
Molecular Survey of Babesia and Anaplasma Infection in Cattle in Bolivia

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
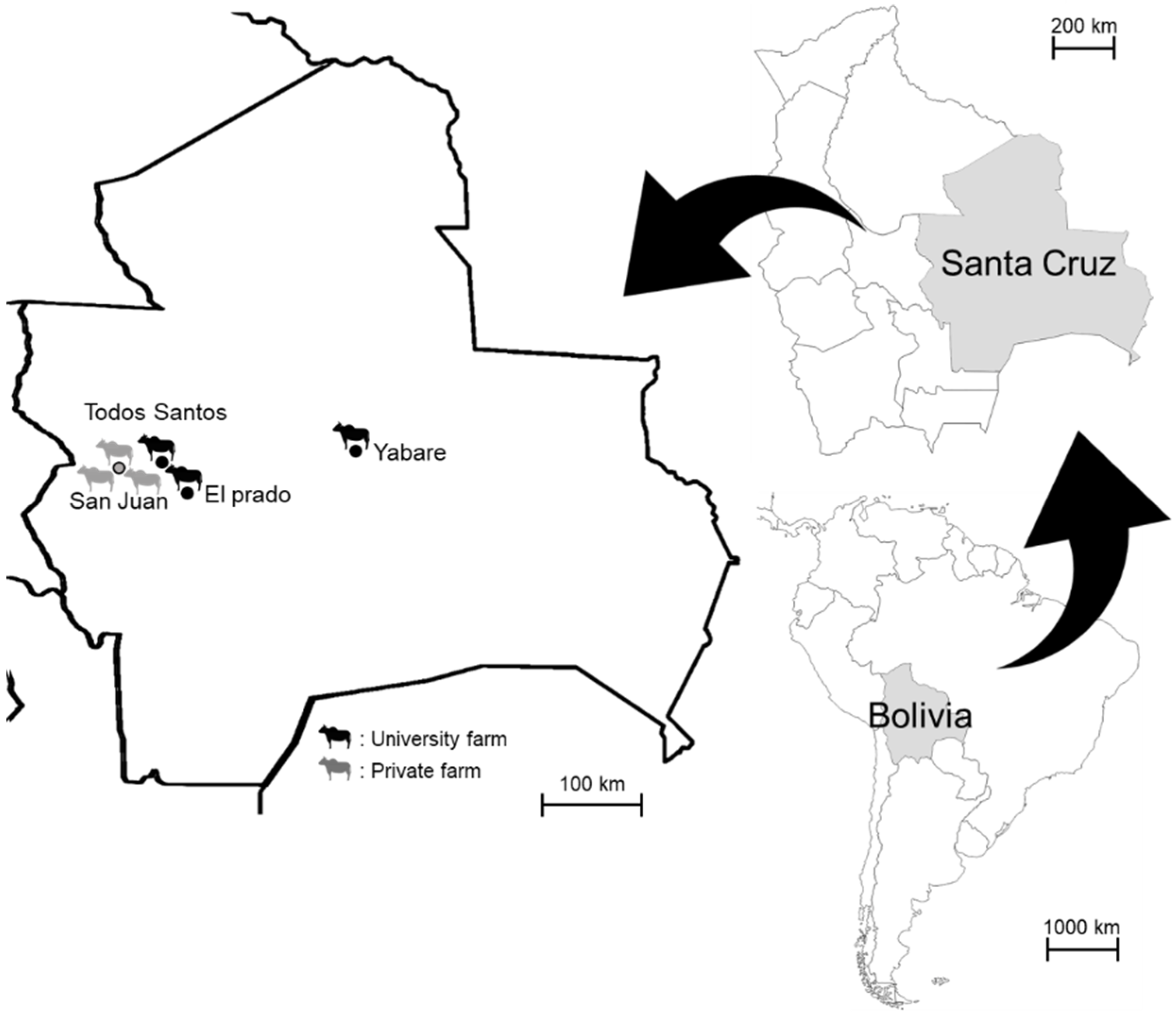
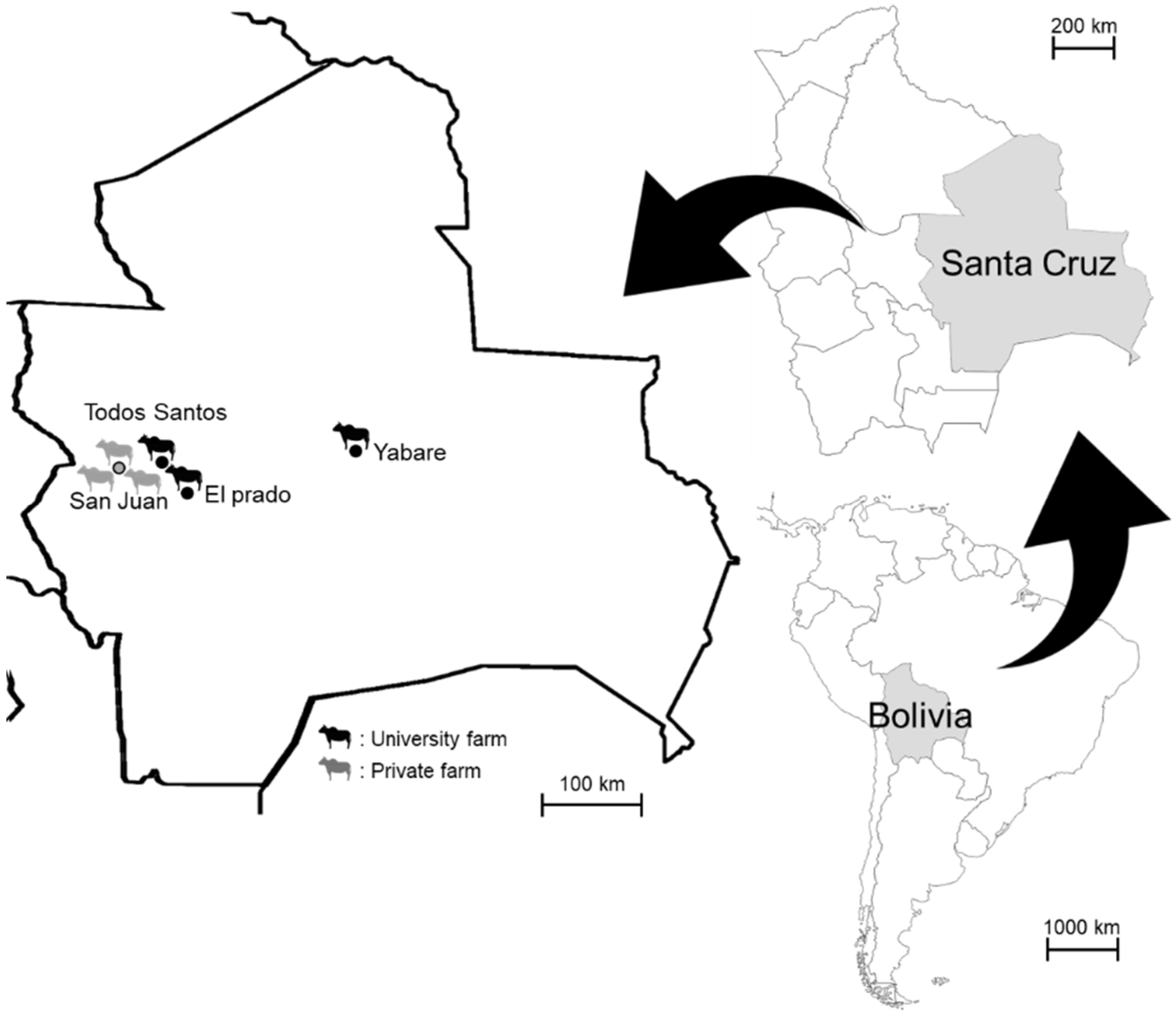
2. Materials and Methods
2.1. Blood Sampling and DNA Extraction
2.2. Detection and Characterisation of Babesia spp.
2.3. Detection and Characterisation of Anaplasmataceae
2.4. Sanger Sequencing
2.5. Data Analysis
3. Results
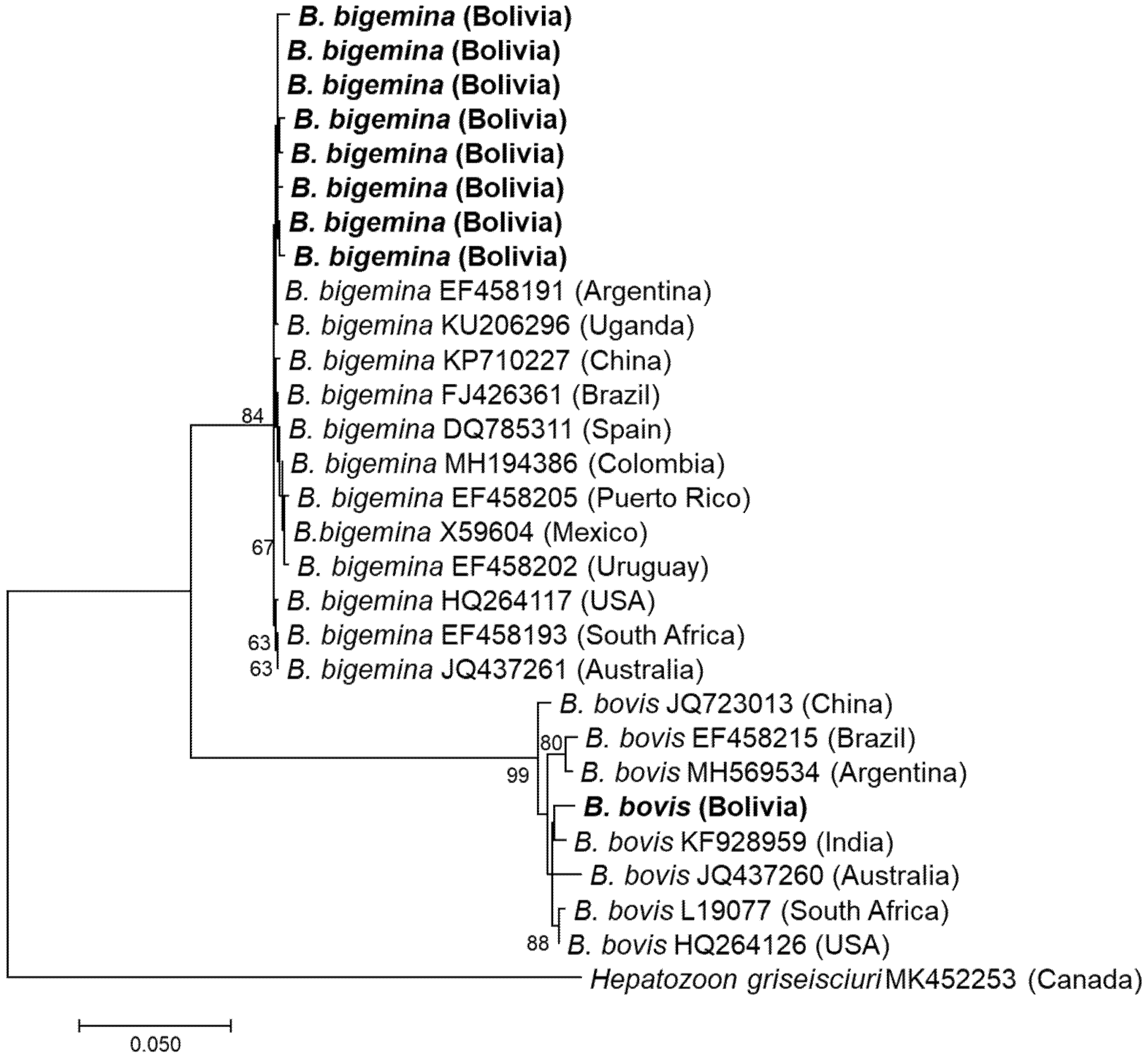
3.1. Detection and Characterisation of Babesia
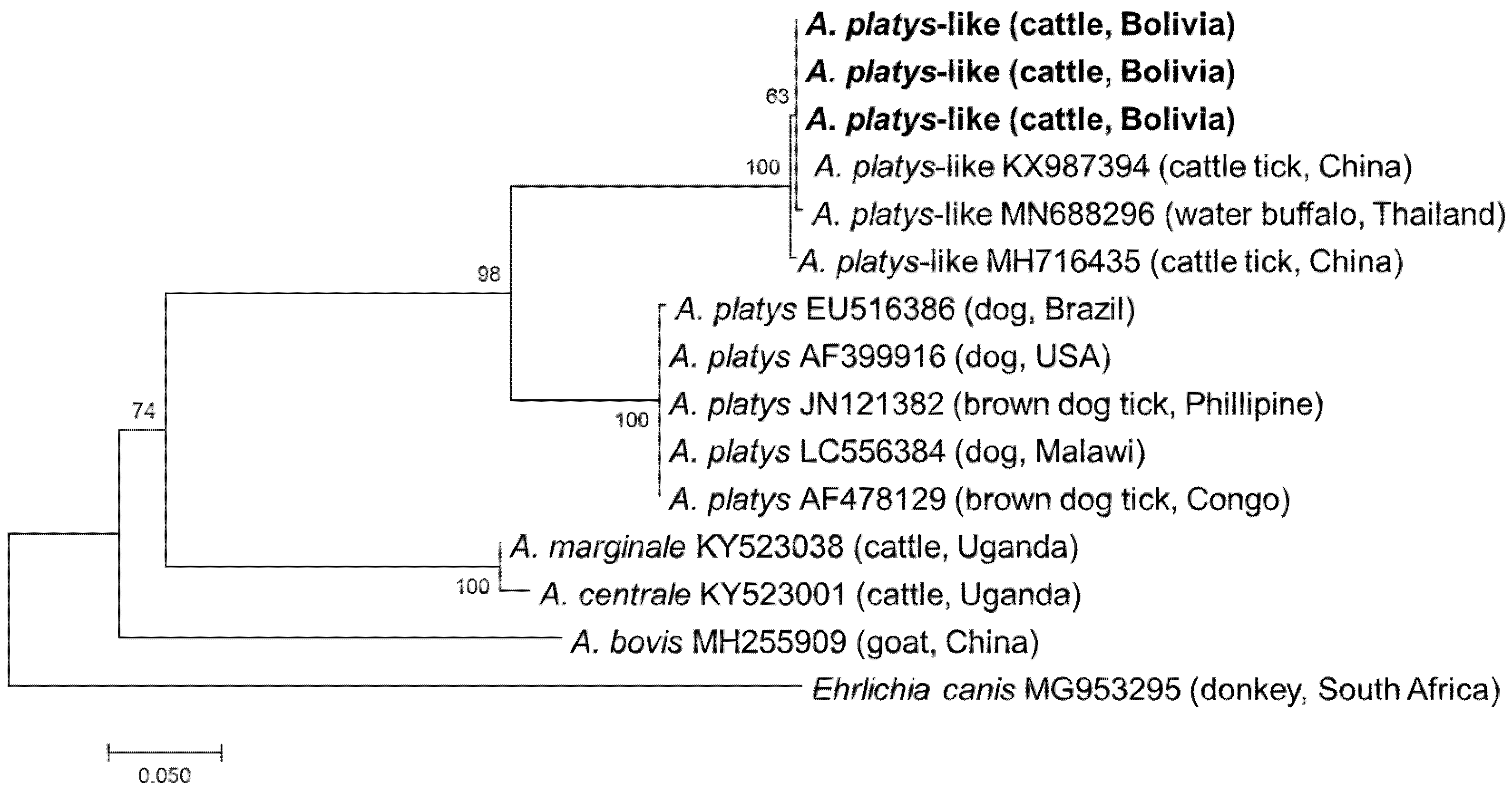
3.2. Detection and Characterisation of Anaplasmataceae
3.3. Co-Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, G.W.; Anderson, D.P. The Latin American Livestock Industry: Growth and Challenges. Choices 2020, 34, 1–11. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Livestock Production in Latin America and the Caribbean. 2017. Available online: http://www.fao.org/americas/priorities/produccion-pecuaria/en/ (accessed on 2 September 2021).
- Grisi, L.; Leite, R.C.; Martins, J.R.D.S.; De Barros, A.T.M.; Andreotti, R.; Cançado, P.H.D.; De León, A.A.P.; Pereira, J.B.; Villela, H.S. Reassessment of the potential economic impact of cattle parasites in Brazil. Bras. Parasitol. Vet. 2014, 23, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, L.L.; Canever, M.F.; Cardozo, L.L.; Cardozo, C.P.; Herkenhoff, M.E.; Neto, A.T.; Vogel, C.I.G.; Miletti, L.C. Prevalence of Anaplasma marginale, Babesia bovis, and Babesia bigemina in cattle in the Campos de Lages region, Santa Catarina state, Brazil, estimated by multiplex-PCR. Parasite Epidemiol. Control 2019, 6, e00114. [Google Scholar] [CrossRef] [PubMed]
- Parodi, P.; Corbellini, L.G.; Leotti, V.B.; Rivero, R.; Miraballes, C.; Correa, F.R.; Venzal, J.M.; Fernández, M.T.A. Validation of a multiplex PCR assay to detect Babesia spp. and Anaplasma marginale in cattle in Uruguay in the absence of a gold standard test. J. Vet. Diagn. Investig. 2021, 33, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Obregón, D.; Cruz, A.C.; Armas, Y.; Silva, J.B.; Fonseca, A.H.; André, M.R.; Alfonso, P.; Oliveira, M.C.S.; Machado, R.Z.; González, B.C. High co-infection rates of Babesia bovis, Babesia bigemina, and Anaplasma marginale in water buffalo in Western Cuba. Parasitol. Res. 2019, 118, 955–967. [Google Scholar] [CrossRef]
- Dueñez, J.J.; Chávez, O.T.; Rocha, A.H.; Castaño, A.T.; Jaramillo, A.M.M. Molecular surveillance and phylogenetic traits of Babesia bigemina and Babesia bovis in cattle (Bos taurus) and water buffaloes (Bubalus bubalis) from Colombia. Parasites Vectors 2018, 11, 510. [Google Scholar] [CrossRef]
- Paoletta, M.S.; Arias, L.L.; Fournière, S.; Guillemi, E.C.; Luciani, C.; Sarmiento, N.F.; Mosqueda, J.; Farber, M.D.; Wilkowsky, S.E. Epidemiology of Babesia, Anaplasma and Trypanosoma species using a new expanded reverse line blot hybridization assay. Ticks Tick Borne Dis. 2018, 9, 155–163. [Google Scholar] [CrossRef]
- Hakimi, H.; Yamagishi, J.; Kegawa, Y.; Kaneko, O.; Kawazu, S.; Asada, S. Establishment of transient and stable transfection systems for Babesia ovata. Parasites Vectors 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Terkawi, M.A.; Thekisoe, O.M.M.; Katsande, C.; Latif, A.A.; Mans, B.J.; Matthee, O.; Mkize, N.; Mabogoane, N.; Marais, F.; Yokoyama, N.; et al. Serological survey of Babesia bovis and Babesia bigemina in cattle in South Africa. Vet. Parasitol. 2011, 182, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.D. Babesiosis. In MSD Veterinary Manual; Merck & Co. Inc.: Kenilworth, NJ, USA, 2015; Available online: https://www.msdvetmanual.com/circulatory-system/blood-parasites/babesiosis (accessed on 2 September 2021).
- Aubry, P.; Geale, D.W. A review of Bovine anaplasmosis. Transbound. Emerg. Dis. 2011, 58, 1–30. [Google Scholar] [CrossRef]
- Belkahia, H.; Said, M.B.; Alberti, A.; Abdi, K.; Issaoui, Z.; Hattab, D.; Gharbi, M.; Messadi, L. First molecular survey and novel genetic variants’ identification of Anaplasma marginale, A. centrale and A. bovis in cattle from Tunisia. Infect. Genet. Evol. 2015, 34, 361–371. [Google Scholar] [CrossRef]
- Kocan, K.M.; Fuente, J.; Guglielmone, A.A.; Meléndez, R.D. Antigens and Alternatives for Control of Anaplasma marginale Infection in Cattle. Clin. Microbiol. Rev. 2003, 16, 698–712. [Google Scholar] [CrossRef] [Green Version]
- M’Ghirbi, Y.; Bèji, M.; Oporto, B.; Khrouf, F.; Hurtado, A.; Bouattour, A. Anaplasma marginale and A. phagocytophilum in cattle in Tunisia. Parasites Vectors 2016, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.P.; Zhang, B.; Wang, Y.H.; Xu, G.; Wang, X.; Ni, X.; Zhou, E.M. Molecular identification and characterization of Anaplasma capra and Anaplasma platys-like in Rhipicephalus microplus in Ankang, Northwest China. BMC Infect. Dis. 2019, 19, 434. [Google Scholar] [CrossRef] [PubMed]
- Zobba, R.; Anfossi, A.G.; Parpaglia, M.L.P.; Dore, G.M.; Chessa, B.; Spezzigu, A.; Rocca, S.; Visco, S.; Pittau, M.; Alberti, A. Molecular investigation and phylogeny of Anaplasma spp. in mediterranean ruminants reveal the presence of neutrophil-tropic strains closely related to A. platys. Appl. Environ. Microbiol. 2014, 80, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, W.M.A.; Ali, A.O.; Mahmoud, H.Y.A.H.; Omar, M.A.; Chatanga, E.; Salim, B.; Naguib, D.; Anders, J.L.; Nonaka, N.; Moustafa, M.A.M.; et al. Exploring Prokaryotic and Eukaryotic Microbiomes Helps in Detecting Tick-Borne Infectious Agents in the Blood of Camels. Pathogens 2021, 10, 351. [Google Scholar] [CrossRef]
- Said, M.B.; Belkahia, H.; El Mabrouk, N.; Saidani, M.; Alberti, A.; Zobba, R.; Cherif, A.; Mahjoub, T.; Bouattour, A.; Messadi, L. Anaplasma platys-like strains in ruminants from Tunisia. Infect. Genet. Evol. 2017, 49, 226–233. [Google Scholar] [CrossRef]
- André, M.R.; Calchi, A.C.; Herrera, H.M.; Zanatto, D.C.S.; Horta, B.C.L.S.; Tasso, J.B.; Ramos, I.S.A.; Mello, V.V.C.; Machado, R.Z. The co-infection with Ehrlichia minasensis, Anaplasma marginale and Anaplasma platys is not associated with anemia in beef cattle in the Brazilian Pantanal. Vet. Parasitol. Reg. Stud. Rep. 2020, 21, 100437. [Google Scholar] [CrossRef]
- Ray, R.; Albright, Z.C.; Wang, K. China-Latin America Economic Bulletin, 2021; Global Development Policy Center: Boston, MA, USA, 2021; Available online: https://www.bu.edu/gdp/2021/02/22/china-latin-america-economic-bulletin-2021/ (accessed on 2 September 2021).
- Mas, J.J.C.; Widdowson, M.A.; Cuéllar, A.M.; Ribera, H.; Walker, A.R. Risk of babesiosis and anaplasmosis in different ecological zones of Santa Cruz Department, Bolivia. Vet. Parasitol. 2000, 93, 29–38. [Google Scholar] [CrossRef]
- Carrique, J.J.; Morales, G.J.; Edelsten, M. Endemic Instability for Babesiosis and Anaplasmosis in Cattle in the Bolivian Chaco. Vet. J. 2000, 160, 162–164. [Google Scholar] [CrossRef]
- Masatani, T.; Hayashi, K.; Andoh, M.; Tateno, M.; Endo, Y.; Asada, M.; Kusakisako, K.; Tanaka, T.; Gokuden, M.; Hozumi, N.; et al. Detection and molecular characterization of Babesia, Theileria, and Hepatozoon species in hard ticks collected from Kagoshima, the southern region in Japan. Ticks Tick Borne Dis. 2017, 8, 581–587. [Google Scholar] [CrossRef]
- Chatanga, E.; Hayashida, K.; Muleya, W.; Kusakisako, K.; Moustafa, M.A.M.; Salim, B.; Katakura, K.; Sugimoto, C.; Nonaka, N.; Nakao, R. Genetic diversity and sequence polymorphism of two genes encoding Theileria parva antigens recognized by CD8+ T cells among vaccinated and unvaccinated cattle in Malawi. Pathogens 2020, 9, 334. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Roux, V.; Camicas, J.L.; Baradji, I.; Brouqui, P.; Raoult, D. Detection of ehrlichiae in African ticks by polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 707–708. [Google Scholar] [CrossRef]
- Qiu, Y.; Kaneko, C.; Kajihara, M.; Ngonda, S.; Simulundu, E.; Muleya, W.; Thu, M.J.; Hang’ombe, M.B.; Katakura, K.; Takada, A.; et al. Tick-borne haemoparasites and Anaplasmataceae in domestic dogs in Zambia. Ticks Tick Borne Dis. 2018, 9, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adenyo, C.; Ohya, K.; Qiu, Y.; Takashima, Y.; Ogawa, H.; Matsumoto, T.; Thu, M.J.; Sato, K.; Kawabata, H.; Katayama, Y.; et al. Bacterial and protozoan pathogens/symbionts in ticks infecting wild grasscutters (Thryonomys swinderianus) in Ghana. Acta Trop. 2020, 205, 105388. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Dueñez, J.J.; Chávez, O.T.; Jaramillo, A.M.M. Parasitological and molecular surveys reveal high rates of infection with vector-borne pathogens and clinical anemia signs associated with infection in cattle from two important livestock areas in Colombia. Ticks Tick Borne Dis. 2017, 8, 290–299. [Google Scholar] [CrossRef]
- Amorim, L.S.; Wenceslau, A.A.; Carvalho, F.S.; Carneiro, P.L.S.; Albuquerque, G. Bovine babesiosis and anaplasmosis complex: Diagnosis and evaluation of the risk factors from Bahia, Brazil. Rev. Bras. Parasitol. Veterinária 2014, 23, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.J.; Yamasaki, M.; Nakamura, K.; Sasaki, N.; Murakami, M.; Kumara, B.; Rajapakshage, W.; Ohta, H.; Maede, Y.; Takiguchi, M. Development and characterization of a strain of Babesia gibsoni resistant to diminazene aceturate in vitro. J. Vet. Med. Sci. 2010, 72, 765–771. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, M.; Watanabe, N.; Idaka, N.; Yamamori, T.; Otsuguro, K.; Uchida, N.; Iguchi, A.; Ohta, H.; Takiguchi, M. Intracellular diminazene aceturate content and adenosine incorporation in diminazene aceturate-resistant Babesia gibsoni isolate in vitro. Exp. Parasitol. 2017, 183, 92–98. [Google Scholar] [CrossRef]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Genotypic analysis of piroplasms and associated pathogens from ticks infesting cattle in Korea. Microorganisms 2020, 8, 728. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Ouh, I.O.; Lee, H.; Geraldino, P.J.L.; Rhee, M.H.; Kwon, O.D.; Kwak, D. Differential identification of Anaplasma in cattle and potential of cattle to serve as reservoirs of Anaplasma capra, an emerging tick-borne zoonotic pathogen. Vet. Microbiol. 2018, 226, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Tumwebaze, M.A.; Lee, S.-H.; Moumouni, P.F.A.; Mohammed-Geba, K.; Sheir, S.K.; Galal-Khallaf, A.; El Latif, H.M.A.; Morsi, D.S.; Bishr, N.M.; Galon, E.M.; et al. Parasitology International First detection of Anaplasma ovis in sheep and Anaplasma platys-like variants from cattle in Menoufia governorate, Egypt. Parasitol. Int. 2020, 78, 102150. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.H.L.; Tiawsirisup, S.; Kaewthamasorn, M. Molecular detection and genetic characterization of Anaplasma marginale and Anaplasma platys-like (Rickettsiales: Anaplasmataceae) in water buffalo from eight provinces of Thailand. BMC Vet. Res. 2020, 16, 380. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, M.; Davoust, B.; Seghir, M.; Fenollar, F.; Raoult, D.; Mediannikov, O. Comparative Immunology, Microbiology and Infectious Diseases Development of a new PCR-based assay to detect Anaplasmataceae and the first report of Anaplasma phagocytophilum and Anaplasma platys in cattle from Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2015, 39, 39–45. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Liu, Z.; Liu, J.; Yang, J.; Li, Q.; Li, Y.; Luo, J.; Yin, H. Molecular Survey of Anaplasma and Ehrlichia of Red Deer and Sika Deer in Gansu, China in 2013. Transbound. Emerg. Dis. 2016, 63, e228–e236. [Google Scholar] [CrossRef] [PubMed]
- Janer, E.C.; Rifran, L.; Piaggio, J.; Gil, A.; Miller, R.J.; Schumaker, T.T.S. In vitro tests to establish LC 50 and discriminating concentrations for fipronil against Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) and their standardization. Vet. Parasitol. 2009, 162, 120–128. [Google Scholar] [CrossRef]
- Bandara, K.M.U.J.; Karunaratne, S.H.P.P. Mechanisms of acaricide resistance in the cattle tick Rhipicephalus (Boophilus) microplus in Sri Lanka. Pestic. Biochem. Physiol. 2017, 139, 68–72. [Google Scholar] [CrossRef]
- Piper, E.K.; Jackson, L.A.; Ohmann, H.B.; Gondro, C.; Tabor, A.E.L.; Jonsson, N.N. Tick-susceptible Bos taurus cattle display an increased cellular response at the site of larval Rhipicephalus (Boophilus) microplus attachment, compared with tick-resistant Bos indicus cattle. Int. J. Parasitol. 2010, 40, 431–441. [Google Scholar] [CrossRef]
- Giglioti, R.; Oliveira, H.N.; Santana, C.H.; Ibelli, A.M.G.; Néo, T.A.; Bilhassi, T.B.; Rabelo, M.D.; Machado, R.Z.; Brito, L.G.; Oliveira, M.C.S. Babesia bovis and Babesia bigemina infection levels estimated by qPCR in Angus cattle from an endemic area of São Paulo state, Brazil. Ticks Tick Borne Dis. 2016, 7, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Guglielmone, A.A. Epidemiology of babesiosis and anaplasmosis in South and Central America. Vet. Parasitol. 1995, 57, 109–119. [Google Scholar] [CrossRef]
- Hosary, A.L.; Vectors, P.; Hosary, A.A.L.; Răileanu, C.; Tauchmann, O.; Fischer, S.; Nijhof, A.M.; Silaghi, C. Epidemiology and genotyping of Anaplasma marginale and co-infection with piroplasms and other Anaplasmataceae in cattle and buffaloes from Egypt. Parasit. Vectors 2020, 13, 495. [Google Scholar] [CrossRef]
- Hailemariam, Z.; Krücken, J.; Baumann, M.; Ahmed, J.S.; Clausen, P.H.; Nijhof, A.M. Molecular detection of tick-borne pathogens in cattle from Southwestern Ethiopia. PLoS ONE 2017, 12, e0188248. [Google Scholar] [CrossRef] [Green Version]
- Ogden, N.H.; Lindsay, L.R. Effects of Climate and Climate Change on Vectors and Vector-Borne Diseases: Ticks Are Different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Target Gene | Organism | Annealing Temperature (°C) | Purpose | Reference |
|---|---|---|---|---|---|---|
| EHR16SD | GGTACCYACAGAAGAAGTCC | 16S rDNA | Anaplasmataceae | 55 | PCR | [26] |
| EHR16SR | TAGCACTCATCGTTTACAGC | |||||
| Pglt-F | ATGAWAGAAAAWGCTGTTTT | gltA | Anaplasma | 60 | Sequencing | [16] |
| Pglt-R1 | TCATGRTCTGCATGCATKATG | |||||
| Pglt-R2 | CATGCATKATGAARATMGCAT | |||||
| Pglt-L-F1 | GATGCWCATCCYATSGCMATGT | gltA | Anaplasma | 60 | Sequencing | [16] |
| Pglt-L-F2 | CGTGMTSGCTATAGCGMAART | |||||
| Pglt-L-R | TCAYACCATTGDGAYRCCCAT | |||||
| Pgro-F1 | TTGATCATCGCTGAAGACGT | groEL | Anaplasma | 60 | Sequencing | [16] |
| Pgro-F2 | ACTCTCGTCTTGAACAAGCT | |||||
| Pgro-R | CCACTCTGTCTTTACGCTCT | |||||
| Pgro-F-F | AAATGKCAAATACGGTWGTC | groEL | Anaplasma | 60 | Sequencing | [16] |
| Pgro-F-R1 | ACAACACCTTCCTCKACAGC | |||||
| Pgro-F-R2 | CTGKCTTTRCGYTCTTTAACTTC | |||||
| Pgro-L-F1 | GAYGGTATGCAGTTTGATCGCG | groEL | Anaplasma | 60 | Sequencing | [16] |
| Pgro-L-F2 | ATGCAGTTTGATCGCGGWTATC | |||||
| Pgro-L-R | CAGCRAGGTCGAAYGCAATAC | |||||
| BTH 1st F | GTGAAACTGCGAATGGCTCATTAC | 18S rDNA | Babesia | 55 | PCR | [24] |
| BTH 1st R | AAGTGATAAGGTTCACAAAACTTCCC | Theileria | ||||
| BTH 2nd F | GGCTCATTACAACAGTTATAGTTTATTTG | Hepatozoon | ||||
| BTH 2nd R | CGGTCCGAATAATTCACCGGAT | |||||
| BTH18SinterF | ATTTTCCGACTCCTTCAGCA | 18S rDNA | Babesia | NA | Sequencing | This study |
| BTH18SinterR | AACTAAGAACGGCCATGCAC |
| Region | Breed | Number | Babesia | Anaplasma (EHR PCR) | Anaplasma (groEL) | Anaplasma (gltA) | Co-Infection |
|---|---|---|---|---|---|---|---|
| University Farm 1 | Holstein | 10 | 5 (50%) | 8 (80%) | 8 (80%) | 5 (50%) | 5 (50%) |
| University Farm 2 | Nelore | 10 | 2 (20%) | 2 (20%) | 1 (10%) | - | 2 (20%) |
| Gyr | 10 | - | 6 (60%) | 6 (60%) | 3 (30%) | - | |
| Gyrolando | 8 | 1 (13%) | 7 (88%) | 5 (63%) | 1 (13%) | 1 (13%) | |
| University Farm 3 | Criollo | 22 | 1 (5%) | 7 (32%) | 2 (9%) | 5 (23%) | - |
| Private Farm 1 | Nelore | 10 | - | - | - | - | - |
| Mestizo | 2 | - | - | - | - | - | |
| Senepol | 10 | - | - | - | - | - | |
| Private Farm 2 | Nelore | 11 | - | 1 (9%) | - | - | - |
| Neloblanca | 1 | - | - | - | - | - | |
| Private Farm 3 | Nelore | 10 | - | 1 (10%) | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogata, S.; Pereira, J.A.C.; Jhonny, L.V.A.; Carolina, H.P.G.; Matsuno, K.; Orba, Y.; Sawa, H.; Kawamori, F.; Nonaka, N.; Nakao, R. Molecular Survey of Babesia and Anaplasma Infection in Cattle in Bolivia. Vet. Sci. 2021, 8, 188. https://doi.org/10.3390/vetsci8090188
Ogata S, Pereira JAC, Jhonny LVA, Carolina HPG, Matsuno K, Orba Y, Sawa H, Kawamori F, Nonaka N, Nakao R. Molecular Survey of Babesia and Anaplasma Infection in Cattle in Bolivia. Veterinary Sciences. 2021; 8(9):188. https://doi.org/10.3390/vetsci8090188
Chicago/Turabian StyleOgata, Shohei, Juan Antonio Cristian Pereira, Loza Vega Ariel Jhonny, Herbas Perez Gladys Carolina, Keita Matsuno, Yasuko Orba, Hirofumi Sawa, Fumihiko Kawamori, Nariaki Nonaka, and Ryo Nakao. 2021. "Molecular Survey of Babesia and Anaplasma Infection in Cattle in Bolivia" Veterinary Sciences 8, no. 9: 188. https://doi.org/10.3390/vetsci8090188
APA StyleOgata, S., Pereira, J. A. C., Jhonny, L. V. A., Carolina, H. P. G., Matsuno, K., Orba, Y., Sawa, H., Kawamori, F., Nonaka, N., & Nakao, R. (2021). Molecular Survey of Babesia and Anaplasma Infection in Cattle in Bolivia. Veterinary Sciences, 8(9), 188. https://doi.org/10.3390/vetsci8090188