.63 b | 12.82 b | 50.93 a | 0.001 | ||||
| Enterococcaceae | C | 6.47 | 0.05 | 0.45 | 3.38 | 2.59 | 0.750 | 0.950 | |
| Br | 3.86 | 0.06 | 1.51 | 1.13 | 1.64 | ||||
| Sy | 11.95 | 0.79 | 0.25 | 3.62 | 4.15 | ||||
| Mean (Age) | 7.42 | 0.30 | 0.74 | 2.71 | 0.250 | ||||
| Lachnospiraceae | C | 0.22 | 1.92 | 1.76 | 0.56 | 1.11 | 0.680 | 0.680 | |
| Br | 1.27 | 1.99 | 3.21 | 1.39 | 1.97 | ||||
| Sy | 3.41 | 1.94 | 1.74 | 0.32 | 1.86 | ||||
| Mean (Age) | 1.63 | 1.95 | 2.24 | 0.76 | 0.490 | ||||
| Burkholderiaceae | C | 0.45 | 0.06 | 0.19 | 0.01 | 0.18 | 0.590 | 0.110 | |
| Br | 1.18 | 0.08 | 0.09 | 0.01 | 0.34 | ||||
| Sy | 2.67 | 0.03 | 0.11 | 0.00 | 0.70 | ||||
| Mean (Age) | 1.43 a | 0.06 b | 0.13 b | 0.01 b | 0.002 | ||||
| Ruminococcaceae | C | 0.19 | 2.82 | 2.12 | 0.60 | 1.44 | 0.990 | 0.600 | |
| Br | 0.83 | 1.52 | 2.32 | 1.54 | 1.55 | ||||
| Sy | 2.59 | 2.36 | 1.17 | 0.16 | 1.57 | ||||
| Mean (Age) | 1.20 | 2.24 | 1.87 | 0.77 | 0.440 | ||||
| Pseudomonadaceae | C | 0.06 | 1.98 | 3.46 | 0.16 | 1.41 | 0.980 | 0.170 | |
| Br | 0.25 | 3.32 | 1.60 | 0.10 | 1.32 | ||||
| Sy | 0.69 | 3.11 | 1.32 | 0.09 | 1.30 | ||||
| Mean (Age) | 0.33 b | 2.80 a | 2.12 a | 0.12 b | 0.001 | ||||
| Streptococcaceae | C | 0.17 | 0.50 | 1.01 | 14.73 | 4.10 | 0.590 | 0.048 | |
| Br | 0.20 | 0.85 | 1.39 | 6.76 | 2.30 | ||||
| Sy | 0.19 | 2.02 | 0.51 | 5.40 | 2.03 | ||||
| Mean (Age) | 0.19 b | 1.12 b | 0.97 b | 8.96 a | 0.001 | ||||
| Erysipelotrichaceae | C | 0.03 | 0.09 | 0.84 | 10.91 | 2.97 | 0.670 | 0.810 | |
| Br | 0.01 | 0.05 | 0.19 | 6.06 | 1.58 | ||||
| Sy | 0.12 | 0.13 | 0.24 | 9.31 | 2.45 | ||||
| Mean (Age) | 0.06 b | 0.09 b | 0.43 b | 8.76 a | 0.001 | ||||
| Bacillaceae | C | 0.26 | 0.17 | 0.17 | 0.01 | 0.15 | 0.590 | 0.077 | |
| Br | 0.67 | 0.09 | 0.14 | 0.04 | 0.24 | ||||
| Sy | 1.66 | 0.25 | 0.09 | 0.01 | 0.50 | ||||
| Mean (Age) | 0.86 a | 0.17 b | 0.14 c | 0.02 d | 0.003 | ||||
| Family | Caecum Chymus | FDR p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Dietary Treatment | Age of Birds | Mean (Dietary Treatment) | |||||||
| d 7 | d 14 | d 21 | d 40 | Dietary Treatment | Age | Int. | |||
| Ruminococcaceae | C | 41.49 | 42.64 | 36.99 | 29.84 | 37.74 | 0.960 | 0.920 | |
| Br | 41.54 | 47.36 | 36.87 | 30.13 | 38.98 | ||||
| Sy | 39.87 | 49.55 | 35.57 | 29.03 | 38.50 | ||||
| Mean (Age) | 40.96 ab | 46.52 a | 36.48 bc | 29.67 c | 0.001 | ||||
| Lachnospiraceae | C | 30.35 | 30.30 | 21.86 | 19.39 | 25.47 | 0.960 | 0.910 | |
| Br | 30.20 | 27.64 | 21.22 | 18.46 | 24.38 | ||||
| Sy | 33.51 | 25.03 | 21.67 | 18.11 | 24.58 | ||||
| Mean (Age) | 31.35 a | 27.65 ab | 21.58 bc | 18.65 c | 0.001 | ||||
| Rikenellaceae | C | 0.01 | 8.90 | 10.41 | 5.37 | 6.17 | 0.970 | 0.830 | |
| Br | 0.06 | 7.10 | 8.52 | 7.33 | 5.75 | ||||
| Sy | 0.00 | 8.62 | 8.15 | 6.61 | 5.84 | ||||
| Mean (Age) | 0.02 b | 8.21 a | 9.03 a | 6.44 a | 0.001 | ||||
| Clostridiales vadinBB60 group | C | 10.92 | 7.53 | 1.95 | 2.55 | 5.74 | 0.960 | 0.430 | |
| Br | 15.68 | 6.27 | 2.03 | 2.32 | 6.57 | ||||
| Sy | 8.00 | 8.38 | 2.82 | 1.01 | 5.05 | ||||
| Mean (Age) | 11.53 a | 7.39 a | 2.27 b | 1.96 b | 0.001 | ||||
| Anaeroplasmataceae | C | 4.89 | 1.87 | 0.08 | 0.06 | 1.72 | 0.960 | 0.430 | |
| Br | 1.31 | 2.37 | 0.31 | 0.08 | 1.02 | ||||
| Sy | 2.88 | 1.76 | 0.08 | 0.04 | 1.19 | ||||
| Mean (Age) | 3.03 a | 2.00 ab | 0.16 b | 0.06 b | 0.001 | ||||
| Lactobacillaceae | C | 1.67 | 3.03 | 10.95 | 15.45 | 7.78 | 0.960 | 0.020 | |
| Br | 0.31 | 0.91 | 10.87 | 21.61 | 8.43 | ||||
| Sy | 1.74 | 1.73 | 8.02 | 20.85 | 8.09 | ||||
| Mean (Age) | 1.24 c | 1.89 c | 9.95 b | 19.30 a | 0.001 | ||||
| Enterobacteriaceae | C | 5.46 | 1.52 | 0.50 | 0.16 | 1.91 | 0.960 | 0.770 | |
| Br | 8.16 | 3.62 | 0.38 | 0.10 | 3.07 | ||||
| Sy | 10.06 | 1.94 | 0.59 | 0.18 | 3.19 | ||||
| Mean (Age) | 7.89 a | 2.36 b | 0.49 b | 0.15 b | 0.001 | ||||
| Christensenellaceae | C | 0.20 | 0.16 | 1.26 | 1.69 | 0.83 | 0.990 | 0.890 | |
| Br | 0.07 | 0.33 | 1.07 | 1.88 | 0.84 | ||||
| Sy | 0.06 | 0.43 | 1.13 | 1.77 | 0.85 | ||||
| Mean (Age) | 0.11 c | 0.31 c | 1.15 b | 1.78 a | 0.001 | ||||
| Erysipelotrichaceae | C | 1.50 | 0.95 A | 3.47 | 6.85 A | 3.19 | 0.370 | 0.150 | |
| Br | 1.02 | 0.52 B | 2.97 | 4.31 B | 2.21 | ||||
| Sy | 1.65 | 0.64 AB | 3.39 | 5.34 AB | 2.75 | ||||
| Mean (Age) | 1.39 c | 0.70 c | 3.28 b | 5.50 a | 0.001 | ||||
| Bacillaceae | C | 1.53 | 0.89 | 2.07 | 0.95 | 1.36 | 0.940 | 0.980 | |
| Br | 0.87 | 0.72 | 1.28 | 0.63 | 0.88 | ||||
| Sy | 0.82 | 0.64 | 1.27 | 0.85 | 0.89 | ||||
| Mean (Age) | 1.07 | 0.75 | 1.54 | 0.81 | 0.220 | ||||
| Peptostreptococcaceae | C | 0.08 | 0.04 | 1.75 | 4.81 | 1.67 | 0.940 | 0.160 | |
| Br | 0.03 | 0.06 | 1.23 | 4.09 | 1.35 | ||||
| Sy | 0.07 | 0.07 | 0.96 | 5.40 | 1.63 | ||||
| Mean (Age) | 0.06 c | 0.06 c | 1.31 b | 4.77 a | 0.001 | ||||
| Akkermansiaceae | C | 0.00 | 0.00 | 2.59 | 0.49 | 0.77 | 0.970 | 1.000 | |
| Br | 0.00 | 0.01 | 2.78 | 0.77 | 0.89 | ||||
| Sy | 0.00 | 0.04 | 2.89 | 0.82 | 0.94 | ||||
| Mean (Age) | 0.00 b | 0.01 b | 2.75 a | 0.69 b | 0.001 | ||||
| Bacteroidaceae | C | 0.00 | 0.71 | 2.61 | 6.44 | 0.83 | 0.960 | 0.230 | |
| Br | 0.00 | 0.00 | 6.77 | 3.93 | 1.69 | ||||
| Sy | 0.00 | 0.00 | 9.45 | 3.31 | 2.36 | ||||
| Mean (Age) | 0.00 b | 0.24 b | 6.28 a | 4.56 a | 0.001 | ||||
| Streptococcaceae | C | 0.23 | 0.14 | 1.62 | 1.45 | 0.86 | 0.960 | 0.900 | |
| Br | 0.03 | 0.04 | 1.44 | 0.69 | 0.55 | ||||
| Sy | 0.02 | 0.11 | 2.09 | 0.68 | 0.72 | ||||
| Mean (Age) | 0.09 b | 0.10 b | 1.72 a | 0.94 ab | 0.001 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Such, N.; Farkas, V.; Csitári, G.; Pál, L.; Márton, A.; Menyhárt, L.; Dublecz, K. Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens. Vet. Sci. 2021, 8, 187. https://doi.org/10.3390/vetsci8090187
Such N, Farkas V, Csitári G, Pál L, Márton A, Menyhárt L, Dublecz K. Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens. Veterinary Sciences. 2021; 8(9):187. https://doi.org/10.3390/vetsci8090187
Chicago/Turabian StyleSuch, Nikoletta, Valéria Farkas, Gábor Csitári, László Pál, Aliz Márton, László Menyhárt, and Károly Dublecz. 2021. "Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens" Veterinary Sciences 8, no. 9: 187. https://doi.org/10.3390/vetsci8090187
APA StyleSuch, N., Farkas, V., Csitári, G., Pál, L., Márton, A., Menyhárt, L., & Dublecz, K. (2021). Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens. Veterinary Sciences, 8(9), 187. https://doi.org/10.3390/vetsci8090187