Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
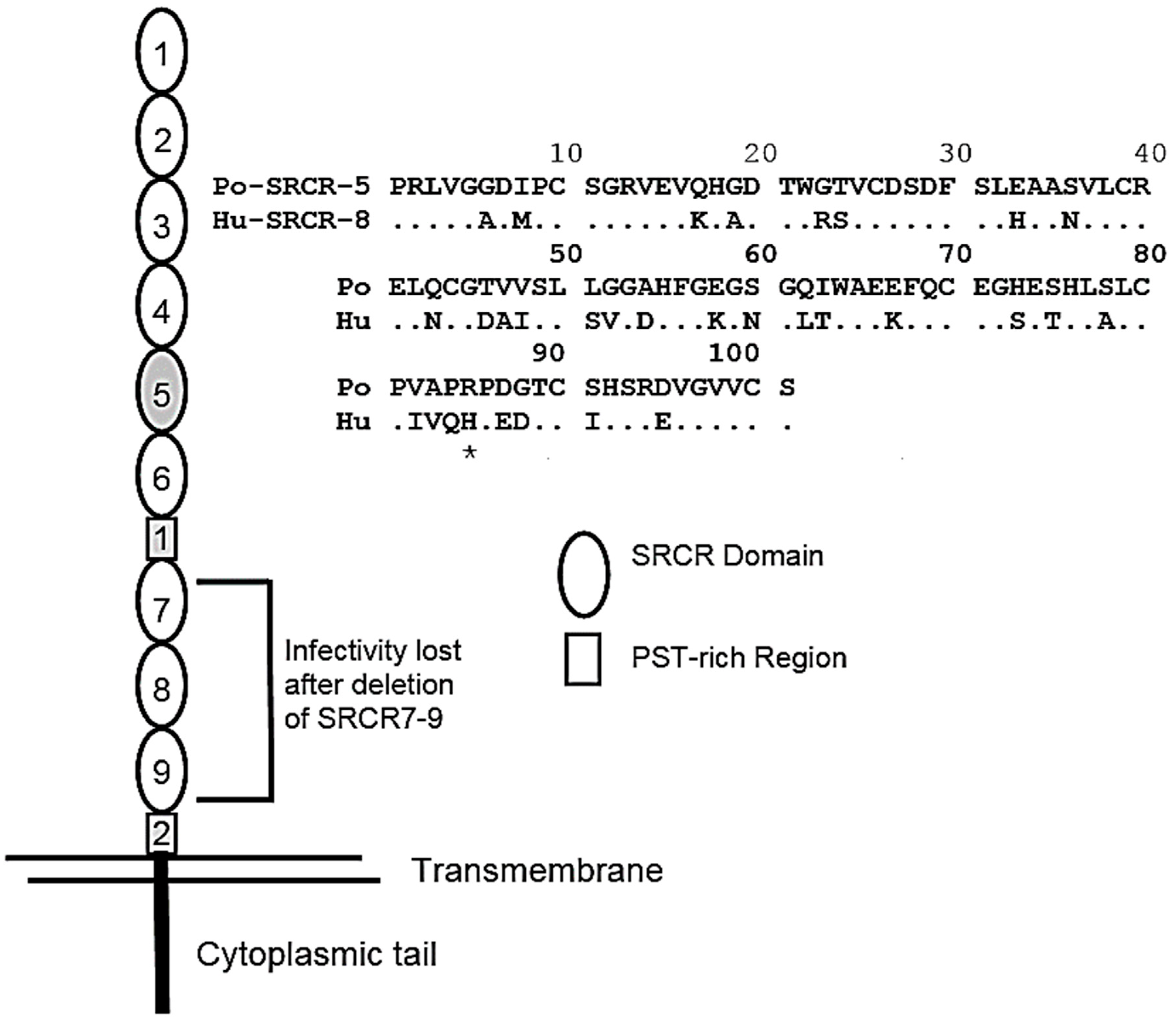
2. CD163 as the Sole Receptor for PRRSV
3. The Interaction between pCD163 and PRRSV Glycoproteins
4. PRRSV Neutralization and Escape from Homologous and broadly Neutralizing Activity through the GP5/M Heterodimer
5. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gulyaeva, A.A.; Lauber, C.; Samborskiy, D.V.; Leontovich, A.M.; Sidorov, I.A.; Gorbalenya, A.E. Evolutionary based classification of genomic diversity of nidoviruses connects metagenomics and experimental research. In Proceedings of the 14th International Nidovirus Symposium, Kansas City, MO, USA, 4–9 June 2017. [Google Scholar]
- Plagemann, P.G.W. Porcine Reproductive and Respiratory Syndrome Virus: Origin Hypothesis. Emerg. Infect. Dis. 2003, 9, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Matanin, B.M.; Huang, Y.; Meng, X.J.; Zhang, C. Purification of the Major Envelop Protein GP5 of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) from Native Virions. J. Virol. Methods 2008, 147, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.R.; Griggs, T.F.; Gnanandarajah, J.; Murtaugh, M.P. Novel Structural Protein in Porcine Reproductive and Respiratory Syndrome Virus Encoded by an Alternative ORF5 Present in All Arteriviruses. J. Gen. Virol. 2011, 92, 1107–1116. [Google Scholar] [CrossRef]
- Wu, W.H.; Fang, Y.; Farwell, R.; Steffen-Bien, M.; Rowland, R.R.R.; Christopher-Hennings, J.; Nelson, E.A. A 10-KDa Structural Protein of Porcine Reproductive and Respiratory Syndrome Virus Encoded by ORF2b. Virology 2001, 287, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Kappes, M.A.; Miller, C.L.; Faaberg, K.S. Highly Divergent Strains of Porcine Reproductive and Respiratory Syndrome Virus Incorporate Multiple Isoforms of Nonstructural Protein 2 into Virions. J. Virol. 2013, 87, 13456–13465. [Google Scholar] [CrossRef] [PubMed]
- Spilman, M.S.; Welbon, C.; Nelson, E.; Dokland, T. Cryo-Electron Tomography of Porcine Reproductive and Respiratory Syndrome Virus: Organization of the Nucleocapsid. J. Gen. Virol. 2009, 90, 527–535. [Google Scholar] [CrossRef]
- Das, P.B.; Vu, H.L.X.; Dinh, P.X.; Cooney, J.L.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Glycosylation of Minor Envelope Glycoproteins of Porcine Reproductive and Respiratory Syndrome Virus in Infectious Virus Recovery, Receptor Interaction, and Immune Response. Virology 2011, 410, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Ansari, I.H.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Influence of N-Linked Glycosylation of Porcine Reproductive and Respiratory Syndrome Virus GP5 on Virus Infectivity, Antigenicity, and Ability to Induce Neutralizing Antibodies Israrul. J. Virol. 2006, 80, 3994–4004. [Google Scholar] [CrossRef] [PubMed]
- Van Breedam, W.; Van Gorp, H.; Zhang, J.Q.; Crocker, P.R.; Delputte, P.L.; Nauwynck, H.J. The M/GP5glycoprotein Complex of Porcine Reproductive and Respiratory Syndrome Virus Binds the Sialoadhesin Receptor in a Sialic Acid-Dependent Manner. PLoS Pathog. 2010, 6, e1000730. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Vanderheijden, N.; Nauwynck, H.J.; Pensaert, M.B. Involvement of the Matrix Protein in Attachment of Porcine Reproductive and Respiratory Syndrome Virus to a Heparinlike Receptor on Porcine Alveolar Macrophages. J. Virol. 2002, 76, 4312–4320. [Google Scholar] [CrossRef]
- Vanderheijden, N.; Delputte, P.L.; Favoreel, H.W.; Vandekerckhove, J.; Van Damme, J.; van Woensel, P.A.; Nauwynck, H.J. Involvement of Sialoadhesin in Entry of Porcine Reproductive and Respiratory Syndrome Virus into Porcine Alveolar Macrophages. J. Virol. 2003, 77, 8207–8215. [Google Scholar] [CrossRef]
- Calvert, J.G.; Slade, D.E.; Shields, S.L.; Jolie, R.; Mannan, R.M.; Ankenbauer, R.G.; Welch, S.-K.W. CD163 Expression Confers Susceptibility to Porcine Reproductive and Respiratory Syndrome Viruses. J. Virol. 2007, 81, 7371–7379. [Google Scholar] [CrossRef] [PubMed]
- Welch, S.K.W.; Calvert, J.G. A Brief Review of CD163 and Its Role in PRRSV Infection. Virus Res. 2010, 154, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Prather, R.S.; Rowland, R.R.R.; Ewen, C.; Trible, B.; Kerrigan, M.; Bawa, B.; Teson, J.M.; Mao, J.; Lee, K.; Samuel, M.S.; et al. An Intact Sialoadhesin (Sn/SIGLEC1/CD169) Is Not Required for Attachment/Internalization of the Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2013, 87, 9538–9546. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Christiaens, I.; Yang, B.; Trus, I.; Devriendt, B.; Cui, T.; Wei, R.; Nauwynck, H.J. Preferential Use of Siglec-1 or Siglec-10 by Type 1 and Type 2 PRRSV Strains to Infect PK15S1-CD163 and PK15S10-CD163 Cells. Vet. Res. 2018, 49, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.M.; Rowland, R.R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-Edited Pigs Are Protected from Porcine Reproductive and Respiratory Syndrome Virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Burkard, C.; Lillico, S.G.; Reid, E.; Jackson, B.; Mileham, A.J.; Ait-Ali, T.; Whitelaw, C.B.A.; Archibald, A.L. Precision Engineering for PRRSV Resistance in Pigs: Macrophages from Genome Edited Pigs Lacking CD163 SRCR5 Domain Are Fully Resistant to Both PRRSV Genotypes While Maintaining Biological Function. PLoS Pathog. 2017, 13, e1006206. [Google Scholar] [CrossRef] [PubMed]
- Van Gorp, H.; Van Breedam, W.; Van Doorsselaere, J.; Delputte, P.L.; Nauwynck, H.J. Identification of the CD163 Protein Domains Involved in Infection of the Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2010, 84, 3101–3105. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D.; Bardot, R.; Whitworth, K.M.; Trible, B.R.; Fang, Y.; Mileham, A.; Kerrigan, M.A.; Samuel, M.S.; Prather, R.S.; Rowland, R.R.R. Replacement of Porcine CD163 Scavenger Receptor Cysteine-Rich Domain 5 with a CD163-Like Homolog Confers Resistance of Pigs to Genotype 1 but Not Genotype 2 Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2017, 91, e01521-16. [Google Scholar] [CrossRef]
- Ma, H.; Jiang, L.; Qiao, S.; Zhi, Y.; Chen, X.-X.; Yang, Y.; Huang, X.; Huang, M.; Li, R.; Zhang, G.-P. The Crystal Structure of the Fifth Scavenger Receptor Cysteine-Rich Domain of Porcine CD163 Reveals an Important Residue Involved in Porcine Reproductive and Respiratory Syndrome Virus Infection. J. Virol. 2017, 91, e01897-16. [Google Scholar] [CrossRef]
- Burkard, C.; Opriessnig, T.; Mileham, A.J.; Stadejek, T.; Ait-Ali, T.; Lillico, S.G.; Whitelaw, C.B.A.; Archibald, A.L. Pigs Lacking the Scavenger Receptor Cysteine-Rich Domain 5 of CD163 Are Resistant to PRRSV-1 Infection. J. Virol. 2018, 92, e00415-18. [Google Scholar] [CrossRef]
- Das, P.B.; Dinh, P.X.; Ansari, I.H.; de Lima, M.; Osorio, F.A.; Pattnaik, A.K. The Minor Envelope Glycoproteins GP2a and GP4 of Porcine Reproductive and Respiratory Syndrome Virus Interact with the Receptor CD163. J. Virol. 2010, 84, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, M.; Costers, S.; Van Breedam, W.; Geldhof, M.F.; Van Doorsselaere, J.; Nauwynck, H.J. A Variable Region in GP4 of European-Type Porcine Reproductive and Respiratory Syndrome Virus Induces Neutralizing Antibodies Against Homologous but Not Heterologous Virus Strains. Viral Immunol. 2010, 23, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Pattnaik, A.K.; Song, C.; Yoo, D.; Li, G. Glycosyl-Phosphatidylinositol (GPI)-Anchored Membrane Association of the Porcine Reproductive and Respiratory Syndrome Virus GP4 Glycoprotein and Its Co-Localization with CD163 in Lipid Rafts. Virology 2012, 424, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cui, J.; Wang, J.; Sun, Y.; Ji, C.; Song, R.; Zeng, Y.; Pan, H.; Sheng, J.; Zhang, G.; et al. Phages Bearing Specific Peptides with Affinity for Porcine Reproductive and Respiratory Syndrome Virus GP4 Protein Prevent Cell Penetration of the Virus. Vet. Microbiol. 2018, 22, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Kimpston-Burkgren, K.; Correas, I.; Osorio, F.A.; Steffen, D.; Pattnaik, A.K.; Fang, Y.; Vu, H.L.X. Relative Contribution of Porcine Reproductive and Respiratory Syndrome Virus Open Reading Frames 2–4 to the Induction of Protective Immunity. Vaccine 2017, 35, 4408–4413. [Google Scholar] [CrossRef] [PubMed]
- Costers, S.; Lefebvre, D.J.; van Doorsselaere, J.; Vanhee, M.; Delputte, P.L.; Nauwynck, H.J. GP4 of Porcine Reproductive and Respiratory Syndrome Virus Contains a Neutralizing Epitope That Is Susceptible to Immunoselection in Vitro. Arch. Virol. 2010, 155, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Oleksiewicz, M.B.; Botner, A.; Toft, P.; Normann, P.; Storgaard, T. Epitope map- ping porcine reproductive and respiratory syndrome virus by phage display: The nsp2 fragment of the replicase polyprotein contains a cluster of B-cell epitopes. J. Virol. 2001, 75, 3277–3290. [Google Scholar] [CrossRef]
- Popescu, L.N.; Trible, B.R.; Chen, N.; Rowland, R.R.R. GP5 of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) as a Target for Homologous and Broadly Neutralizing Antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef]
- Robinson, S.R.; Li, J.; Nelson, E.A.; Murtaugh, M.P. Broadly Neutralizing Antibodies against the Rapidly Evolving Porcine Reproductive and Respiratory Syndrome Virus. Virus Res. 2015, 203, 56–65. [Google Scholar] [CrossRef]
- Trible, B.R.; Popescu, L.N.; Monday, N.; Calvert, J.G.; Rowland, R.R.R. A Single Amino Acid Deletion in the Matrix Protein of Porcine Reproductive and Respiratory Syndrome Virus Confers Resistance to a Polyclonal Swine Antibody with Broadly Neutralizing Activity. J. Virol. 2015, 89, 6515–6520. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of Neutralizing and Nonneutralizing Epitopes in the Porcine Reproductive and Respiratory Syndrome Virus GP5 Ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef] [PubMed]
- Plagemann, P.G.W.; Rowland, R.R.R.; Faaberg, K.S. The Primary Neutralization Epitope of Porcine Respiratory and Reproductive Syndrome Virus Strain VR-2332 Is Located in the Middle of the GP5 Ectodomain. Arch. Virol. 2002, 147, 2327–2347. [Google Scholar] [CrossRef] [PubMed]
- Plagemann, P.G.W. The Primary GP5 Neutralization Epitope of North American Isolates of Porcine Reproductive and Respiratory Syndrome Virus. Vet. Immunol. Immunopathol. 2004, 102, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Delisle, B.; Gagnon, C.A.; Lambert, M.-È.; D’Allaire, S. Porcine Reproductive and Respiratory Syndrome Virus Diversity of Eastern Canada Swine Herds in a Large Sequence Dataset Reveals Two Hypervariable Regions under Positive Selection. Infect. Genet. Evol. 2012, 12, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Plagemann, P.G.W. Complexity of the Single Linear Neutralization Epitope of the Mouse Arterivirus Lactate Dehydrogenase-Elevating Virus. Virology 2001, 290, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Faaberg, K.S.; Hocker, J.D.; Erdman, M.M.; Harris, D.L.H.; Nelson, E.A.; Torremorell, M.; Plagemann, P.G.W. Neutralizing Antibody Responses of Pigs Infected with Natural GP5 N-Glycan Mutants of Porcine Reproductive and Respiratory Syndrome Virus. Viral Immunol. 2006, 19, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.L.X.; Kwon, B.; Yoon, K.-J.; Laegreid, W.W.; Pattnaik, A.K.; Osorio, F.A. Immune Evasion of Porcine Reproductive and Respiratory Syndrome Virus through Glycan Shielding Involves Both Glycoprotein 5 as Well as Glycoprotein 3. J. Virol. 2011, 85, 5555–5564. [Google Scholar] [CrossRef]
- Meulenberg, J.J.; van Nieuwstadt, A.P.; van Essen-Zandbergen, A.; Langeveld, J.P. Posttranslational processing and identification of a neutralization domain of the GP4 protein encoded by ORF4 of Lelystad virus. J. Virol. 1997, 71, 6061–6067. [Google Scholar]
- Vanhee, M.; Van Breedam, W.; Costers, S.; Geldhof, M.; Noppe, Y.; Nauwynck, H. Characterization of antigenic regions in the porcine reproductive and respiratory syndrome virus by the use of peptide-specific serum antibodies. Vaccine 2011, 29, 4794–4804. [Google Scholar] [CrossRef]
- Zhou, Y.-J.; An, T.-Q.; He, Y.-X.; Liu, J.-X.; Qiu, H.-J.; Wang, Y.-F.; Tong, G. Antigenic structure analysis of glycosylated protein 3 of porcine reproductive and respiratory syndrome virus. Virus Res. 2006, 118, 98–104. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoian, A.M.M.; Rowland, R.R.R. Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization. Vet. Sci. 2019, 6, 9. https://doi.org/10.3390/vetsci6010009
Stoian AMM, Rowland RRR. Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization. Veterinary Sciences. 2019; 6(1):9. https://doi.org/10.3390/vetsci6010009
Chicago/Turabian StyleStoian, Ana M. M., and Raymond R. R. Rowland. 2019. "Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization" Veterinary Sciences 6, no. 1: 9. https://doi.org/10.3390/vetsci6010009
APA StyleStoian, A. M. M., & Rowland, R. R. R. (2019). Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization. Veterinary Sciences, 6(1), 9. https://doi.org/10.3390/vetsci6010009