
Gastrointestinal Parasites of Domestic Mammalian Hosts in Southeastern Iran

 ,
,  , , , ,
, , , ,  ,
,  and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
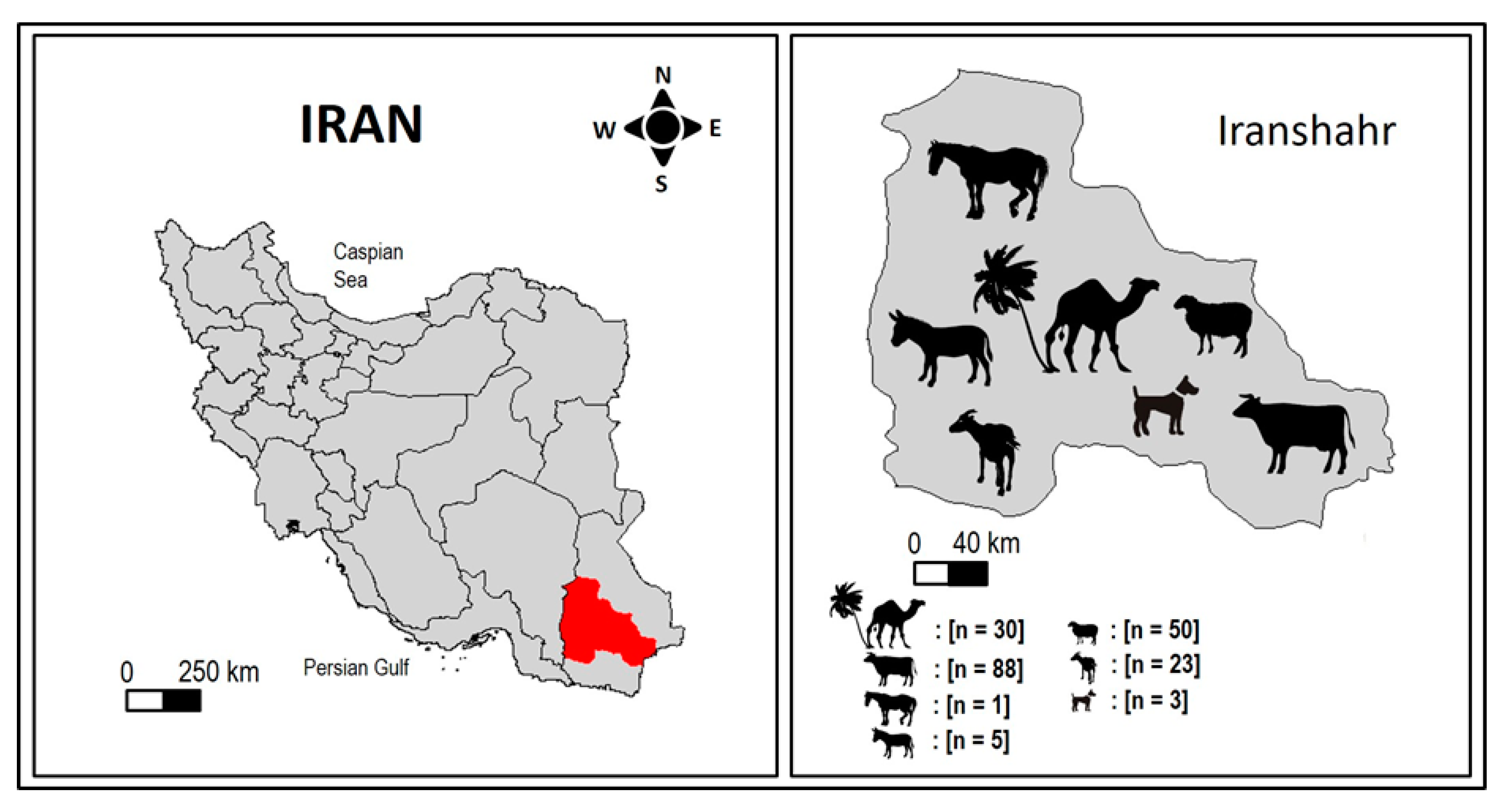
2.1. Study Area
2.2. Sample Size Estimation
2.3. Sample Collection
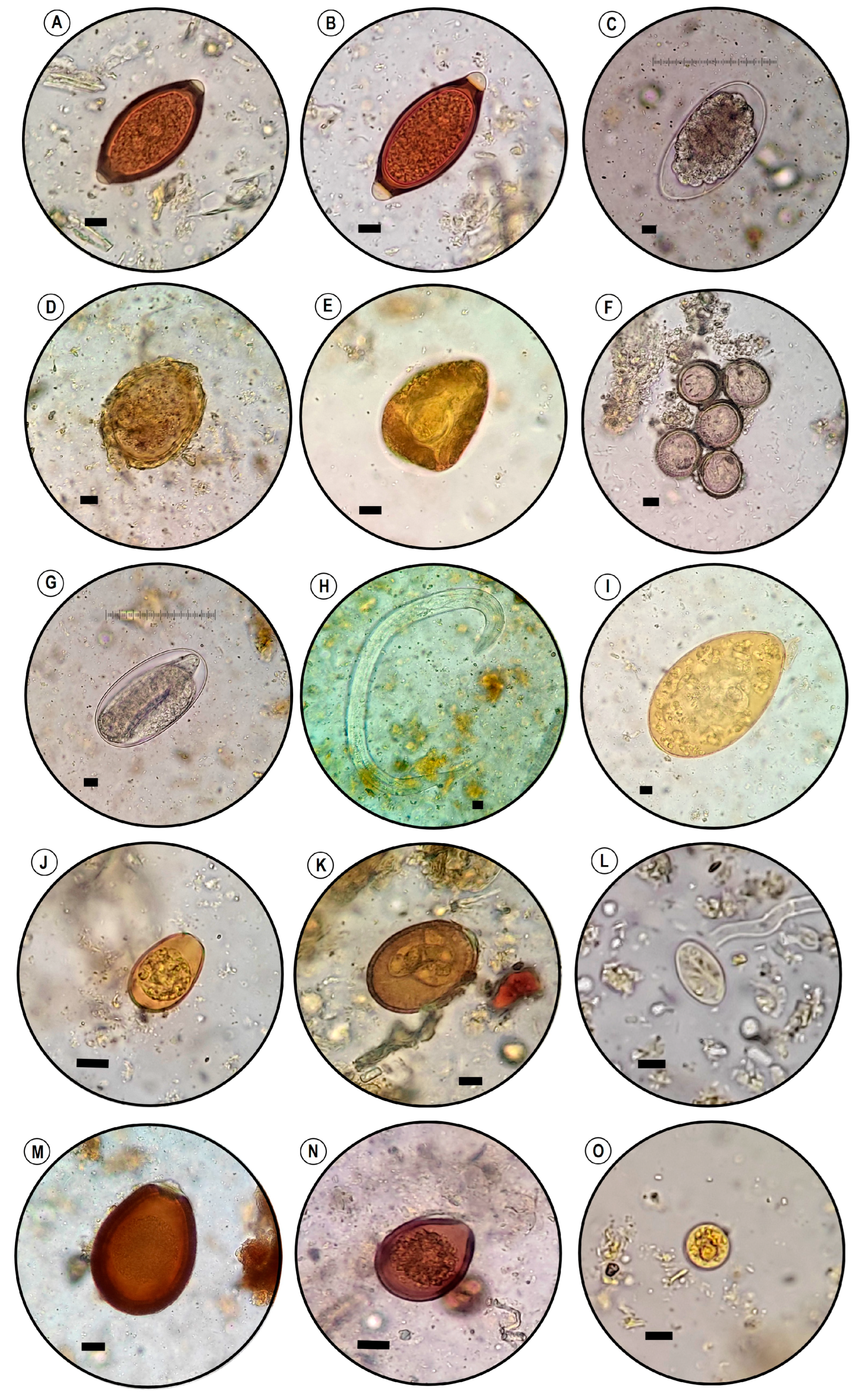
2.4. Coprological Analysis
2.5. Statistical Analysis
3. Results
3.1. Parasitological Findings
3.2. Risk Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raza, M.A.; Iqbal, Z.; Jabbar, A.; Yaseen, M. Point prevalence of gastrointestinal helminthiasis in ruminants in southern Punjab, Pakistan. J. Helminthol. 2007, 81, 323–328. [Google Scholar] [CrossRef]
- Lim, Y.A.; Mahdy, M.A.; Tan, T.K.; Goh, X.T.; Jex, A.R.; Nolan, M.J.; Sharma, R.S.; Gasser, R.B. First molecular characterization of Giardia duodenalis from goats in Malaysia. Mol. Cell. Probes 2013, 27, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Sultan, K.; Elmonir, W.; Hegazy, Y. Gastrointestinal parasites of sheep in Kafrelsheikh governorate, Egypt: Prevalence, control and public health implications. Beni-Suef. Univ. J. Basic Appl. Sci. 2016, 5, 79–84. [Google Scholar] [CrossRef][Green Version]
- Tavassoli, M.; Arjmand Yamchi, J.; Hajipour, N. A survey on the prevalence of strongyles species in working donkeys in North-West of Iran. J. Parasit. Dis. 2016, 40, 1210–1212. [Google Scholar] [CrossRef]
- Sazmand, A.; Bahari, A.; Papi, S.; Otranto, D. Parasitic diseases of equids in Iran (1931–2020): A literature review. Parasit. Vectors 2020, 13, 586. [Google Scholar] [CrossRef]
- Asghari, A.; Mahdavi, F.; Shamsi, L.; Motazedian, M.H.; Asgari, Q.; Shahabi, S.; Mohammadi-Ghalehbin, B.; Alireza Sadrebazzaz, A. Prevalence and molecular characterization of Giardia duodenalis in small ruminants of Shiraz, southwestern Iran: A zoonotic concern. Comp. Immunol. Microbiol. Infect. Dis. 2022, 86, 101819. [Google Scholar] [CrossRef] [PubMed]
- Vredenberg, I.; van Schaik, G.; van der Poel, W.H.M.; Stegeman, A. Coverage and representativeness of passive surveillance components for cattle and swine in The Netherlands. Animals 2022, 12, 3344. [Google Scholar] [CrossRef]
- Hewavithana, D.K.; Wijesinghe, M.R.; Udagama, P.V. Gastrointestinal parasites of six large mammals in the Wasgomuwa National Park, Sri Lanka. Int. J. Parasitol. Parasites Wildl. 2022, 17, 1–6. [Google Scholar] [CrossRef]
- Hatam-Nahavandi, K.; Fallah, E.; Asgharzadeh, M.; Mirsamadi, N.; Mahdavipour, B. Glutamate dehydrogenase and triosephosphate-isomerase coding genes for detection and genetic characterization of Giardia lamblia in human feces by PCR and PCR-RFLP. Turk. J. Med. Sci. 2011, 41, 283–289. [Google Scholar]
- Al-Habsi, K.; Yang, R.; Ryan, U.; Jacobson, C.; Miller, D.W. Morphological and molecular characterization of an uninucleated cyst-producing Entamoeba spp. in captured Rangeland goats in Western Australia. Vet. Parasitol. 2017, 235, 41–46. [Google Scholar] [CrossRef]
- Nolan, M.J.; Unger, M.; Yeap, Y.T.; Rogers, E.; Millet, I.; Harman, K.; Fox, M.; Kalema-Zikusoka, G.; Blake, D.P. Molecular characterisation of protist parasites in human-habituated mountain gorillas (Gorilla beringei beringei), humans and livestock, from Bwindi impenetrable National Park, Uganda. Parasit. Vectors 2017, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Ai, S.; Zhang, Z.; Wang, X.; Zhang, Q.; Yin, W.; Duan, Z. The first survey and molecular identification of Entamoeba spp. in farm animals on Qinghai-Tibetan Plateau of China. Comp. Immunol. Microbiol. Infect. Dis. 2021, 75, 101607. [Google Scholar] [CrossRef] [PubMed]
- Fallah, E.; Hatam-Nahavandi, K.; Jamali, R.; Mahdavi, B.; Asgharzadeh, M. Molecular identification of Giardia duodenalis isolates from human and animal reservoirs by PCR-RFLP. J. Biol. Sci. 2008, 8, 896–901. [Google Scholar] [CrossRef][Green Version]
- Fallah, E.; Mahdavi Poor, B.; Jamali, R.; Hatam-Nahavandi, K.; Asgharzadeh, M. Molecular characterization of Cryptosporidium isolates from cattle in a slaughterhouse in Tabriz, northwestern Iran. J. Biol. Sci. 2008, 8, 639–643. [Google Scholar] [CrossRef]
- Hatam-Nahavandi, K.; Mohebali, M.; Mahvi, A.H.; Keshavarz, H.; Najafian, H.R.; Mirjalali, H.; Rezaei, S.; Rezaeian, M. Microscopic and molecular detection of Cryptosporidium andersoni and Cryptosporidium xiaoi in wastewater samples of Tehran province, Iran. Iran. J. Parasitol. 2016, 11, 499–506. [Google Scholar]
- Eslami, A.; Ranjbar-Bahadori, S.; Meshgi, B.; Dehghan, M.; Bokaie, S. Helminth infections of stray dogs from Garmsar, Semnan province, central Iran. Iran. J. Parasitol. 2010, 5, 37–41. [Google Scholar]
- Carmena, D.; Cardona, G.A. Canine echinococcosis: Global epidemiology and genotypic diversity. Acta Trop. 2013, 128, 441–460. [Google Scholar] [CrossRef]
- Smith, L.A.; Marion, G.; Swain, D.L.; White, P.C.L.; Hutchings, M.R. The effect of grazing management on livestock exposure to parasites via the faecal-oral route. Prev. Vet. Med. 2009, 91, 95–106. [Google Scholar] [CrossRef]
- Lwanga, S.K.; Lemeshow, S. Sample Size Determination in Health Studies: A Practical Manual; World Health Organization: Geneva, Switzerland, 1991. [Google Scholar]
- Anvari-Tafti, M.; Sazmand, A.; Hekmatimoghaddam, S.; Moobedi, I. Gastrointestinal helminths of camels (Camelus dromedarius) in center of Iran. Trop. Biomed. 2013, 30, 56–61. [Google Scholar]
- Singh, E.; Kaur, P.; Singla, L.D.; Bal, M.S. Prevalence of gastrointestinal parasitism in small ruminants in western zone of Punjab, India. Vet. World 2017, 10, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Hatam-Nahavandi, K.; Mohebali, M.; Mahvi, A.H.; Keshavarz, H.; Khanaliha, K.; Tarighi, F.; Molaei Rad, M.B.; Rezaeian, T.; Charehdar, S.; Salimi, M.; et al. Evaluation of Cryptosporidium oocyst and Giardia cyst removal efficiency from urban and slaughterhouse wastewater treatment plants and assessment of cyst viability in wastewater effluent samples from Tehran, Iran. J. Water Reuse Desalination 2015, 5, 372–390. [Google Scholar] [CrossRef]
- Nahavandi, K.H.; Mahvi, A.H.; Mohebali, M.; Keshavarz, H.; Rezaei, S.; Mirjalali, H.; Elikaei, S.; Rezaeian, M. Molecular typing of Eimeria ahsata and Eimeria crandallis isolated from slaughterhouse wastewater. Jundishapur J. Microbiol. 2016, 9, e34140. [Google Scholar]
- Soulsby, E.J.L. Helminths, Arthropods and Protozoa of Domesticated Animals, 7th ed.; Bailliere Tindall: London, UK, 1982. [Google Scholar]
- Pilarczyk, B.; Tomza-Marciniak, A.; Pilarczyk, R.; Bombik, E.; Seremak, B.; Udała, J.; Sadowska, N. A Comparison of the prevalence of the parasites of the digestive tract in goats from organic and conventional farms. Animals 2021, 11, 2581. [Google Scholar] [CrossRef]
- Mhoma, J.R.L.; Kanyari, P.W.N.; Kagira, J.M. The prevalence of gastrointestinal parasites in goats in urban and peri-urban areas of Mwanza City, Tanzania. Sci. Parasitol. 2011, 12, 191–196. [Google Scholar]
- Tavassoli, M.; Khoshvaghti, H. Helminthes and coccidia infection of wild sheep (Ovis ammon orientalis) in Kabodan Island of National Park of Urmia Lake. Iran. Vet. Res. Forum 2010, 1, 26–29. [Google Scholar]
- Sazmand, A.; Hamidnejat, H.; Hekmatimoghaddam, S.; Asadollahi, Z.; Mirabdollahi, S. Eimeria infection in camels (Camelus dromedarius) in Yazd province, central Iran. Trop. Biomed. 2012, 29, 77–80. [Google Scholar] [PubMed]
- Borji, H.; Razmi, G.; Movassaghi, A.M. Prevalence of Cryptosporidium and Eimeria infections in dromedary (Camelus dromedarius) in abattoir of Mashhad, Iran. J. Camel Pract. Res. 2009, 16, 167–170. [Google Scholar]
- Yakhchali, M.; Cheraghi, E. Eimeriosis in Bactrian and dromedary camels in the Miandoab region, Iran. Acta Vet. 2007, 57, 545–552. [Google Scholar]
- Adinehbeigi, K.; Khedri, J.; Rahmani, K.; Afshari Moghaddam, A.; Hashemi, H. Determination and diversity of bovine coccidia in Zabol, east of Iran. Arch. Razi Inst. 2018, 73, 45–51. [Google Scholar]
- Kiani-Salmi, N.; Fattahi-Bafghi, A.; Astani, A.; Sazmand, A.; Zahedi, A.; Firoozi, Z.; Ebrahimi, B.; Dehghani-Tafti, A.; Ryan, U.; Akrami-Mohajeri, F. Molecular typing of Giardia duodenalis in cattle, sheep and goats in an arid area of central Iran. Infect. Genet. Evol. 2019, 75, 104021. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X.; Wang, R.; Liu, A.; Shen, Y.; Ling, H.; Cao, J.; Yang, F.; Zhang, X.; Zhang, L. Genetic characterizations of Giardia duodenalis in sheep and goats in Heilongjiang Province, China and possibility of zoonotic transmission. PLoS Negl. Trop. Dis. 2012, 6, e1826. [Google Scholar] [CrossRef]
- Peng, X.Q.; Tian, G.R.; Ren, G.J.; Yu, Z.Q.; Lok, J.B.; Zhang, L.X.; Wang, X.T.; Song, J.K.; Zhao, G.H. Infection rate of Giardia duodenalis, Cryptosporidium spp. and Enterocytozoon bieneusi in cashmere, dairy and meat goats in China. Infect. Genet. Evol. 2016, 41, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Gardner, G.E.; Ryan, U.; Jacobson, C. Prevalence and pathogen load of Cryptosporidium and Giardia in sheep faeces collected from saleyards and in abattoir effluent in Western Australia. Small Rumin. Res. 2015, 130, 216–220. [Google Scholar] [CrossRef]
- Santin, M.; Trout, J.M.; Fayer, R. Prevalence and molecular characterization of Cryptosporidium and Giardia species and genotypes in sheep in Maryland. Vet. Parasitol. 2007, 146, 17–24. [Google Scholar] [CrossRef]
- Jian, Y.; Zhang, X.; Li, X.; Karanis, G.; Ma, L.; Karanis, P. Prevalence and molecular characterization of Giardia duodenalis in cattle and sheep from the Qinghai-Tibetan Plateau Area (QTPA), northwestern China. Vet. Parasitol. 2018, 250, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Jafari, H.; Razi Jalali, M.H.; Shapouri, M.S.A.; Haji Hajikolaii, M.R. Determination of Giardia duodenalis genotypes in sheep and goat from Iran. J. Parasit. Dis. 2014, 38, 81–84. [Google Scholar] [CrossRef]
- Ozmen, O.; Yukari, B.A.; Haligur, M.; Sahinduran, S. Observations and immunohistochemical detection of Coronavirus, Cryptosporidium parvum and Giardia intestinalis in neonatal diarrhoea in lambs and kids. Schweiz. Arch. Tierheilkd. 2006, 148, 357–364. [Google Scholar] [CrossRef]
- Utaaker, K.S.; Myhr, N.; Singh Bajwa, R.; Joshi, H.; Kumar, A.; Robertson, L.J. Goats in the city: Prevalence of Giardia duodenalis and Cryptosporidium spp. in extensively reared goats in northern India. Acta Vet. Scand. 2017, 59, 86. [Google Scholar] [CrossRef]
- Minetti, C.; Taweenan, W.; Hogg, R.; Featherstone, C.; Randle, N.; Latham, S.M.; Wastling, J.M. Occurrence and diversity of Giardia duodenalis assemblages in livestock in the UK. Transbound. Emerg. Dis. 2014, 61, e60–e67. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Field, D.; Ryan, U. Molecular typing of Giardia duodenalis in humans in Queensland—First report of Assemblage E. Parasitology 2017, 144, 1154–1161. [Google Scholar] [CrossRef]
- Nouri, N.V.; Rahmatian, R.; Salehi, A. Prevalence of helminthic infections in the gastrointestinal tract of cattle in Mazandaran Province (Northern Iran). J. Parasitol. Res. 2022, 2022, 7424647. [Google Scholar] [CrossRef]
- Rahimi Esboei, B.; Mobedi, I.; Mizani, A.; Zare, R.; Vazini, H. A Seasonal survey on the helminths infections of the ruminants slaughtered in the abattoirs of Mazandaran province, northern Iran. J. Hum. Environ. Health Promot. 2020, 6, 142–146. [Google Scholar] [CrossRef]
- Sultan, K.; Desoukey, A.; Elsiefy, M.; Elbahy, N. An abattoir study on the prevalence of some gastrointestinal helminths of sheep in Gharbia Governorate, Egypt. Glob. Vet. 2010, 5, 84–87. [Google Scholar]
- Ashrafi, K.; Sharifdini, M.; Heidari, Z.; Rahmati, B.; Kia, E.B. Zoonotic transmission of Teladorsagia circumcincta and Trichostrongylus species in Guilan province, northern Iran: Molecular and morphological characterizations. BMC Infect. Dis. 2020, 20, 28. [Google Scholar] [CrossRef]
- Waruiru, R.; Kyvsgaard, N.; Thamsborg, S.; Nansen, P.; Bøgh, H.; Munyua, W.; Gathuma, J. The prevalence and intensity of helminth and coccidial infections in dairy cattle in central kenya. Vet. Res. Commun. 2000, 24, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Zvinorova, P.I.; Halimani, T.E.; Muchadeyi, F.C.; Matika, O.; Riggio, V.; Dzama, K. Prevalence and risk factors of gastrointestinal parasitic infections in goats in low-input low-output farming systems in Zimbabwe. Small Rumin. Res. 2016, 143, 75–83. [Google Scholar] [CrossRef]
- Lello, J.; Boag, B.; Fenton, A.; Stevenson, I.R.; Hudson, P. Competition and Mutualism among the gut helminths of a mammalian host. Nature 2004, 428, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.H.; Meshgi, B.; Eslami, A.; Bokai, S.; Sobhani, M.; Ebrahimi Samani, R. Prevalence and biodiversity of helminth parasites in donkeys (Equus asinus) in Iran. Int. J. Vet. Res. 2009, 3, 95–99. [Google Scholar]
- Živković, S.; Pavlović, I.; Mijatović, B.; Trailović, I.; Trailović, D. Prevalence, intensity and risks involved in helminth infections in domestic mountain pony and Balkan donkey in Nature Park Stara Planina, Serbia. Iran. J. Parasitol. 2021, 16, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Fesseha, H.; Aliye, S.; Mathewos, M.; Nigusie, K. Prevalence and risk factors associated with donkey gastrointestinal parasites in Shashemane and Suburbs, Oromia Region, Ethiopia. Heliyon 2022, 8, e12244. [Google Scholar] [CrossRef]
- Radfar, M.H.; Ebrahimy Maimand, A.; Sharify, A. A report on parasitic infections in camel (Camelus dromedarius) of Kerman slaughter house. J. Vet. Res. 2006, 61, 165–168. [Google Scholar]
- Borji, H.; Razmi, G.R.; Movassaghi, A.R.; Naghibi, A.; Maleki, M. A study on gastrointestinal helminths of camels in Mashhad Abattoir, Iran. Iran. J. Vet. Res. 2010, 11, 174–179. [Google Scholar]
- Omer, M.M.; Ahmed, A.M.; Abusalab, A. A retrospective study on animal parasitic diseases diagnosis at Kassala veterinary research lab (KVRL), Eastern Sudan. Vet. Res. 2007, 1, 68–70. [Google Scholar]
- Farooq, Z.; Mushtaq, S.; Iqbal, Z.; Akhtar, S. Parasitic helminths of domesticated and wild ruminants in Cholistan desert of Pakistan. Int. J. Agric. Biol. 2012, 14, 63–66. [Google Scholar]
- Ozdal, N.; Gul, A.; Ilhan, F.; Deger, S. Prevalence of Paramphistomum infection in cattle and sheep in Van Province, Turkey. Helminthologia 2010, 47, 20–24. [Google Scholar] [CrossRef]
- Titi, A.; Mekroud, A.; Sedraoui, S.; Vignoles, P.; Rondelaud, D. Prevalence and intensity of Paramphistomum daubneyi infections in cattle from north-eastern Algeria. J. Helminthol. 2010, 84, 177–181. [Google Scholar] [CrossRef] [PubMed]
- González-Warleta, M.; Lladosa, S.; Castro-Hermida, J.A.; Martínez-Ibeas, A.M.; Conesa, D.; Muñoz, F.; Mezo, M. Bovine paramphistomosis in Galicia (Spain): Prevalence, intensity, aetiology and geospatial distribution of the infection. Vet. Parasitol. 2013, 191, 252–263. [Google Scholar] [CrossRef]
- Nikpay, A.; Houshmand, E.; Eslami, A.; Bokaie, S. Epidemiology of cattle amphistomiasis in Gilan province, north of Iran. Comp. Clin. Pathol. 2019, 28, 83–87. [Google Scholar] [CrossRef]
- Eslami, A.; Halajian, A.; Bokaie, S. A survey on the bovine amphistomiasis in Mazanderan province, north of Iran. Iran. J. Vet. Res. 2011, 12, 52–55. [Google Scholar]
- Khedri, J.; Radfar, M.H.; Borji, H.; Mirzaei, M. Prevalence and intensity of Paramphistomum spp. in cattle from South-Eastern Iran. Iran. J. Parasitol. 2015, 10, 268–270. [Google Scholar]
- Hajipour, N.; Mirshekar, F.; Hajibemani, A.; Ghorani, M. Prevalence and risk factors associated with amphistome parasites in cattle in Iran. Vet. Med. Sci. 2021, 7, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Mansourian, M.; Moazeni, M.; Nikahval, B.; Barband, S. Liver distomatosis in cattle, sheep and goats of Northeastern Iran. Glob. Vet. 2011, 6, 241–246. [Google Scholar]
- Abdi, J.; Naserifar, R.; Nejad, M.R.; Mansouri, V. New features of fascioliasis in human and animal infections in Ilam province, Western Iran. Gastroenterol. Hepatol. Bed Bench 2013, 6, 152–155. [Google Scholar]
- Moghaddam, A.S.; Massoud, J.; Mahmoodi Mahvi, A.H.; Periago, M.V.; Artigas, P.; Fuentes, M.V.; Bargues, M.D.; Mas-Coma, S. Human and animal fascioliasis in Mazandaran province, northern Iran. Parasitol. Res. 2004, 94, 61–69. [Google Scholar] [CrossRef]
- Khoramian, H.; Arbabi, M.; Mahami Osqoi, M.; Delavari, M.; Hooshyar, H.; Asgari, M. Prevalence of ruminants’ fascioliasis and their economic effects in Kashan, center of Iran. Asian Pac. J. Trop. Biomed. 2014, 4, 918–922. [Google Scholar] [CrossRef]
- Khosravi, A.; Babaahmady, E. Epidemiology of Fasciola hepatica in Iran. Int. J. Biol. 2012, 4, 86–90. [Google Scholar] [CrossRef]
- Zhu, G.Q.; Li, L.; Ohiolei, J.A.; Wu, Y.T.; Li, W.H.; Zhang, N.Z.; Fu, B.Q.; Yan, H.B.; Jia, W.Z. A multiplex PCR assay for the simultaneous detection of Taenia hydatigena, T. multiceps, T. pisiformis, and Dipylidium caninum infections. BMC Infect. Dis. 2019, 19, 854. [Google Scholar] [CrossRef]
- Anene, B.M.; Onyekwodiri, E.O.; Chim, A.B.; Anika, S.M. Gastrointestinal parasites in sheep and goats of southeastern Nigeria. Small Rumin. Res. 1994, 13, 187–192. [Google Scholar] [CrossRef]
- Mirzaei, M.; Dahmardeh, E.; Sharifi, H. The prevalence of Eimeria species in sheep in Zabol city, Iran. Sci. Res. Iran. Vet. J. 2016, 11, 98–105. [Google Scholar]
- Etsay, K.; Megbey, S.; Yohannes, H. Prevalence of sheep and goat coccidiosis in different districts of Tigray region, Ethiopia. Niger. J. Anim. Sci. 2020, 22, 61–69. [Google Scholar]
- Carneiro, P.G.; Sasse, J.P.; Silva, A.C.S.; Seixas, M.; Paschoal, A.T.P.; Minutti, A.F.; Martins, T.A.; Cardim, S.T.; Rodrigues, F.S.; Barros, L.D.; et al. Prevalence and risk factors of Eimeria spp. natural infection in sheep from northern Paraná, Brazil. Braz. J. Vet. Parasitol. 2022, 31, e017421. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Ruiz, A.; Taubert, A. Eimeria bovis: An update on parasite-host cell interactions. Int. J. Med. Microbiol. 2012, 302, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Chartier, C.; Paraud, C. Coccidiosis due to Eimeria in sheep and goats, a review. Small Rumin. Res. 2012, 103, 84–92. [Google Scholar] [CrossRef]
- Waruiru, R.; Mutune, M.; Otieno, R. Gastrointestinal parasite infections of sheep and goats in semi-arid area of Machakos district, Kenya. Bull. Anim. Health Prod. Afr. 2005, 53, 25–34. [Google Scholar] [CrossRef]
- Dawit, I.; Weldegebriel, W.; Dejene, D.; Israel, I. Prevalence of gastrointestinal tract nematodes parasites in sheep in Hawasa town, Southern Ethiopia. Biomed. J. Sci. Tech. Res. 2022, 41, 33046–33052. [Google Scholar]
- Vlasoff, A.; Leathwick, D.M.; Heath, A.C.G. The epidemiology of nematode infections of sheep. N. Z. Vet. J. 2001, 49, 213–221. [Google Scholar] [CrossRef]
- Khan, M.N.; Sajid, M.S.; Khan, M.K.; Iqbal, Z.; Hussain, A. Gastrointestinal helminthiasis: Prevalence and associated determinants in domestic ruminants of district Toba Tek Singh, Punjab, Pakistan. Parasitol. Res. 2010, 107, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Jadidoleslami, A.; Siyadatpanah, A.; Borji, H.; Zarean, M.; Jarahi, L.; Moghaddas, E.; Budke, C.M. Prevalence and seasonality of adult and arrested larvae of gastrointestinal nematodes of sheep from Mashhad city, Northeastern Iran. Iran. J. Parasitol. 2022, 17, 214–222. [Google Scholar]



{kind=link}
{kind=link}
| Giardia duodenalis | Entamoeba spp. | Eimeria spp. | Helminths | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Host | Variable | Pos./Total | % | Pos./Total | % | Pos./Total | % | Pos./Total | % |
| Cattle | ≤1 yr. | 0/33 | 0.0 | 23/33 | 69.7 | 5/33 | 15.2 | 7/33 | 21.2 |
| >1 yr. | 0/55 | 0.0 | 41/55 | 74.5 | 7/55 | 12.7 | 10/55 | 18.2 | |
| Male | 0/21 | 0.0 | 13/21 | 61.9 | 7/21 | 33.3 | 5/21 | 23.8 | |
| Female | 0/67 | 0.0 | 51//67 | 76.1 | 5/67 | 7.5 | 12/67 | 17.9 | |
| Sub-total | 0/88 | 0.0 | 64/88 | 72.8 | 12/88 | 13.6 1 | 17/88 | 19.3 2 | |
| Sheep | ≤1 yr. | 1/24 | 4.2 | 1/24 | 4.2 | 17/24 | 70.8 | 6/24 | 25.0 |
| >1 yr. | 1/26 | 3.8 | 2/26 | 7.7 | 9/26 | 34.6 | 15/26 | 57.7 | |
| Male | 2/17 | 11.7 | 0/17 | 0.0 | 11/17 | 64.7 | 5/17 | 29.4 | |
| Female | 0/33 | 0.0 | 3/33 | 9.1 | 15/33 | 45.4 | 16/33 | 48.4 | |
| Sub-total | 2/50 | 4.0 | 3/50 | 6.0 | 26/50 | 52.0 3 | 21/50 | 42.0 4 | |
| Goat | ≤1 yr. | 1/12 | 8.3 | 1/12 | 8.3 | 9/12 | 75.0 | 2/12 | 16.6 |
| >1 yr. | 0/11 | 0.0 | 0/11 | 0.0 | 9/11 | 81.8 | 5/11 | 45.5 | |
| Male | 1/8 | 12.5 | 1/8 | 12.5 | 7/8 | 87.5 | 2/8 | 25.0 | |
| Female | 0/15 | 0.0 | 0/15 | 0.0 | 11/15 | 73.3 | 5/15 | 33.3 | |
| Sub-total | 1/23 | 4.3 | 1/23 | 4.3 | 18/23 | 78.2 5 | 7/23 | 30.4 6 | |
| Camel | ≤1 yr. | 0/5 | 0.0 | 0/5 | 0.0 | 2/5 | 40.0 | 1/5 | 20.0 |
| >1 yr. | 0/25 | 0.0 | 0/25 | 0.0 | 2/25 | 8.0 | 10/25 | 40.0 | |
| Male | 0/7 | 0.0 | 0/7 | 0.0 | 1/7 | 14.3 | 3/7 | 42.8 | |
| Female | 0/23 | 0.0 | 0/23 | 0.0 | 3/23 | 13.0 | 8/23 | 34.8 | |
| Sub-total | 0/30 | 0.0 | 0/30 | 0.0 | 4/30 | 13.3 7 | 11/30 | 36.6 8 | |
| Donkey | ≤1 yr. | 0/1 | 0.0 | 0/1 | 0.0 | 0/1 | 0.0 | 1/1 | 100 |
| >1 yr. | 0/4 | 0.0 | 0/4 | 0.0 | 0/4 | 0.0 | 3/4 | 75.0 | |
| Male | 0/1 | 0.0 | 0/1 | 0.0 | 0/1 | 0.0 | 1/1 | 100 | |
| Female | 0/4 | 0.0 | 0/4 | 0.0 | 0/4 | 0.0 | 3/4 | 75.0 | |
| Sub-total | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 4/5 | 80.0 9 | |
| Dog | ≤1 yr. | 0/1 | 0.0 | 0/1 | 0.0 | 0/1 | 0.0 | 1/1 | 100 |
| >1 yr. | 0/2 | 0.0 | 0/2 | 0.0 | 0/2 | 0.0 | 2/2 | 100 | |
| Male | 0/2 | 0.0 | 0/2 | 0.0 | 0/2 | 0.0 | 2/2 | 100 | |
| Female | 0/0 | 0.0 | 0/0 | 0.0 | 0/0 | 0.0 | 1/1 | 100 | |
| Sub-total | 0/3 | 0.0 | 0/3 | 0.0 | 0/3 | 0.0 | 3/3 | 100 10 | |
| Cattle (n = 88) | Sheep (n = 50) | Goat (n = 23) | Camel (n = 30) | Donkey (n = 5) | Dog (n = 3) | |
|---|---|---|---|---|---|---|
| Protozoa | ||||||
| Eimeria spp. | 13.6 (7.2–22.6) | 52.0 (37.4–66.3) | 78.2 (56.3–92.5) | 13.3 (3.7–30.7) | 0 (0.0) | 0 (0.0) |
| Entamoeba spp. | 72.8 (62.2–81.7) | 6.0 (1.2–16.5) | 4.3 (0.1–21.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Giardia duodenalis | 0 (0.0) | 4.0 (0.5–13.7) | 4.3 (0.1–21.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Helminths | ||||||
| Amphistomes | 2.3 (0.2–7.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Moniezia expansa | 0 (0.0) | 0 (0.0) | 0 (0.0) | 3.3 (0.1–17.2) | 0 (0.0) | 0 (0.0) |
| Strongyles spp. | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 80 (28.3–99.4) | 0 (0.0) |
| Taenia spp. | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 100 (29.2–100) |
| Trichostrongyles spp. | 15.9 (8.9–25.2) | 36 (22.9–50.8) | 17.3 (4.9–38.7) | 23.3 (9.9–42.2) | 0 (0.0) | 0 (0.0) |
| Trichuris spp. | 1.1 (0.1–6.1) | 6.0 (1.2–16.5) | 13 (2.7–33.6) | 10 (2.1–26.5) | 0 (0.0) | 0 (0.0) |
| Protozoa | Helminths | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Total (n) | Infected (n) | % | OR | 95% CI | Total (n) | Infected (n) | % | OR | 95% CI |
| Cattle (n = 88) | ||||||||||
| ≤1 yr. | 33 | 26 | 78.8 | 1.0 | Reference | 33 | 7 | 21.2 | 1.2 | 0.34–4.02 |
| >1 yr. | 55 | 46 | 83.6 | 1.4 | 0.38–4.69 | 55 | 10 | 18.2 | 1.0 | Reference |
| Male | 21 | 19 | 90.5 | 2.5 | 0.49–24.57 | 21 | 5 | 23.8 | 1.4 | 0.34–5.21 |
| Female | 67 | 53 | 79.1 | 1.0 | Reference | 67 | 12 | 17.9 | 1.0 | Reference |
| Sheep (n = 50) | ||||||||||
| ≤1 yr. | 24 | 18 | 75.0 | 3.5 | 1.05–11.66 | 24 | 6 | 25.0 | 1.0 | Reference |
| >1 yr. | 26 | 12 | 46.2 | 1.0 | Reference | 26 | 15 | 57.7 | 4.1 | 1.06–16.59 |
| Male | 17 | 13 | 76.5 | 3.1 | 0.72–15.29 | 17 | 5 | 29.4 | 1.0 | Reference |
| Female | 33 | 17 | 51.5 | 1.0 | Reference | 33 | 16 | 48.4 | 2.3 | 0.56–9.96 |
| Goat (n = 23) | ||||||||||
| ≤1 yr. | 12 | 9 | 75.0 | 1.0 | Reference | 12 | 2 | 8.3 | 1.0 | Reference |
| >1 yr. | 11 | 9 | 81.8 | 1.5 | 0.13–21.71 | 11 | 5 | 45.5 | 4.2 | 0.45–53.54 |
| Male | 8 | 7 | 87.5 | 2.5 | 0.23–27.71 | 8 | 2 | 12.5 | 1.0 | Reference |
| Female | 15 | 11 | 73.3 | 1.0 | Reference | 15 | 5 | 33.3 | 1.5 | 0.16–20.18 |
| Camel (n = 30) | ||||||||||
| ≤1 yr. | 5 | 2 | 40.0 | 7.7 | 0.77–76.45 | 5 | 1 | 20.0 | 1.0 | Reference |
| >1 yr. | 25 | 2 | 8.0 | 1.0 | Reference | 25 | 10 | 40.0 | 2.7 | 0.26–27.49 |
| Male | 7 | 1 | 14.3 | 1.1 | 0.02–17.15 | 7 | 3 | 42.8 | 1.4 | 0.16–10.64 |
| Female | 23 | 3 | 13.0 | 1.0 | Reference | 23 | 8 | 34.8 | 1.0 | Reference |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatam-Nahavandi, K.; Carmena, D.; Rezaeian, M.; Mirjalali, H.; Rahimi, H.M.; Badri, M.; Vafae Eslahi, A.; Shahrivar, F.F.; Rodrigues Oliveira, S.M.; Pereira, M.d.L.; et al. Gastrointestinal Parasites of Domestic Mammalian Hosts in Southeastern Iran. Vet. Sci. 2023, 10, 261. https://doi.org/10.3390/vetsci10040261
Hatam-Nahavandi K, Carmena D, Rezaeian M, Mirjalali H, Rahimi HM, Badri M, Vafae Eslahi A, Shahrivar FF, Rodrigues Oliveira SM, Pereira MdL, et al. Gastrointestinal Parasites of Domestic Mammalian Hosts in Southeastern Iran. Veterinary Sciences. 2023; 10(4):261. https://doi.org/10.3390/vetsci10040261
Chicago/Turabian StyleHatam-Nahavandi, Kareem, David Carmena, Mostafa Rezaeian, Hamed Mirjalali, Hanieh Mohammad Rahimi, Milad Badri, Aida Vafae Eslahi, Farzaneh Faraji Shahrivar, Sonia M. Rodrigues Oliveira, Maria de Lourdes Pereira, and et al. 2023. "Gastrointestinal Parasites of Domestic Mammalian Hosts in Southeastern Iran" Veterinary Sciences 10, no. 4: 261. https://doi.org/10.3390/vetsci10040261
APA StyleHatam-Nahavandi, K., Carmena, D., Rezaeian, M., Mirjalali, H., Rahimi, H. M., Badri, M., Vafae Eslahi, A., Shahrivar, F. F., Rodrigues Oliveira, S. M., Pereira, M. d. L., & Ahmadpour, E. (2023). Gastrointestinal Parasites of Domestic Mammalian Hosts in Southeastern Iran. Veterinary Sciences, 10(4), 261. https://doi.org/10.3390/vetsci10040261