
Genome Mining Reveals a Sactipeptide Biosynthetic Cluster in Staphylococcus pseudintermedius



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates Included in the Study
2.2. Genome Mining by antiSMASH and BAGEL4
2.3. Sequence Analysis
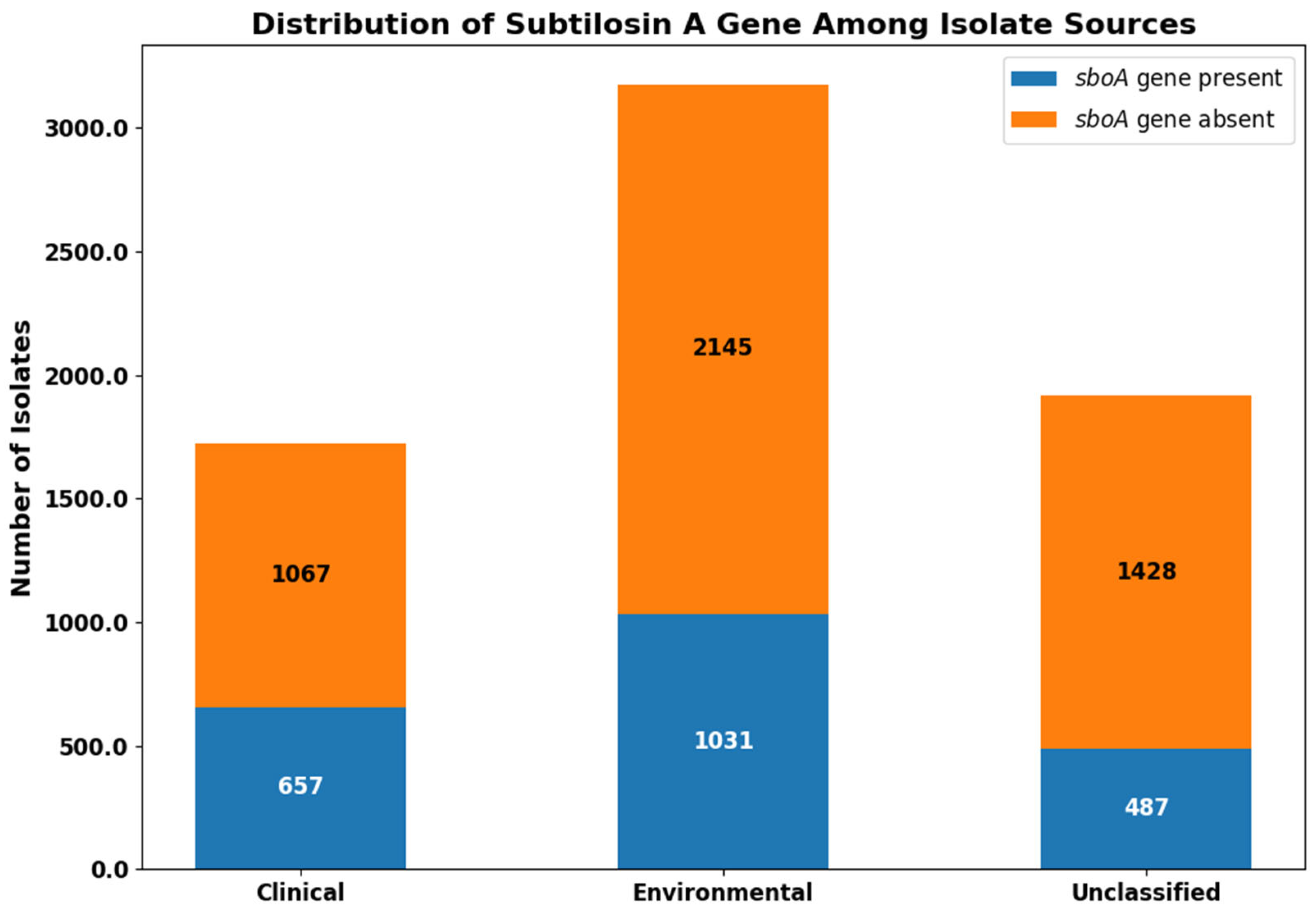
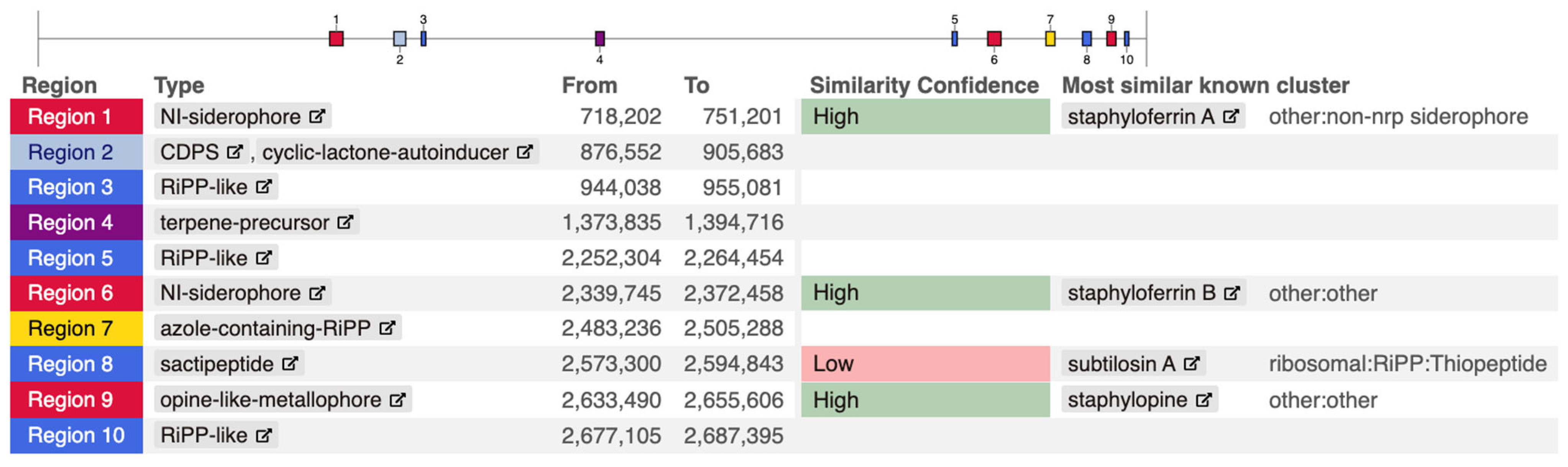
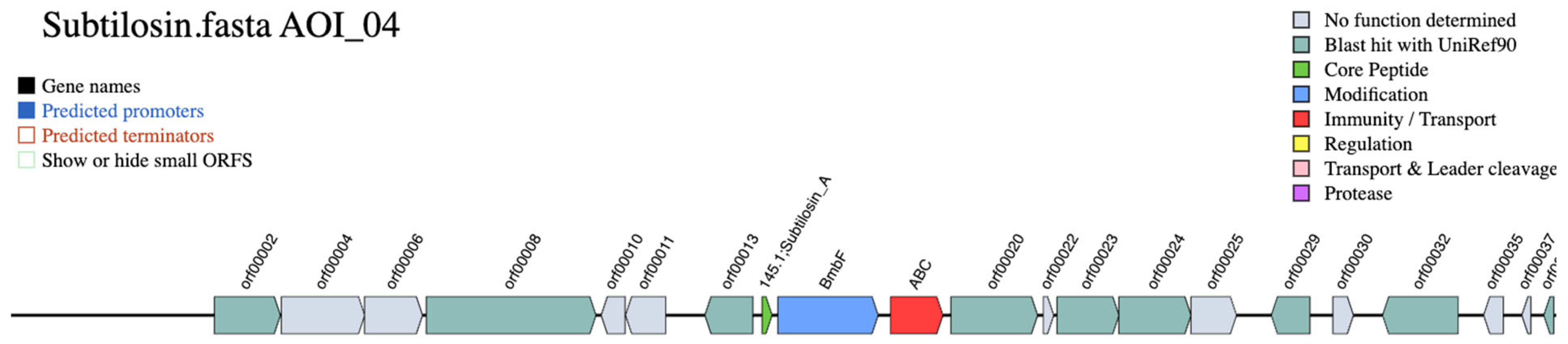
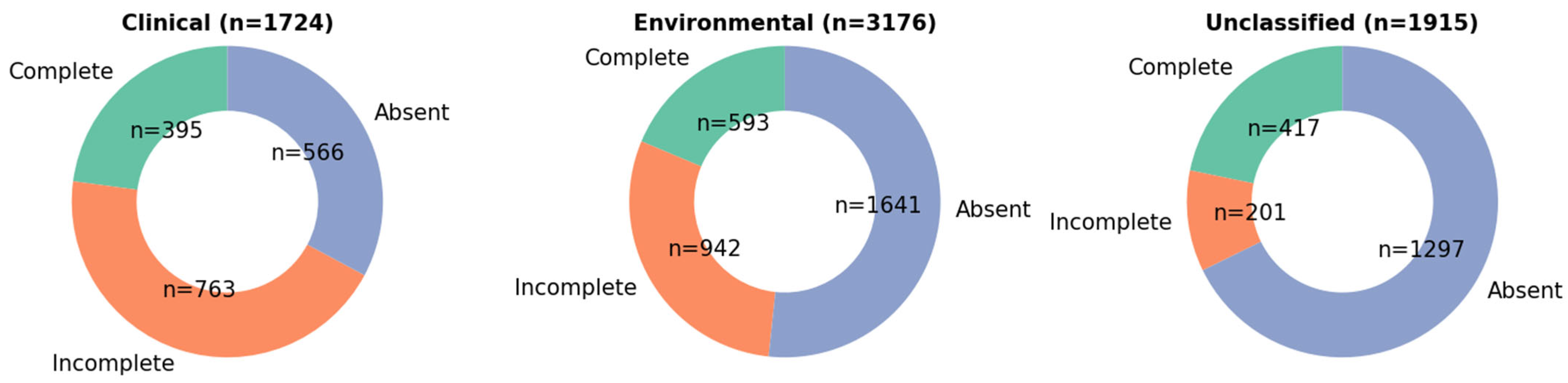
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shelburne, C.E.; An, F.Y.; Dholpe, V.; Ramamoorthy, A.; Lopatin, D.E.; Lantz, M.S. The spectrum of antimicrobial activity of the bacteriocin subtilosin A. J. Antimicrob. Chemother. 2007, 59, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Nakamura, A.; Kojima, S. Crystal structure of the AlbEF complex involved in subtilosin A biosynthesis. Structure 2022, 30, 1637–1646.E3. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Yan, L.Z.; Vederas, J.C.; Zuber, P. Genes of the sbo-alb locus of Bacillus subtilis are required for production of the antilisterial bacteriocin subtilosin. J. Bacteriol. 1999, 181, 7346–7355. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Oliveira, A.; Manageiro, V.; Caniça, M.; Contente, D.; Capita, R.; Alonso-Calleja, C.; Carvalho, I.; Capelo, J.L.; Igrejas, G.; et al. Clonal Diversity and Antimicrobial Resistance of Methicillin-Resistant Staphylococcus pseudintermedius Isolated from Canine Pyoderma. Microorganisms 2021, 9, 482. [Google Scholar] [CrossRef]
- Viñes, J.; Verdejo, M.Á.; Horvath, L.; Vergara, A.; Vila, J.; Francino, O.; Morata, L.; Espasa, M.; Casals-Pascual, C.; Soriano, À.; et al. Isolation of Staphylococcus pseudintermedius in Immunocompromised Patients from a Single Center in Spain: A Zoonotic Pathogen from Companion Animals. Microorganisms 2024, 12, 1695. [Google Scholar] [CrossRef]
- Paul, N.C.; Moodley, A.; Ghibaudo, G.; Guardabassi, L. Carriage of methicillin-resistant Staphylococcus pseudintermedius in small animal veterinarians: Indirect evidence of zoonotic transmission. Zoonoses Public Health 2011, 58, 533–539. [Google Scholar] [CrossRef]
- Usui, M.; Sabala, R.F.; Morita, S.; Fukuda, A.; Tsuyuki, Y.; Torii, K.; Nakamura, Y.; Okamura, K.; Komatsu, T.; Sasaki, J.; et al. Antimicrobial susceptibility and genetic diversity of staphylococcus pseudintermedius isolated from companion animals and human clinical patients in Japan: Potential zoonotic implications. J. Glob. Antimicrob. Resist. 2025, 42, 66–72. [Google Scholar] [CrossRef]
- Moses, I.B.; Santos, F.F.; Gales, A.C. Human Colonization and Infection by Staphylococcus pseudintermedius: An Emerging and Underestimated Zoonotic Pathogen. Microorganisms 2023, 11, 581. [Google Scholar] [CrossRef]
- Sayem, S.A.J.; Lee, G.Y.; Abbas, M.A.; Park, S.C.; Lee, S.J. Pharmacodynamic Profiling of Amoxicillin: Targeting Multidrug-Resistant Gram-Positive Pathogens Staphylococcus aureus and Staphylococcus pseudintermedius in Canine Clinical Isolates. Antibiotics 2025, 14, 99. [Google Scholar] [CrossRef]
- Grist, L.F.; Brown, A.; Fitzpatrick, N.; Mariano, G.; La Ragione, R.M.; Van Vliet, A.H.M.; Mehat, J.W. Global phylogenomic analysis of Staphylococcus pseudintermedius reveals genomic and prophage diversity in multidrug-resistant lineages. Microb. Genom. 2025, 11, 001369. [Google Scholar] [CrossRef]
- Myrenås, M.; Pedersen, K.; Windahl, U. Genomic Analyses of Methicillin-Resistant Staphylococcus pseudintermedius from Companion Animals Reveal Changing Clonal Populations, Multidrug Resistance, and Virulence. Antibiotics 2024, 13, 962. [Google Scholar] [CrossRef] [PubMed]
- Phophi, L.; Abouelkhair, M.; Jones, R.; Henton, M.; Qekwana, D.N.; Kania, S.A.; Zhang, K. The molecular epidemiology and antimicrobial resistance of Staphylococcus pseudintermedius canine clinical isolates submitted to a veterinary diagnostic laboratory in South Africa. PLoS ONE 2023, 18, e0290645. [Google Scholar] [CrossRef] [PubMed]
- Phophi, L.; Abouelkhair, M.A.; Jones, R.; Zehr, J.; Kania, S.A. Temporal changes in antibiotic resistance and population structure of methicillin-resistant Staphylococcus pseudintermedius between 2010 and 2021 in the United States. Comp. Immunol. Microbiol. Infect. Dis. 2023, 100, 102028. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Vader, L.; Szenei, J.; Reitz, Z.L.; E Augustijn, H.; Cediel-Becerra, J.D.D.; de Crécy-Lagard, V.; A Koetsier, R.; E Williams, S.; et al. antiSMASH 8.0: Extended gene cluster detection capabilities and analyses of chemistry, enzymology, and regulation. Nucleic Acids Res. 2025, gkaf334. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253-e52. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Amrouche, T.; Noll, K.S.; Wang, Y.; Huang, Q.; Chikindas, M.L. Antibacterial Activity of Subtilosin Alone and Combined with Curcumin, Poly-Lysine and Zinc Lactate Against Listeria monocytogenes Strains. Probiotics Antimicrob. Proteins 2010, 2, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Zehr, J.D.; Sun, Q.; Ceres, K.; Merrill, A.; Tyson, G.H.; Ceric, O.; Guag, J.; Pauley, S.; McQueary, H.C.; Sams, K.; et al. Population and pan-genomic analyses of Staphylococcus pseudintermedius identify geographic distinctions in accessory gene content and novel loci associated with AMR. Appl. Environ. Microbiol. 2025, 91, e00010-25. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Hehn, R.; Zuber, P. Mutational analysis of the sbo-alb locus of Bacillus subtilis: Identification of genes required for subtilosin production and immunity. J. Bacteriol. 2000, 182, 3266–3273. [Google Scholar] [CrossRef]
- de Freire Bastos, M.D.C.; de Farias, F.M.; Fagundes, P.C.; Coelho, M.L.V. Staphylococcins: An update on antimicrobial peptides produced by staphylococci and their diverse potential applications. Appl. Microbiol. Biotechnol. 2020, 104, 10339–10368. [Google Scholar] [CrossRef]
- Fernández-Fernández, R.; Elsherbini, A.M.A.; Lozano, C.; Martínez, A.; de Toro, M.; Zarazaga, M.; Peschel, A.; Krismer, B.; Torres, C. Genomic Analysis of Bacteriocin-Producing Staphylococci: High Prevalence of Lanthipeptides and the Micrococcin P1 Biosynthetic Gene Clusters. Probiotics Antimicrob. Proteins 2025, 17, 159–174. [Google Scholar] [CrossRef]
- Wladyka, B.; Piejko, M.; Bzowska, M.; Pieta, P.; Krzysik, M.; Mazurek, Ł.; Guevara-Lora, I.; Bukowski, M.; Sabat, A.J.; Friedrich, A.W.; et al. A peptide factor secreted by Staphylococcus pseudintermedius exhibits properties of both bacteriocins and virulence factors. Sci. Rep. 2015, 5, 14569. [Google Scholar] [CrossRef]
- Hong, J.; Kim, J.; Quan, L.H.; Heu, S.; Roh, E. Purification and Characterization of Pasteuricin Produced by Staphylococcus pasteuri RSP-1 and Active against Multidrug-Resistant Staphylococcus aureus. J. Food Prot. 2018, 81, 1768–1775. [Google Scholar] [CrossRef]
- de Souza Duarte, A.F.; Ceotto-Vigoder, H.; Barrias, E.S.; Souto-Padrón, T.C.B.S.; Nes, I.F.; de Freire Bastos, M.D.C. Hyicin 4244, the first sactibiotic described in staphylococci, exhibits an anti-staphylococcal biofilm activity. Int. J. Antimicrob. Agents 2018, 51, 349–356. [Google Scholar] [CrossRef]
- Kommineni, S.; Bretl, D.J.; Lam, V.; Chakraborty, R.; Hayward, M.; Simpson, P.; Cao, Y.; Bousounis, P.; Kristich, C.J.; Salzman, N.H. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature 2015, 526, 719–722. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Goncheva, M.I.; Flannagan, R.S.; Deecker, S.R.; Guariglia-Oropeza, V.; Ensminger, A.W.; Heinrichs, D.E. Coagulase-negative staphylococci release a purine analog that inhibits Staphylococcus aureus virulence. Nat. Commun. 2021, 12, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Leistikow, K.R.; May, D.S.; Suh, W.S.; Asensio, G.V.; Schaenzer, A.J.; Currie, C.R.; Hristova, K.R. Bacillus subtilis-derived peptides disrupt quorum sensing and biofilm assembly in multidrug-resistant Staphylococcus aureus. mSystems 2024, 9, e00712–24. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, G.; Mercuri, S.; Martella, A.; Ferrara, F.; Simonetti, V.; Trifirò, C.; Emanuele, E. Topical application of bacteriocins from Bacillus subtilis promotes Staphylococcus aureus decolonization in acneic skin and improves the clinical appearance of mild-to-moderate acne. Adv. Dermatol. Allergol./Postępy Dermatol. I Alergol. 2023, 40, 115–118. [Google Scholar] [CrossRef]
- Chao, L.; Levin, B.R. Structured habitats and the evolution of anticompetitor toxins in bacteria. Proc. Natl. Acad. Sci. USA 1981, 78, 6324–6328. [Google Scholar] [CrossRef]
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, Ecology, and Application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Alajlani, M.M. Characterization of subtilosin gene in wild type Bacillus spp. and possible physiological role. Sci. Rep. 2022, 12, 10521. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsakhawy, O.K.; Abouelkhair, M.A. Genome Mining Reveals a Sactipeptide Biosynthetic Cluster in Staphylococcus pseudintermedius. Vet. Sci. 2025, 12, 635. https://doi.org/10.3390/vetsci12070635
Elsakhawy OK, Abouelkhair MA. Genome Mining Reveals a Sactipeptide Biosynthetic Cluster in Staphylococcus pseudintermedius. Veterinary Sciences. 2025; 12(7):635. https://doi.org/10.3390/vetsci12070635
Chicago/Turabian StyleElsakhawy, Ola K., and Mohamed A. Abouelkhair. 2025. "Genome Mining Reveals a Sactipeptide Biosynthetic Cluster in Staphylococcus pseudintermedius" Veterinary Sciences 12, no. 7: 635. https://doi.org/10.3390/vetsci12070635
APA StyleElsakhawy, O. K., & Abouelkhair, M. A. (2025). Genome Mining Reveals a Sactipeptide Biosynthetic Cluster in Staphylococcus pseudintermedius. Veterinary Sciences, 12(7), 635. https://doi.org/10.3390/vetsci12070635