
Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages


_Schenk.png)
Abstract
:1. Introduction
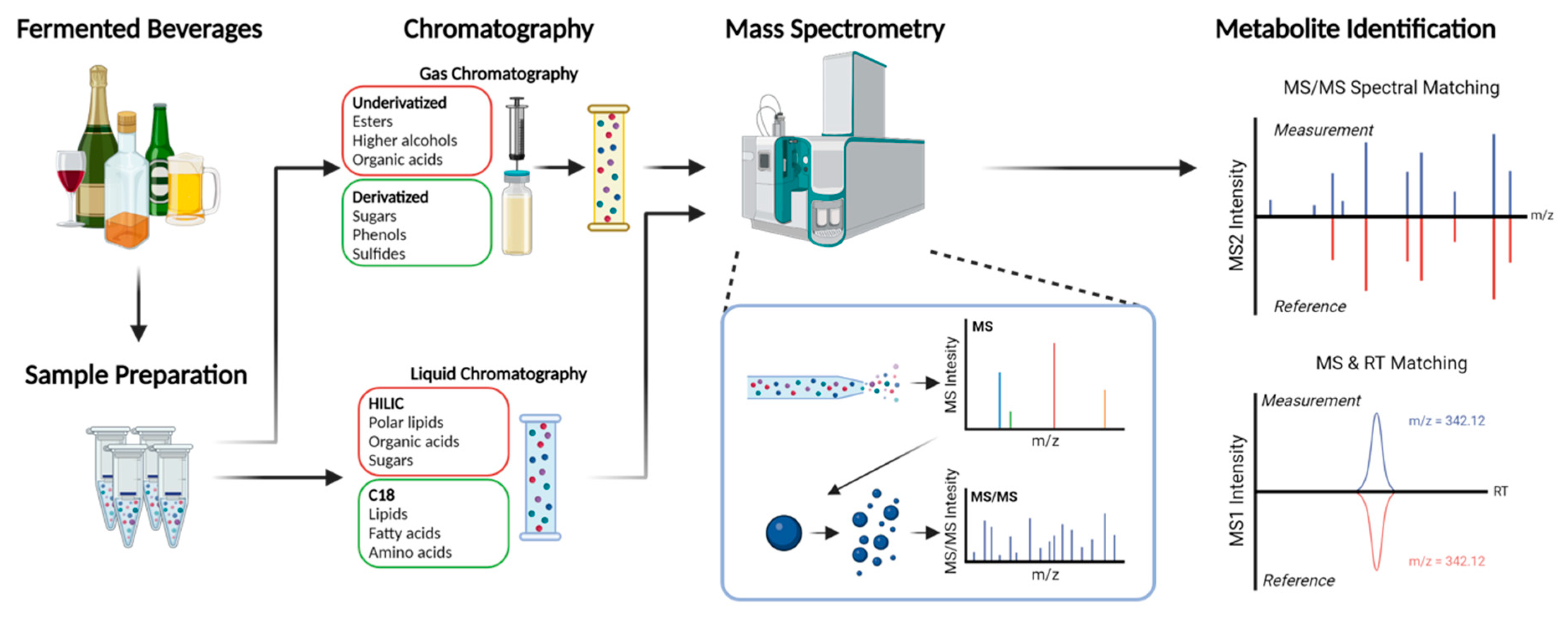
2. Metabolomics Techniques
3. Non-Saccharomyces Yeasts in Beer and Wine
3.1. Brettanomyces
3.2. Lachancea
3.3. Lambic and Mixed-Culture Beer
3.4. Torulaspora
3.5. Saccharomycodes ludwigii
3.6. Other Non-Saccharomyces Yeasts
4. Current Issues and Emerging Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.-S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erten, H.; Ağirman, B.; Gündüz, C.P.B.; Çarşanba, E.; Sert, S.; Bircan, S.; Tangüler, H. Importance of Yeasts and Lactic Acid Bacteria in Food Processing. In Food Processing: Strategies for Quality Assessment; Malik, A., Erginkaya, Z., Ahmad, S., Erten, H., Eds.; Springer: New York, NY, USA, 2014; pp. 351–378. [Google Scholar]
- Lahue, C.; Madden, A.A.; Dunn, R.R.; Smukowski Heil, C. History and Domestication of Saccharomyces cerevisiae in Bread Baking. Front. Genet. 2020, 11, 584718. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Verstrepen, K.J. Taming wild yeast: Potential of conventional and nonconventional yeasts in industrial fermentations. Annu. Rev. Microbiol. 2014, 68, 61–80. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef] [Green Version]
- Alsammar, H.F.; Naseeb, S.; Brancia, L.B.; Gilman, R.T.; Wang, P.; Delneri, D. Targeted metagenomics approach to capture the biodiversity of Saccharomyces genus in wild environments. Environ. Microbiol. Rep. 2019, 11, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Dequin, S.; Casaregola, S. The genomes of fermentative Saccharomyces. Comptes Rendus. Biol. 2011, 334, 687–693. [Google Scholar] [CrossRef]
- Maicas, S. The Role of Yeasts in Fermentation Processes. Microorganisms 2020, 8, 1142. [Google Scholar] [CrossRef]
- Coton, M.; Pawtowski, A.; Taminiau, B.; Burgaud, G.; Deniel, F.; Coulloumme-Labarthe, L.; Fall, A.; Daube, G.; Coton, E. Unraveling microbial ecology of industrial-scale Kombucha fermentations by metabarcoding and culture-based methods. FEMS Microbiol. Ecol. 2017, 93, fix048. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.P.; Taillandier, P. Understanding Kombucha Tea Fermentation: A Review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef]
- Tsutsumi, S.; Mochizuki, M.; Sakai, K.; Ieda, A.; Ohara, R.; Mitsui, S.; Ito, A.; Hirano, T.; Shimizu, M.; Kato, M. Ability of Saccharomyces cerevisiae MC87-46 to assimilate isomaltose and its effects on sake taste. Sci. Rep. 2019, 9, 13908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinkraus, K.H. Fermentations in World Food Processing. Compr. Rev. Food Sci. Food Saf. 2002, 1, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.K.; Ray, R.C. Wine Making: Basics and Applied Aspects, 1st ed.; Joshi, V.K., Ray, R.C., Eds.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Parker, D.K. 6—Beer: Production, sensory characteristics and sensory analysis. In Alcoholic Beverages; Piggott, J., Ed.; Woodhead Publishing: Sawston, UK, 2012; pp. 133–158. [Google Scholar]
- Briggs, D.E.; Brookes, P.A.; Stevens, R.; Boulton, C.A. Brewing: Science and Practice; Elsevier Science & Technology: Cambridge, UK, 2004. [Google Scholar]
- Walker, G.; Stewart, G. Saccharomyces cerevisiae in the Production of Fermented Beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Kerr, E.D.; McDiarmid, D.C.; Fraser, J.A.; Schulz, B.L. Cell Wall and Whole Cell Proteomes Define Flocculation and Fermentation Behavior of Yeast. Fermentation 2018, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Kerr, E.D.; Pegg, C.L.; Schulz, B.L. Proteomics and glycoproteomics of beer and wine. Proteomics 2022, e2100329. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Brewers Association. National Beer Sales & Production Data. New Brew. 2020. Available online: https://www.brewersassociation.org/statistics-and-data/national-beer-stats/ (accessed on 19 January 2022).
- Grand View Research. Kombucha Market Size, Share & Trends Analysis Report by Flavour (Original, Flavoured), By Distribution Channel (Supermarkets, Health Stores, Online Stores), By Region, And Segment Forecasts, 2020–2027. Available online: https://www.grandviewresearch.com/industry-analysis/kombucha-market (accessed on 1 February 2022).
- Brewers Association. Brewers Association Releases 2018 Econmimc Impact Report; Brewers Association: Boulder, CO, USA, 2018. [Google Scholar]
- Collins, S.L.; Koo, I.; Peters, J.M.; Smith, P.B.; Patterson, A.D. Current Challenges and Recent Developments in Mass Spectrometry-Based Metabolomics. Annu. Rev. Anal. Chem. 2021, 14, 467–487. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, N.; Johnson, D.L.; Herderich, M.J. Metabolomics approaches for resolving and harnessing chemical diversity in grapes, yeast and wine. Aust. J. Grape Wine Res. 2015, 21, 723–740. [Google Scholar] [CrossRef]
- Lei, Z.; Huhman, D.V.; Sumner, L.W. Mass spectrometry strategies in metabolomics. J. Biol. Chem. 2011, 286, 25435–25442. [Google Scholar] [CrossRef] [Green Version]
- Zeki, O.C.; Eylem, C.C.; Recber, T.; Kir, S.; Nemutlu, E. Integration of GC-MS and LC-MS for untargeted metabolomics profiling. J. Pharm. Biomed. Anal. 2020, 190, 113509. [Google Scholar] [CrossRef]
- Zhou, B.; Xiao, J.F.; Tuli, L.; Ressom, H.W. LC-MS-based metabolomics. Mol. Biosyst. 2012, 8, 470–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, M.; Vararu, F.; Moreno-Garcia, J.; Mauricio, J.C.; Moreno, J.; Garcia-Martinez, T. Analyzing the minor volatilome of Torulaspora delbrueckii in an alcoholic fermentation. Eur. Food Res. Technol. 2022, 248, 613–624. [Google Scholar] [CrossRef]
- Crauwels, S.; Van Opstaele, F.; Jaskula-Goiris, B.; Steensels, J.; Verreth, C.; Bosmans, L.; Paulussen, C.; Herrera-Malaver, B.; de Jonge, R.; De Clippeleer, J.; et al. Fermentation assays reveal differences in sugar and (off-) flavor metabolism across different Brettanomyces bruxellensis strains. FEMS Yeast Res. 2017, 17, fow105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.P.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Merlot wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to produce low-alcohol beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- De Roos, J.; Vandamme, P.; De Vuyst, L. Wort Substrate Consumption and Metabolite Production During Lambic Beer Fermentation and Maturation Explain the Successive Growth of Specific Bacterial and Yeast Species. Front. Microbiol. 2018, 9, 2763. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef]
- Stewart, G. The Production of Secondary Metabolites with Flavour Potential during Brewing and Distilling Wort Fermentations. Fermentation 2017, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Wedral, D.; Shewfelt, R.; Frank, J. The challenge of Brettanomyces in wine. LWT Food Sci. Technol. 2010, 43, 1474–1479. [Google Scholar] [CrossRef]
- Shimotsu, S.; Asano, S.; Iijima, K.; Suzuki, K.; Yamagishi, H.; Aizawa, M. Investigation of beer-spoilage ability of Dekkera/Brettanomyces yeasts and development of multiplex PCR method for beer-spoilage yeasts. J. Inst. Brew. 2015, 121, 177–180. [Google Scholar] [CrossRef]
- Branco, P.; Coutinho, R.; Malfeito-Ferreira, M.; Prista, C.; Albergaria, H. Wine Spoilage Control: Impact of Saccharomycin on Brettanomyces bruxellensis and Its Conjugated Effect with Sulfur Dioxide. Microorganisms 2021, 9, 2528. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Steensels, J.; Aerts, G.; Willems, K.; Verstrepen, K.; Lievens, B. Brettanomyces Bruxellensis, Essential Contributor in Spontaneous Beer Fermentations Providing Novel Opportunities for the Brewing Industry. Brew. Sci. 2015, 68, 110–121. [Google Scholar]
- Colomer, M.S.; Chailyan, A.; Fennessy, R.T.; Olsson, K.F.; Johnsen, L.; Solodovnikova, N.; Forster, J. Assessing Population Diversity of Brettanomyces Yeast Species and Identification of Strains for Brewing Applications. Front. Microbiol. 2020, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; House, J.F.; Joseph, C.M.L.; Bisson, L.F.; Bamforth, C.W. Lachancea thermotolerans as an alternative yeast for the production of beer†. J. Inst. Brew. 2016, 122, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Osburn, K.; Amaral, J.; Metcalf, S.R.; Nickens, D.M.; Rogers, C.M.; Sausen, C.; Caputo, R.; Miller, J.; Li, H.; Tennessen, J.M.; et al. Primary souring: A novel bacteria-free method for sour beer production. Food Microbiol. 2018, 70, 76–84. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Hutzler, M.; Zarnkow, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Investigation into the Potential of Lachancea fermentati Strain KBI 12.1 for Low Alcohol Beer Brewing. J. Am. Soc. Brew. Chem. 2019, 77, 157–169. [Google Scholar] [CrossRef]
- Bellut, K.; Krogerus, K.; Arendt, E.K. Lachancea fermentati Strains Isolated From Kombucha: Fundamental Insights, and Practical Application in Low Alcohol Beer Brewing. Front. Microbiol. 2020, 11, 764. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons, Incorporated: New York, NY, USA, 2016; p. 464. [Google Scholar]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef]
- Hranilovic, A.; Bely, M.; Masneuf-Pomarede, I.; Jiranek, V.; Albertin, W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE 2017, 12, e0184652. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Bamforth, C.W.; Mills, D.A. Brewhouse-resident microbiota are responsible for multi-stage fermentation of American coolship ale. PLoS ONE 2012, 7, e35507. [Google Scholar] [CrossRef]
- Verachtert, H.; Iserentant, D. Properties of Belgian acid beers and their microflora. The production of Gueuze and related refreshing acid beers. Cerevisia Belg. J. Brew. Biotechnol. 1995, 20, 37–41. [Google Scholar]
- Van Oevelen, D.; Spaepen, M.; Timmermans, P.; Verachtert, H. Microbiological aspects of spontaneous wort fermentation in the production of lambic and gueuze. J. Inst. Brew. 1977, 83, 356–360. [Google Scholar] [CrossRef]
- Reina, L.D.; Perez-Diaz, I.M.; Breidt, F.; Azcarate-Peril, M.A.; Medina, E.; Butz, N. Characterization of the microbial diversity in yacon spontaneous fermentation at 20 degrees C. Int. J. Food Microbiol. 2015, 203, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of an industrially produced lambic beer shares members of a traditionally produced one and reveals a core microbiota for lambic beer fermentation. Food Microbiol. 2015, 49, 23–32. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef]
- Martens, H.; Dawoud, E.; Verachtert, H. Wort Enterobacteria and Other Microbial Populations Involved During the First Month Of Lambic Fermentation. J. Inst. Brew. 1991, 97, 435–439. [Google Scholar] [CrossRef]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Tabanelli, G. Biogenic Amines and Food Quality: Emerging Challenges and Public Health Concerns. Foods 2020, 9, 859. [Google Scholar] [CrossRef]
- Loret, S.; Deloyer, P.; Dandrifosse, G. Levels of biogenic amines as a measure of the quality of the beer fermentation process: Data from Belgian samples. Food Chem. 2005, 89, 519–525. [Google Scholar] [CrossRef]
- Michel, M.; Kopecka, J.; Meier-Dornberg, T.; Zarnkow, M.; Jacob, F.; Hutzler, M. Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast 2016, 33, 129–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Tataridis, P.; Kanelis, A.; Logotetis, S.; Nerancis, E. Use of non-saccharomyces Torulaspora delbrueckii yeast strains in winemaking and brewing. Zb. Matice Srp. Za Prir. Nauk. 2013, 124, 415–426. [Google Scholar] [CrossRef]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Richard Dickinson, J. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Alves-Araujo, C.; Pacheco, A.; Almeida, M.J.; Spencer-Martins, I.; Leao, C.; Sousa, M.J. Sugar utilization patterns and respiro-fermentative metabolism in the baker’s yeast Torulaspora delbrueckii. Microbiology 2007, 153, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Azzolini, M.; Fedrizzi, B.; Tosi, E.; Finato, F.; Vagnoli, P.; Scrinzi, C.; Zapparoli, G. Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 2012, 235, 303–313. [Google Scholar] [CrossRef]
- Albertin, W.; Chasseriaud, L.; Comte, G.; Panfili, A.; Delcamp, A.; Salin, F.; Marullo, P.; Bely, M. Winemaking and bioprocesses strongly shaped the genetic diversity of the ubiquitous yeast Torulaspora delbrueckii. PLoS ONE 2014, 9, e94246. [Google Scholar] [CrossRef] [Green Version]
- Herraiz, T.; Reglero, G.; Herraiz, M.; Martin-Alvarez, P.J.; Cabezudo, M.D. The Influence of the Yeast and Type of Culture on the Volatile Composition of Wines Fermented Without Sulfur Dioxide. Am. J. Enol. Vitic. 1990, 41, 313–318. [Google Scholar]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Krogerus, K.; Eerikainen, R.; Aisala, H.; Gibson, B. Repurposing brewery contaminant yeast as production strains for low-alcohol beer fermentation. Yeast 2022, 39, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.A.; Andres-Iglesias, C.; Montero, O. Low-alcohol Beers: Flavor Compounds, Defects, and Improvement Strategies. Crit. Rev. Food Sci. Nutr. 2016, 56, 1379–1388. [Google Scholar] [CrossRef]
- Larroque, M.N.; Carrau, F.; Farina, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and non-Saccharomyces native yeasts on beer aroma compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef] [PubMed]
- Methner, Y.; Hutzler, M.; Zarnkow, M.; Prowald, A.; Endres, F.; Jacob, F. Investigation of Non-Saccharomyces Yeast Strains for Their Suitability for the Production of Non-Alcoholic Beers with Novel Flavor Profiles. J. Am. Soc. Brew. Chem. 2022, 30, 1–15. [Google Scholar] [CrossRef]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Rodríguez Madrera, R.; Pando Bedriñana, R.; Suárez Valles, B. Evaluation of indigenous non-Saccharomyces cider yeasts for use in brewing. Eur. Food Res. Technol. 2021, 247, 819–828. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Jacob, F.; De Schutter, D.; Daenen, L.; Lynch, K.; Zannini, E.; Arendt, E. Application of Non-Saccharomyces Yeasts Isolated from Kombucha in the Production of Alcohol-Free Beer. Fermentation 2018, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O.; Robertson, D.; Griffin, J.; van der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The metabolomics standards initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Johnson, S.R.; Lange, B.M. Open-access metabolomics databases for natural product research: Present capabilities and future potential. Front. Bioeng. Biotechnol. 2015, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Kind, T.; Scholz, M.; Fiehn, O. How large is the metabolome? A critical analysis of data exchange practices in chemistry. PLoS ONE 2009, 4, e5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alperstein, L.; Gardner, J.M.; Sundstrom, J.F.; Sumby, K.M.; Jiranek, V. Yeast bioprospecting versus synthetic biology-which is better for innovative beverage fermentation? Appl. Microbiol. Biotechnol. 2020, 104, 1939–1953. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for brewers: Exploiting natural diversity for naturally diverse beers. Yeast 2019, 36, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Qvirist, L.A.; De Filippo, C.; Strati, F.; Stefanini, I.; Sordo, M.; Andlid, T.; Felis, G.E.; Mattarelli, P.; Cavalieri, D. Isolation, Identification and Characterization of Yeasts from Fermented Goat Milk of the Yaghnob Valley in Tajikistan. Front. Microbiol. 2016, 7, 1690. [Google Scholar] [CrossRef] [Green Version]
- Buys, E.; Steenkamp, L.; Dlamini, N.R.; Moodley, S.S. Bacteria and yeast isolation and characterisation from a South African fermented beverage. S. Afr. J. Sci. 2019, 115, 1–6. [Google Scholar] [CrossRef]
- Sefa-Dedeh, S.; Sanni, A.I.; Tetteh, G.; Sakyi-Dawson, E. Yeasts in the traditional brewing of pito in Ghana. World J. Microbiol. Biotechnol. 1999, 15, 593–597. [Google Scholar] [CrossRef]
- Jespersen, L. Occurrence and taxonomic characteristics of strains of predominant in African indigenous fermented foods and beverages. FEMS Yeast Res. 2003, 3, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Pu, S.; Zhang, Y.; Lu, N.; Shi, C.; Yan, S. Yeasts from Chinese strong flavour Daqu samples: Isolation and evaluation of their potential for fortified Daqu production. AMB Express 2021, 11, 176. [Google Scholar] [CrossRef]
- Motlhanka, K.; Lebani, K.; Boekhout, T.; Zhou, N. Fermentative Microbes of Khadi, a Traditional Alcoholic Beverage of Botswana. Fermentation 2020, 6, 51. [Google Scholar] [CrossRef]
- Pilo, F.B.; Carvajal-Barriga, E.J.; Guaman-Burneo, M.C.; Portero-Barahona, P.; Dias, A.M.M.; Freitas, L.F.D.; Gomes, F.C.O.; Rosa, C.A. Saccharomyces cerevisiae populations and other yeasts associated with indigenous beers (chicha) of Ecuador. Braz. J. Microbiol. 2018, 49, 808–815. [Google Scholar] [CrossRef]
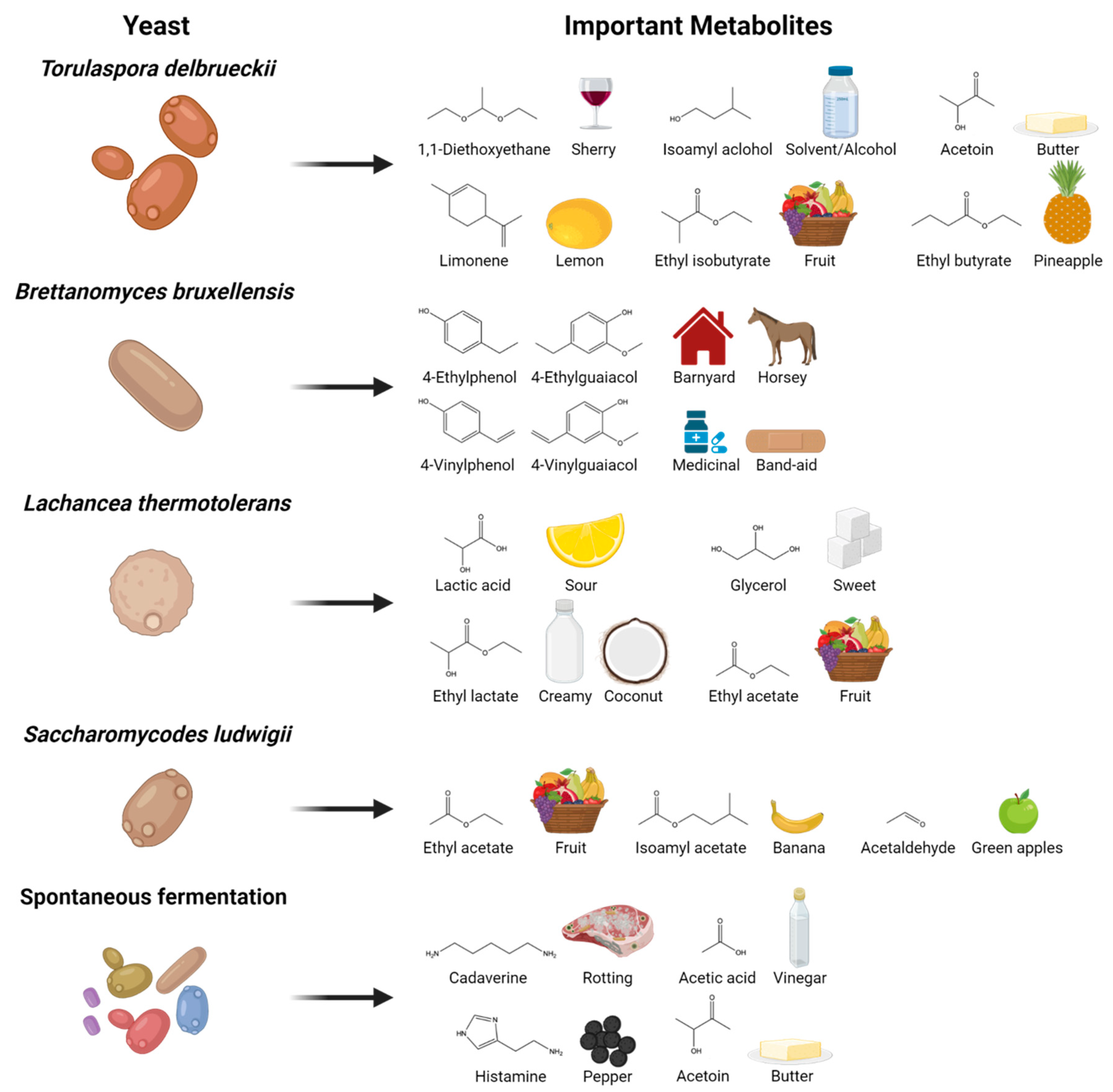

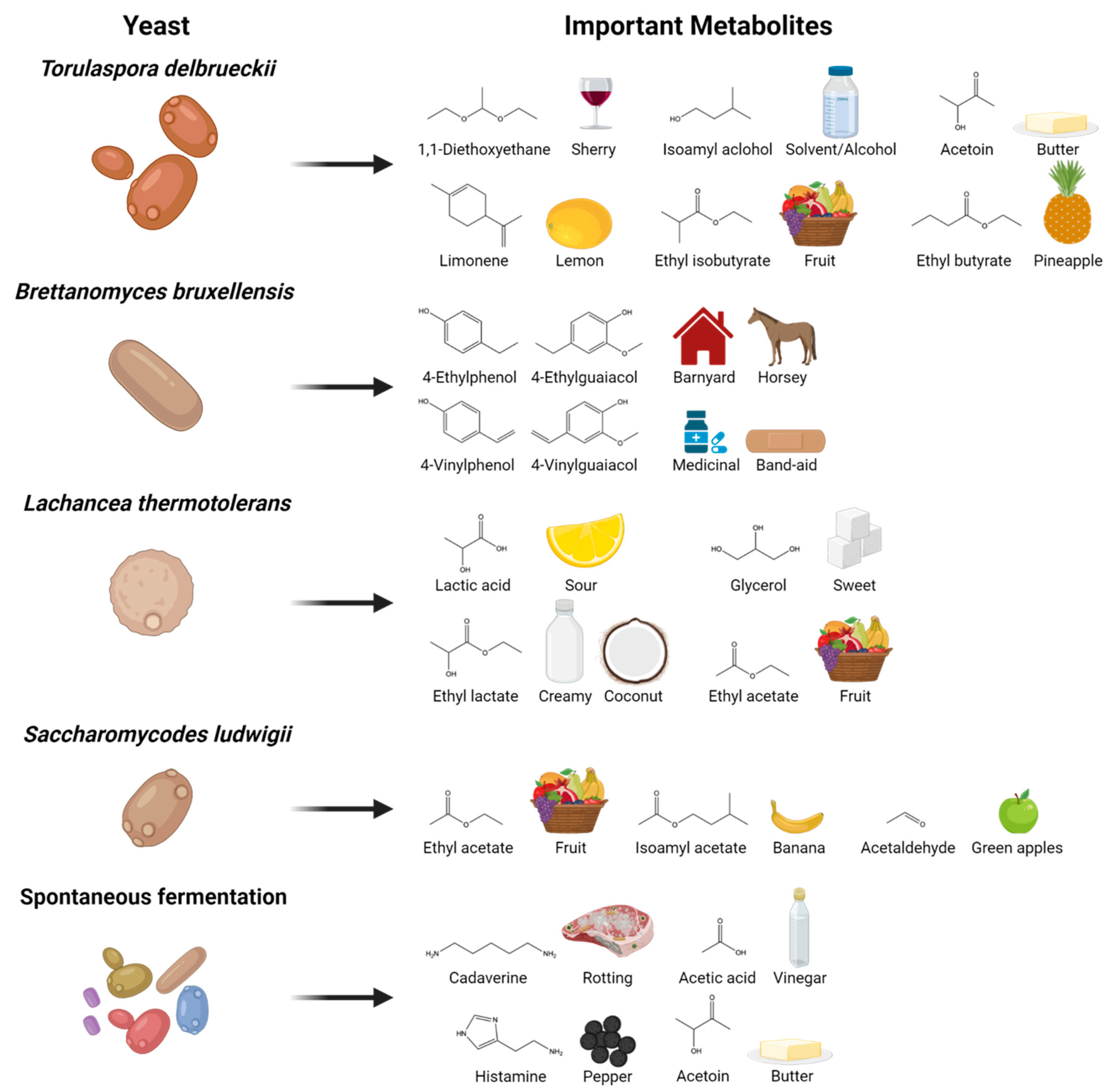{kind=link}
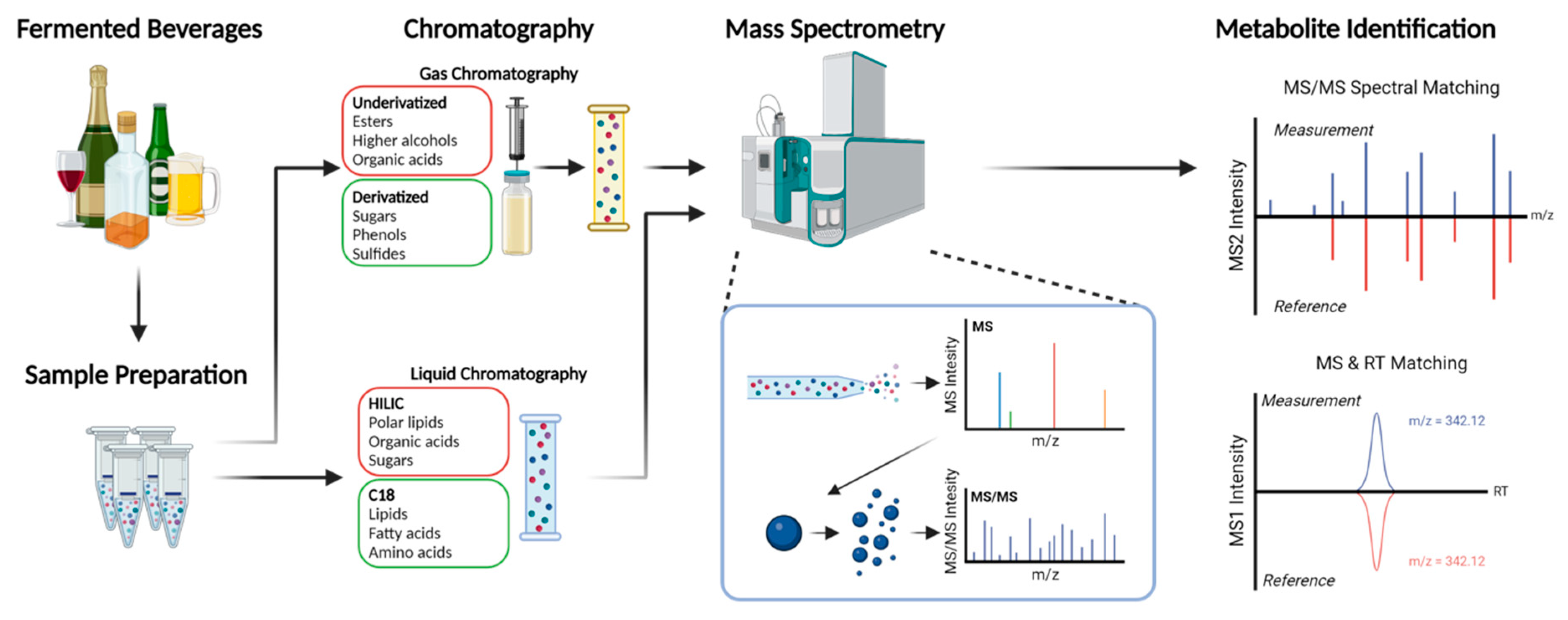{kind=link}
| Yeast | Metabolite of Interest | Analytical Method Used | Reference |
|---|---|---|---|
| Torulaspora delbreuckii | 1,1-Diethoxyethane | SBSE-TD-GCMS | [29] |
| Limonene | SBSE-TD-GCMS | ||
| Isoamyl alcohol | GC-FID | ||
| Acetoin | GC-FID | ||
| Ethyl isobutyrate | SBSE-TD-GCMS | ||
| Ethyl butyrate | SBSE-TD-GCMS | ||
| Brettanomyces bruxellensis | 4-Ethylphenol | SPME-GCMS | [30] |
| 4-Ethylguaiacol | SPME-GCMS | ||
| 4-Vinylphenol | SPME-GCMS | ||
| 4-Vinylguaiacol | SPME-GCMS | ||
| Lachancea thermotolerans | Lactic acid | HPLC-RI | [31] |
| Ethyl lactate | HS-SPME-GCMS | ||
| Ethyl acetate | HS-SPME-GCMS | ||
| Glycerol | HPLC-RI | ||
| Saccharomycodes ludwigii | Ethyl acetate | SPME-GCMS | [32] |
| Isoamyl acetate | SPME-GCMS | ||
| Acetaldehyde | SPME-GCMS | ||
| Spontaneous fermentation | Cadaverine | UPLC-MS/MS | [33] |
| Acetic acid | HPLC-RI | ||
| Histamine | UPLC-MS/MS | ||
| Acetoin | GC-FID |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellis, D.J.; Kerr, E.D.; Schenk, G.; Schulz, B.L. Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages. Beverages 2022, 8, 41. https://doi.org/10.3390/beverages8030041
Ellis DJ, Kerr ED, Schenk G, Schulz BL. Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages. Beverages. 2022; 8(3):41. https://doi.org/10.3390/beverages8030041
Chicago/Turabian StyleEllis, Daniel J., Edward D. Kerr, Gerhard Schenk, and Benjamin L. Schulz. 2022. "Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages" Beverages 8, no. 3: 41. https://doi.org/10.3390/beverages8030041
APA StyleEllis, D. J., Kerr, E. D., Schenk, G., & Schulz, B. L. (2022). Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages. Beverages, 8(3), 41. https://doi.org/10.3390/beverages8030041