Abstract
Probiotic features and the ability of two oenological lactic acid bacteria strains (Pediococcus pentosaceus CIAL-86 and Lactobacillus plantarum CIAL-121) and a reference probiotic strain (Lactobacillus plantarum CLC 17) to metabolize wine polyphenols are examined. After summarizing previous results regarding their resistance to lysozyme, gastric juice and bile salts, the three strains were assessed for their ability to release phenolic metabolites after their incubation with a wine phenolic extract. Neither of the two bacteria were able to metabolize wine polyphenols, at least in the conditions used in this study, although a certain stimulatory effect on bacterial growth was observed in the presence of a wine-derived phenolic metabolite (i.e., 3,4-dihydroxyphenylacetic acid) and a wine phenolic compound (i.e., (+)-catechin). Bacteria cell-free supernatants from the three strains delayed and inhibited almost completely the growth of the pathogen E. coli CIAL-153, probably due to the presence of organic acids derived from the bacterial metabolism of carbohydrates. Lastly, the three strains showed a high percentage of adhesion to intestinal cells, and pre-incubation of Caco-2 cells with bacteria strains prior to the addition of E. coli CIAL-153 produced a notable inhibition of the adhesion of E. coli to the intestinal cells.
Keywords:
wine; lactic acid bacteria; probiotics; polyphenols; cell adhesion; E. coli; phenolic metabolism 1. Introduction
Probiotics are live microorganisms that promote healthy gastrointestinal microbiota and boost the immune response [1]. The main probiotic strains belong to the genera Lactobacillus and Bifidobacterium and were mainly isolated from dairy products or the human gastrointestinal tract. Some studies have evaluated the probiotic potential of bacterial strains isolated from alcoholic fermented beverages such as cider [2] and wine [3]. Recently, the probiotic features of strains of lactic acid bacteria (LAB) from an oenological bacteria collection, including Lactobacillus spp., Pediococcus spp. and Oenococcus oeni, have been assessed, although their mode of action is still poorly understood [4]. On the other hand, consumption of probiotics, which are able to metabolize polyphenols into physiologically active metabolites, has been proposed as a nutritional approach to improve the bioavailability of these phytochemicals, which would, in turn, enhance the health effects attributed to them [5,6].
Beneficial health effects derived from the moderate consumption of wine and its bioactive compounds, especially polyphenols, have been evidenced mainly in relation to diseases associated with oxidative stress and inflammation [7,8]. Currently, the beneficial effects of wine polyphenols on intestinal microbiota growth and functionality is a topic that is attracting research [9,10]. Wine polyphenols include benzoic and cinnamic acids, phenolic alcohols and stilbenes among the non-flavonoids and anthocyanins, and flavan-3-ols, flavonols and others among the flavonoids. Most of them are minimally absorbed in the small intestine but they are extensively metabolized by enzymes from the colonic microbiota [11,12]. As colonic catabolites could be present in higher concentrations than the parent compounds, the biological activities attributed to polyphenols seem to be mainly due to them [13,14]. Therefore, the bioactivity of wine polyphenols is likely to be dependent on the microbiota activity that shows great human inter-individual differences [12].
On the other hand, numerous studies seem to indicate that phenolic compounds could positively modulate gut microbiota through prebiotic effects either promoting the growth of beneficial bacteria or having antimicrobial activity against pathogenic intestinal bacteria [15]. For instance, grape seed extracts of different flavan-3-ol composition have shown to promote the growth of potentially beneficial bacteria (Lactobacillus sp.) and decreased undesirable bacteria such as clostridia after batch culture fermentations [16]. Phenolic compounds contained in a cocoa powder reduced the growth of some members of the genera Staphylococcus and Clostridium, affecting the intestinal microbiota profile [17].
In this paper, we aimed to investigate more deeply the properties of potentially probiotic wine-isolated LAB. Thus, the objectives were: (a) to assess whether LAB were able to degrade wine polyphenols with the subsequent release of phenolic metabolites; (b) to monitor LAB growth in the presence and absence of some wine-related phenolic compounds; and (c) to evaluate LAB adherence to human intestinal cells, also considering the potential inhibition of the adherence of a pathogen E. coli strain.
2. Materials and Methods
2.1. Materials and Reagents
(+)-Catechin and 3,4-dihydroxyphenylacetic acid were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA). A commercial wine phenolic extract (Provinols™) was kindly provided by Safic-Alcan (SAU, Barcelona, Spain). Its total phenolic content was 474 mg of gallic acid per gram, and the main phenolic compounds present in the extract were: anthocyanins (delphinidin-3-O-glucoside, 0.568 mg/g; cyanidin-3-O-glucoside, 0.265 mg/g; petunidin-3-O-glucoside, 1.47 mg/g; peonidin-3-O-glucoside, 1.78 mg/g; malvidin-3-O-glucoside, 9.01 mg/g; malvidin-3-O-(6″-acetyl) glucoside, 1.92 mg/g; and malvidin-3-O-(6″-p-coumaroyl)glucoside, 1.24 mg/g), flavan-3-ols (catechin, 9.90 mg/g; epicatechin, 6.87 mg/g; epicatechin-3-O-gallate, 0.226 mg/g; procyanidin B1, 11.1 mg/g; and procyanidin B2, 4.69 mg/g), benzoic acids (gallic acid, 1.06 mg/g), cinnamates (cutaric acid, 2.00 mg/g; and caftaric acid, 0.192 mg/g), flavonols (quercetin, 22.4 mg/g; kaempherol, 2.71 mg/g; myricetin, 2.55 mg/g; quercetin-3-O-glucoside, 0.137 mg/g; and quercetin-3-O-galactoside, 0.107 mg/g), stilbenes (resveratrol, 0.427 mg/g; and resveratrol-3-O-glucoside, 9.17 mg/g) and others (tyrosol 18.9 mg/g) [18]. The wine extract was dissolved (0.6 mg/mL) in a saline solution (NaCl 0.9%) supplemented with 0.5 g/L of cysteine and sterilized by filtration.
Formic acid was purchased from Scharlau (Scharlau, Barcelona, Spain). Acetonitrile (HPLC-MS grade) was purchased from Labscan (POCH S.A., Gliwice, Poland). Ultrapure water was obtained using a Milli-Q system (Waters Millipore, Milford, MA, USA).
2.2. LAB Strains and E. coli Strain
Two LAB strains (P. pentosaceus CIAL-86 and L. plantarum CIAL-121), previously isolated from wines [19,20], were selected based on their good probiotic features [4]. Also, a previously characterized probiotic LAB strain isolated from breast milk (L. plantarum CLC 17) [21] was included as a reference probiotic strain. The strain E. coli CIAL-153 was isolated from human faeces [4].
All strains were kept frozen at −70 °C in a sterilized mixture of culture medium and glycerol (80:20, v/v). Inocula of the three LAB strains and E. coli CIAL-153 strain (108 CFU/mL) were prepared by growing overnight the contents of thawed cryovials in MRS broth (Pronadisa, Madrid, Spain) at 30 °C and TSB broth (Scharlau, Barcelona, Spain) at 37 °C, respectively. Bacteria were harvested by centrifugation (10,000× g, 10 min, 4 °C) and resuspended in Dulbecco’s phosphate-buffered saline DPBS (DPBS Lonza Walkersville, Inc., USA).
In vitro analyses were previously carried out to evaluate the resistance of LAB strains to conditions in the gastrointestinal tract including saliva and acid resistance, and bile tolerance [4], and the data obtained are now summarized in Table 1. All strains showed great resistance to lysozyme (>51%) and capacity to survive at low pH values (pH 1.8), thereby suggesting good adaptation of the wine LAB strains to the hostile gastrointestinal environment. Moreover, the growth percentages of both oenological LAB strains at the maximum concentration of bile assayed (1%) were higher than 84%, which was even greater than that exhibited by the reference probiotic strain, L. plantarum CLC 17 (73%), which reflected good bile resistance.

Table 1.
Resistance to lysozyme (% Survival), tolerance to simulated gastric juice on the counts (log CFU/mL) at different pH values and incubation times, and bile resistance (% Growth) of the lactic acid bacteria (LAB) strains used in this paper [4].
2.3. Incubations of LAB Strains with Wine Polyphenols
Incubations of bacteria with wine phenolic extract (Provinols™) were carried out as previously described [22]. Briefly, 1 mL of the wine extract solution (0.6 mg/mL) was mixed with 9 mL of inocula of each LAB strain (108 CFU/mL) or sterile saline solution (control). Mixtures of each LAB strain suspension and saline solution (blank) (9:1) were also prepared. The mixtures were incubated at 0, 6 and 24 h, in duplicate, under anaerobic conditions at 37 °C with continuous stirring. Samples were centrifuged (10,000 rpm, for 10 min at 4 °C), and supernatants were kept at −20 °C until undergoing UPLC analyses, which were performed in duplicate.
Phenolic compounds were analysed by UPLC-ESI-MS/MS following the method of Sánchez-Patán et al. [23] with some modifications in the composition of the mobile phases: (a) water/formic acid (99.9:0.1, v/v) and (b) acetonitrile/formic acid (99.9:0.1, v/v). The MS/MS parameters (cone voltage, collision energy and MRM transition) of the 64 phenolic compounds targeted in the present study (mandelic acids, benzoic acids, phenols, hippuric acids, phenylacetic acids, phenylpropionic acids, cinnamic acids, 4-hydroxyvaleric acids, valerolactones and flavan-3-ols) were previously reported [24]. All metabolites were quantified using the calibration curves of their corresponding standards.
2.4. Growth of LAB Strains in the Presence of Phenolic Compounds
Bacterial growth was performed using the method of García-Ruiz et al. [25], slightly modified. Aliquots of 100 μL of (+)-catechin or 3,4-dihydroxyphenylacetic acid solutions (0, 100, 200 and 500 µM) were placed in microplate wells with 100 μL of culture medium (MRS). Then, 20 μL of the diluted LAB strain (inoculum of 1 × 106 CFU/mL) were added to all the microplate wells. The microtitre plates were incubated at 37 °C for 24 h in a Biotek Synergy H1™ multi-mode microplate reader (Winooki, VT, USA). Bacterial growth was determined by reading the absorbance at 600 nm. Assays were conducted twice in triplicate.
2.5. Growth of Pathogen E. coli in the Presence of Free Supernatants (CFS) from LAB Strains
Cell-free supernatants (CFS) of each LAB strain were collected from overnight cultures centrifuged at 4500 rpm for 10 min. After measuring the pH of the CFS (ranging from 5.1 to 5.4), aliquots were taken and adjusted to pH 7 using 1 M NaOH solution. All supernatants were sterilized by filtration (Symta, 0.22 µm PVDF 17 mm pK100). The bacterial growth in the presence of the CFS was measured using the microtitre assay described above. Aliquots of 200 μL of culture medium (MRS), CFS or neutralized CFS were placed in microplate wells. Then, 20 μL of the diluted E. coli strain (inoculum of 1 × 106 CFU/mL) were added to all the microplate wells. Assays were conducted twice in triplicate.
2.6. Cell Culture Assays: LAB Adhesion and Inhibition of E. coli Adherence to Caco-2 Cells
Caco-2™ cells from human colon adenocarcinoma (Caco-2™ ATCC®) were grown and maintained in Dulbecco’s Modified Eagle’s medium (DMEM, Sigma-Aldrich), supplemented with 10% (v/v) foetal calf serum at 37 °C in a 5% CO2/95% air atmosphere. For the adhesion and inhibition experiments, Caco-2 cells were seeded in 24-well tissue plates at 25,000 cells/m2 density and grown over 15 days to obtain a monolayer of differentiated and polarized cells, as previously described by García-Ruiz et al. [4]. Cell culture assays were performed in duplicate and three independent experiments were carried out.
To assess the adhesion of the LAB strains to Caco-2 cells, 0.5 mL of inocula of the LAB strains (108 CFU/mL) was added to Caco-2 cell monolayers previously washed with PBS. After 1 h of incubation at 37 °C in a 5% CO2 atmosphere, the wells were gently washed three times with PBS solution to remove unbound bacteria. Caco-2 cells and adhered bacteria were then detached using a 0.05% trypsin-EDTA solution and bacteria counts were carried out on MRS Agar medium as described above. The adhesion capacity was expressed as the number of adhered bacteria (CFU/mL) relative to the total number of bacteria added initially (% Adhesion = (Adhered bacteria/Total of added bacteria) × 100).
In order to study the effects of LAB on the adhesion of E. coli to Caco-2 cells, two different experiments were carried out: (a) inhibition, to test the ability of the LAB strains to inhibit the adhesion of E. coli; and (b) competition, to test the ability of the LAB strains to compete with E. coli for adhesion to Caco-2 cells. For the inhibition experiments, LAB suspension (108 CFU/mL) was firstly added to Caco-2 cell monolayers, and after 1 h of incubation non-bound bacteria were removed and E. coli suspension (108 CFU/mL) was added to the wells, and the mixture was again incubated for 1 h. Caco-2 cells and adhered bacteria (LAB/E. coli) were then detached and E. coli counting was carried out on TSA plates. The inhibition of the adhesion of E. coli was expressed as a percentage using the following formula: Inhibition of adhesion = 100 × (1 − T1/T2), where T1 and T2 are the percentage of adhesion by E. coli cells in the presence and absence of LAB strains, respectively. The same experimental protocol was carried out for the competition experiments, but adding the LAB and E. coli strains simultaneously (in an initial ratio of 1:1) to the Caco-2 cells followed by incubation for 1 h. Non-bound bacteria were removed and the bacterial counts were carried out as described above.
2.7. Statistical Analysis
A paired-sample t-test was used to evaluate whether the changes in phenolic content of the wine extract (% referred to the values at time 0) after incubations with bacteria was different from 100. Also, one-way analysis of variance (ANOVA) and Tukey test (at p < 0.05) were used for the comparison of the mean values of the LAB growth in each time for each one of time-course graphs. The IBM SPSS program for Windows was used for data processing.
3. Results and Discussion
The selected LAB strains (two oenological LAB strains P. pentosaceus CIAL-86 and L. plantarum CIAL-121 and reference strain L. plantarum CLC 17) were used in the different designed experiments: incubations of LAB strains with wine polyphenols (Section 3.1.), growth of LAB strains in the presence of phenolic compounds (Section 3.2.), and growth of pathogen E. coli and its adherence to Caco-2 cells in the presence of LAB strains (Section 3.3. and Section 3.4.) The three LAB strains used had previously proven good probiotic features in vitro (Table 1) [4]. Other strains belonging to Lactobacillus and Pediococcus genera from different origins have also shown good probiotic properties such as tolerance to gastric conditions and bile tolerance [4,26,27].
3.1. Metabolism of Wine Polyphenols by LAB Strains
The capacity of three selected LAB strains (the probiotic L. plantarum CLC 17, and the oenological strains P. pentosaceus CIAL-86 and L. plantarum CIAL-121) to metabolize wine polyphenols was assessed through their incubation with a commercial wine phenolic extract (Provinols™) under nutrient-restricted culture conditions. Among different phenolic compounds targeted (mandelic acids, benzoic acids, phenols, hippuric acids, phenylacetic acids, phenylpropionic acids, cinnamic acids, 4-hydroxyvaleric acids, valerolactones and flavan-3-ols), a total of 15 compounds were previously quantified by UHPLC-MS/MS analysis [22]. Table 2 shows the data corresponding to the sum of the concentrations of individual compounds at 0, 6 and 24 h of incubation. Out of the three LAB strains tested, only L. plantarum CLC 17 produced significant increases in the concentration of phenolic compounds after 24 h of incubation (133.2% in relation to t = 0 h). Therefore, phenolic-degrading enzymatic activities might be strain-dependent (i.e., L. plantarum CLC 17), as other potential probiotic bacteria belonging to the same species were not active on wine polyphenols (i.e., L. plantarum CIAL-121). However, additional studies to shed light on the enzymatic activities of LAB will be intended to be carried out. Previous studies with the same wine phenolic extract used in this study also reported release of the same phenolic acids, after batch fermentations [18] or gastrointestinal digestion simulation [28] inoculated with human faecal microbiota.
Table 2.
Sum (∑) of the concentrations (C, ppm) of individual phenolic compounds (gallic acid, caffeic acid, ferulic acid, protocatechuic acid, vanillic acid, salicylic acid, 3-O-methyl gallic acid, syringic acid, phthalic acid, 4-hydroxybenzoic acid, p-coumaric acid, (+)-catechin, (−)-epicatechin, procyanidin B1 and procyanidin B2) before and after incubation of the wine extract with LAB strains.
3.2. Effects of Phenolic Compounds on the LAB Growth
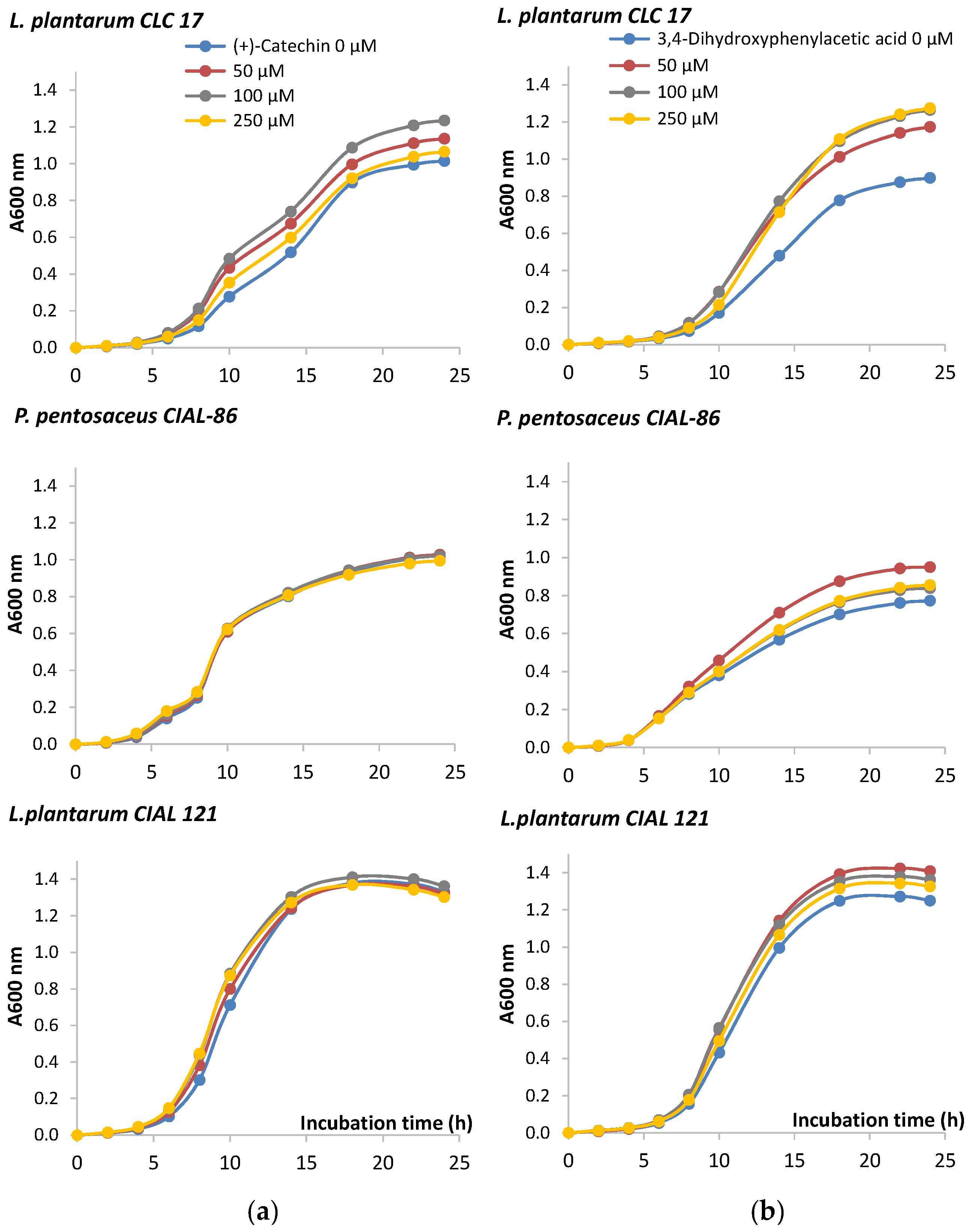
In order to look more deeply into the effects of wine phenolic compounds and their metabolites on bacteria performance, the growth of the probiotic strain L. plantarum CLC 17 and the two oenological LAB strains P. pentosaceus CIAL-86 and L. plantarum CIAL-121 was monitored in the presence of (+)-catechin, a main phenolic compound present in wine, and 3,4-dihydroxyphenylacetic acid, a microbial-derived phenolic metabolite whose concentration in faeces had been reported to significantly increase after moderate consumption of red wine [12]. Time-course graphs indicated a certain stimulatory effect of the growth of the three strains in the presence of 3,4-dihydroxyphenylacetic acid (Figure 1). On the other hand, the monomer (+)-catechin only promoted the growth of L. plantarum CLC 17 (Figure 1). Results of the one way analysis of variance (ANOVA) and Tukey test (at p < 0.05) did not show significant differences in the most of the mean values, except in the case of the L. plantarum CLC 17 in the presence of 3,4-dihydroxyphenylacetic acid (50, 100 and 250 µM) in comparison to the absence of the compound, from 6 to 24 h (Figure 1b). Therefore, our results confirmed that the chemical structure of polyphenols did indeed influence their effects on bacterial growth. In relation to this, other authors observed that flavanols with galloyl moiety ((−)-epigallocatechin, (−)-epicatechin-3-gallate and (−)-epigallocatechin-3-gallate) exhibited more activity on bacteria growth than those without galloyl moiety (catechins and (−)-epicatechin) [29]. Also, the microbial potency of polyphenols towards bacteria growth has also been reported to be dependent upon bacterial strain, species and genera [30], as we have also observed in our study.
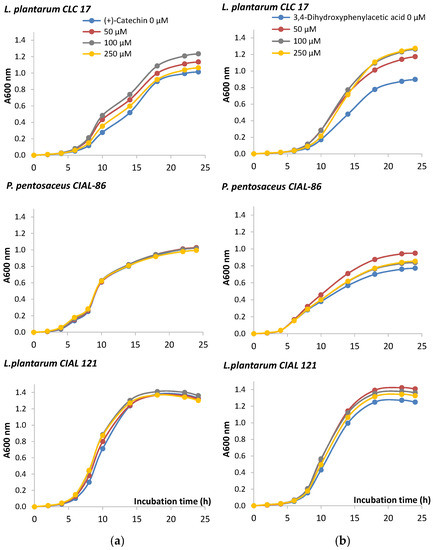
Figure 1.
Growth curves of isolated LAB in the absence and presence of (+)-catechin (a) and 3,4-dihydroxyphenylacetic acid (b). The curve was developed from absorbance measures (OD:600) up to 24 h.
3.3. Effects of the Cell-Free Supernatants (CFS) from the LAB Strains on Growth of Pathogen E. coli
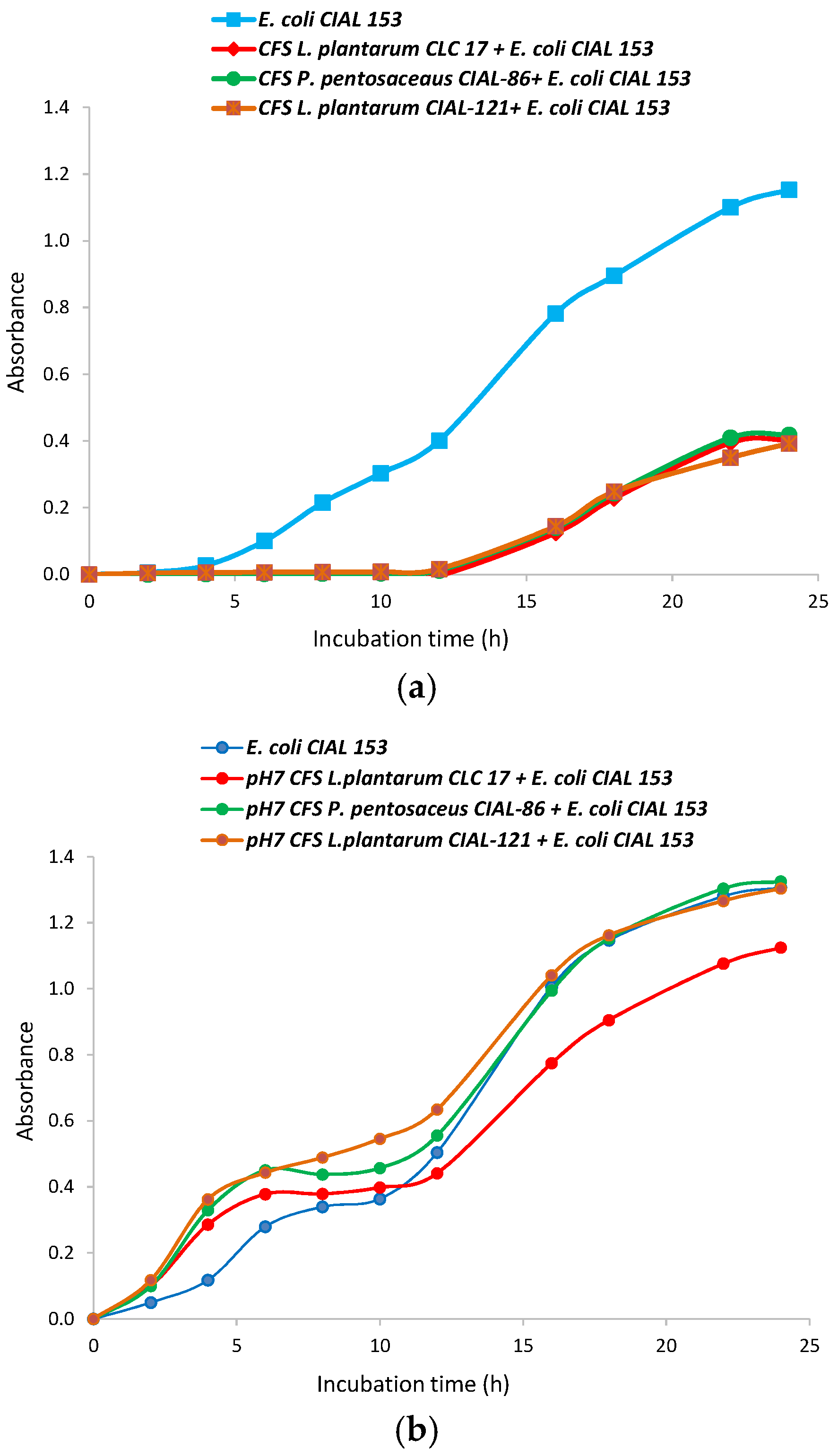
Bacteria cell-free supernatants of LAB strains have been reported to exhibit functions similar to the living bacteria from which they were derived, and to reduce the infection risk of the use of probiotic bacteria in patients with depressed immune systems [31]. So, CFS from the LAB strains were prepared in MRS broth and their antibacterial activity against E. coli CIAL-153 was evaluated (Figure 2). For the three LAB studied, CFS delayed the bacterial lag phase (from 5 to 12 h), and a strong inhibition of pathogen bacteria growth was observed (Figure 2a). Other authors have also reported an extension of the bacterial lag phase and lower growth rates of pathogen bacteria in the presence of the CFS from strains belonging to the Lactobacillus, Bifidobacterium, Lactococcus, Streptococcus and Bacillus genera [32,33]. In agreement with them, we hypothesized that these antimicrobial effects were mainly due to the organic acids that were produced in significant quantities (and consequent lowering of pH) as a result of the ability of LAB strains to ferment carbohydrates. In addition, Gram-negatives pathogens, such as E. coli, tend to be more sensitive to organic acids than to bacteriocins [33], which explains the observed strong inhibition. We confirmed that the pH of the CFS from oenological LAB strains (P. pentosaceus CIAL-86 and L. plantarum CIAL-121), and the probiotic strain (L. plantarum CLC 17), had acidic pH values of 5.25, 5.14 and 5.11, respectively.
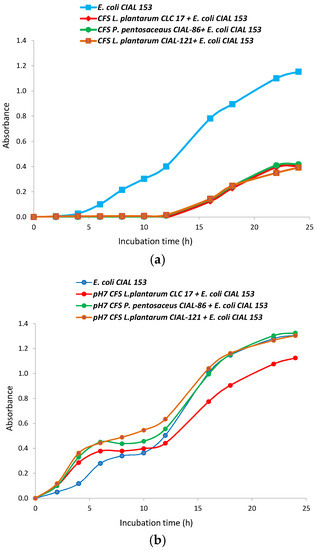
Figure 2.
Growth curves of E. coli CIAL-153 in the presence of cell-free supernatants of L. plantarum CLC 17, P. pentosaceus CIAL-86 and L. plantarum CIAL-121 strains before (a) and after (b) being neutralized at pH 7.
Moreover, with the aim of researching the effect of other antimicrobial substances, such as bacteriocins, in addition to organic acids on E. coli growth, CFS were adjusted to pH 7 and their antibacterial activity against E. coli CIAL-153 was again evaluated (Figure 2b). Neutralization of the supernatants from all the LAB strains counteracted the antagonistic effects of the acid CFS against the pathogen strain, so the lag phase was similar to that of a standard growth curve and a significant increase in the growth of the pathogen was observed. Other authors have also observed that the neutralization of CFS reduced the antimicrobial activity on pathogen viability and growth, but they still observed some effects [32,33]. In our case, neutralized supernatants from the L. plantarum CLC 17 strain still exhibited some inhibition of the growth of E. coli CIAL-153, which suggested that this strain produces other antibacterial active compounds against E. coli. Arena et al. [34] reported that antimicrobial activity is mainly strain-specific rather than genus/species-specific and provided evidence that several of the 79 screened L. plantarum strains possess a significant ability to contrast various pathogenic bacteria, including both Gram-negative and Gram-positive species.
3.4. Effects of LAB on Adherence of Pathogen E. coli to Caco-2 Cells
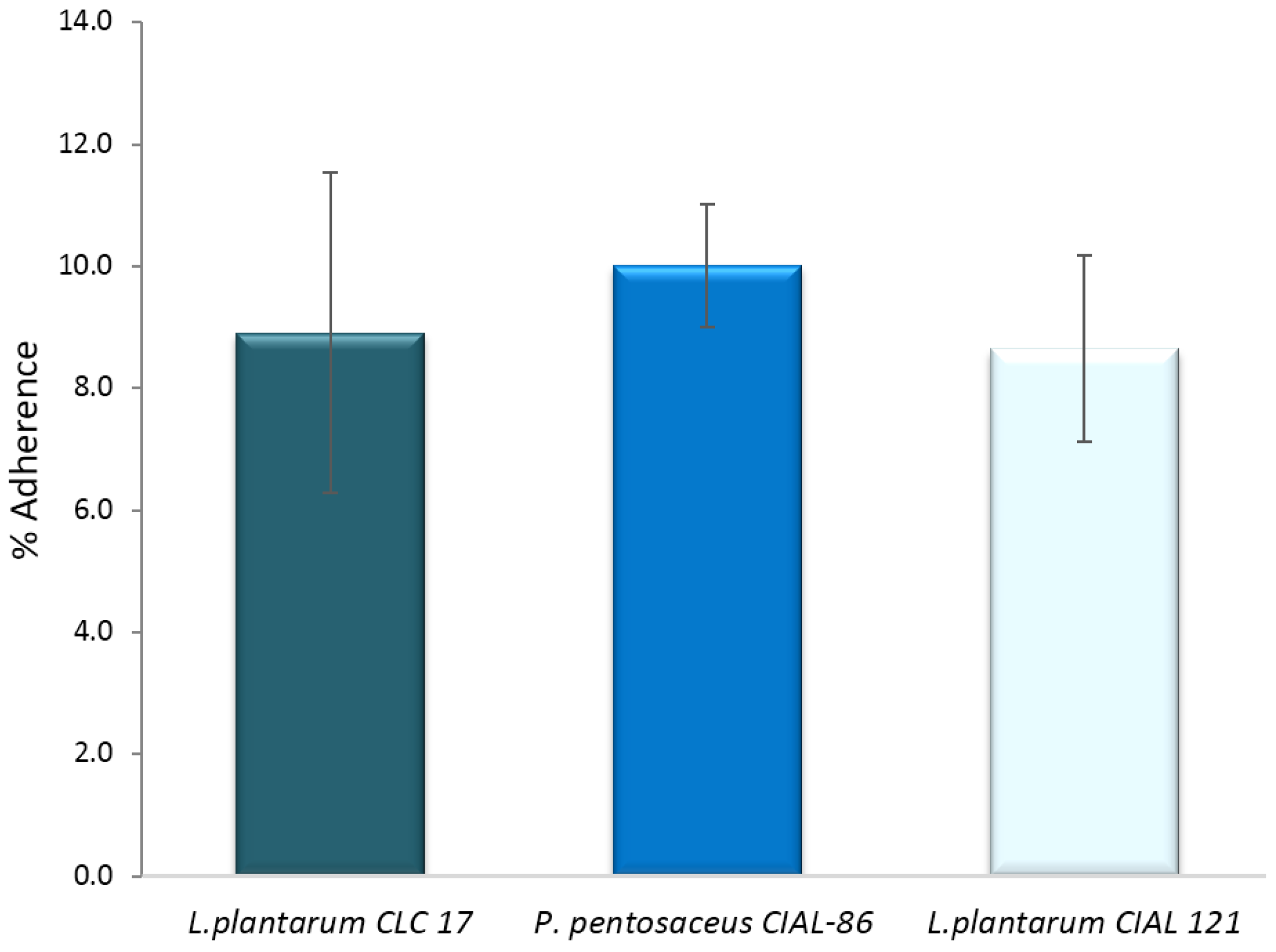
An important property of probiotic candidates is their ability to adhere to intestinal mucosa, which excludes pathogens from cell adherence and infection progression. Initially, we investigated the ability of the three LAB strains (L. plantarum CLC 17, P. pentosaceus CIAL-86 and L. plantarum CIAL-121) to adhere to human intestinal Caco-2 cells because this cellular model expresses morphological and functional differentiation in vitro and shows characteristics of mature enterocytes. Adhesion levels to Caco-2 cells of the three LAB strains ranged from 8.65% to 10.01% (Figure 3) and were in line with those obtained previously [4] and in the range of other probiotics previously reported in the literature under in vitro conditions [35,36].
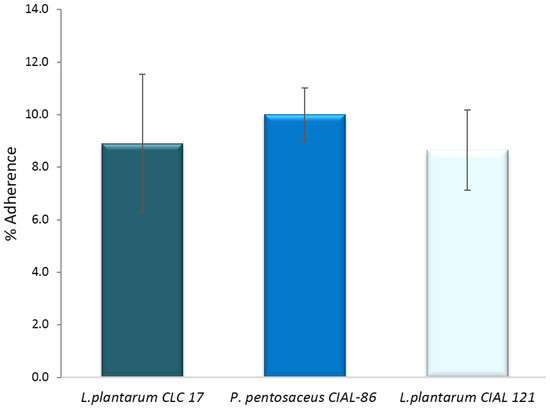
Figure 3.
Adherence (%) of P. pentosaceus CIAL-86, L. plantarum CIAL-121 and L. plantarum CLC 17 strains to Caco-2 cells. Results are shown as media ± standard deviation.
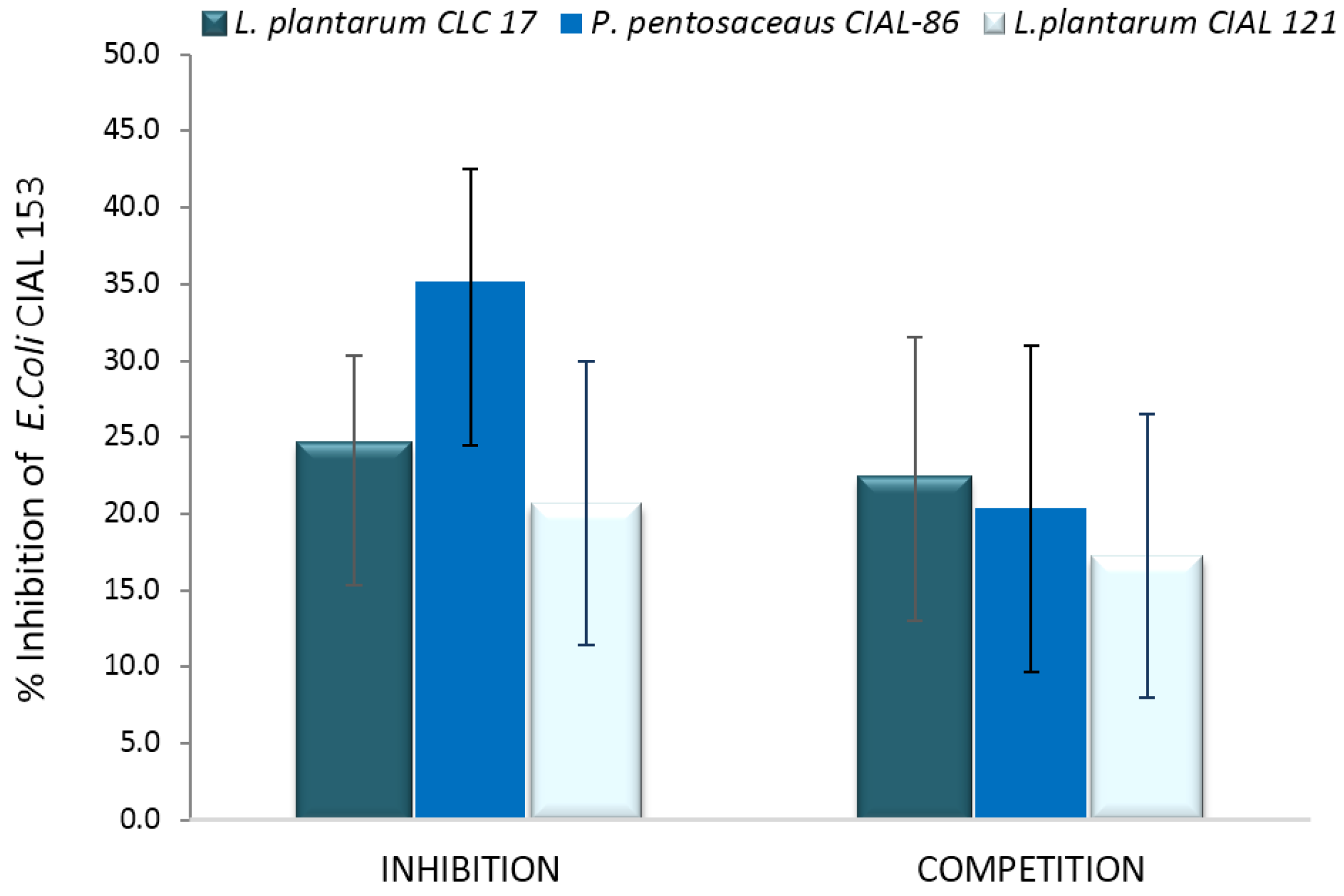
Having confirmed the ability of LAB strains to adhere to Caco-2 cells, the adhesion of E. coli CIAL-153 to these intestinal cells was assessed in the presence of the different LAB strains. Initially, it was found that the adhesion of E. coli CIAL-153 to Caco-2 cells on their own was 8.51% ± 1.83%. The inhibition of adherence of E. coli CIAL-153 to Caco-2 cells for probiotic LAB strains in the anti-adhesion assays (competition and inhibition) is shown in Figure 4. Pre-incubation of Caco-2 cells with LAB strains prior to the addition of E. coli CIAL-153 (inhibition assay) produced a notable inhibition of the adhesion of E. coli to the intestinal cells for three strains in respect to the control (absence of LAB strains). P. pentosaceus CIAL-86 was the most effective strain in inhibiting the adhesion of E. coli CIAL-153 (>35%), while L. plantarum CIAL-121 showed similar inhibition values to those observed in the reference probiotic strain, L. plantarum CLC 17 (20.7% and 22.6%, respectively) (Figure 4). These percentages were similar when Caco-2 cells were incubated at the same time with both E. coli and probiotic LAB strains (competition assay), ranging from 17% to 22% in respect to the control (absence of LAB strains). The high values from the inhibition experiment could indicate an effective competition of LAB strains against E. coli CIAL-153 for common adhesion receptors [37] or other anti-adhesion factors [38]. Thus, the ability to inhibit the adhesion of E. coli CIAL-153 to Caco-2 cells appeared to be influenced by LAB strains, which suggested a certain pathogen-LAB specificity as indicated by other authors [35].
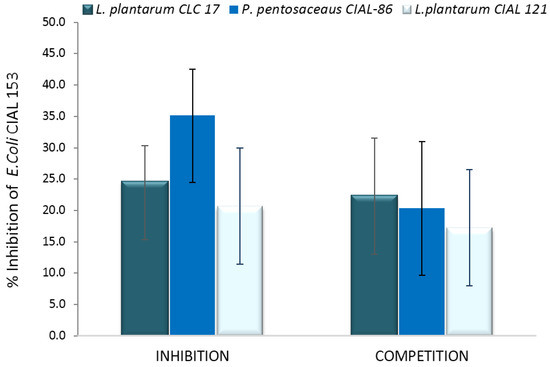
Figure 4.
Inhibition (%) of adherence (%) of E. coli CIAL-153 to Caco-2 cells in the presence of L. plantarum CLC 17, P. pentosaceus CIAL-86 and L. plantarum CIAL-121 in anti-adhesion assays (competition and inhibition). Results are shown as media ± standard deviation.
4. Conclusions
In vivo reports suggest that wine polyphenols exert an essential impact on intestinal microbiota growth and functionality (see [9] for review). However, an important question that remains unsolved is whether these benefits may be enhance by the concomitant interactions by wine polyphenols and probiotics at the gut level. This paper investigates some new metabolic features and probiotic characteristics of oenological lactic acid bacteria, in particular P. pentosaceus CIAL-86 and L. plantarum CIAL-121, based on their interaction with polyphenols. Neither of these two oenological bacteria was able to metabolize wine polyphenols, at least in the conditions used in this study, although this metabolic potential migth be strain-dependent, as the probiotic reference strain L. plantarum CLC 17 was found to be effective in metabolizing wine polyphenols. However, growth of both oenological (P. pentosaceus CIAL-86 and L. plantarum CIAL-121) and reference (L. plantarum CLC 17) strains was stimulated in the presence of wine phenolic compounds (i.e., (+)-catechin) and wine-derived phenolic metabolites (i.e., 3,4-dihydroxyphenylacetic acid), although no clear dose-dependent effect was observed. Bacteria cell-free supernatants from the three LAB strains delayed and inhibited almost completely the E. coli CIAL-153 growth, which may be mainly attributed to the presence of organic acids derived from the metabolism of carbohydrates by LAB. In relation to their adhesion to intestinal cells, the three LAB strains showed a high adhesion percentage, especially P. pentosaceus CIAL-86. Moreover, pre-incubation of Caco-2 cells with LAB strains prior to the addition of E. coli CIAL-153 produced a notable inhibition of the adhesion of E. coli to the intestinal cells. Nevertheless, the effect of selected lactic acid bacteria on the growth and adhesion to intestinal cells of other gut pathogenic bacteria should be investigated. To our knowledge, there are very few reports considering probiotic features of LAB isolated from wine such as the ones investigated here, which emphasizes the novelty of these results. Overall, these in vitro results confirm the potential of oenological LAB strains as probiotics, with the aim of developing general nutritional strategies and designing specific dietary recommendations based on the combination of active phenolic compounds/extracts and probiotics, thus contributing to the ultimate goal of promoting intestinal health. Nevertheless, further in vitro and in vivo investigations are still necessary in order to confirm these potential beneficial effects.
Acknowledgments
The authors are grateful to Laura Laguna for her valuable assistance in the statistical treatment. The study was supported by the projects AGL2012-40172-C02-01 and AGL2015-64522-C2-1-R of the Spanish Ministry of Economy and Competitiveness (MINECO) and ALIBIRD S2013/ABI-2728 of the Comunidad de Madrid, as well as COST Action POSITIVe FA1403. IG-S is the recipient of an FPU contract from the Spanish Ministry of Education. AE-F is the recipient of an FPI contract from the Spanish MINECO (Project AGL2012-40172-C02-01).
Author Contributions
Dolores González de Llano and Begoña Bartolomé conceived and designed the study, analyzed the data and wrote the paper. Irene Gil-Sánchez, Adelaida Esteban-Fernández, Alba M. Ramos and Carolina Cueva performed the experiments. M. Victoria Moreno-Arribas participated in the design and critical revision. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sleator, R.D. Designer probiotics: Development and applications in gastrointestinal health. World J. Gastrointest. Pathophysiol. 2015, 6, 73–78. [Google Scholar] [PubMed]
- De Palencia, P.F.; Werning, M.L.; Sierra-Filardi, E.; Duenas, M.T.; Irastorza, A.; Corbi, A.L.; Lopez, P. Probiotic properties of the 2-substituted (1,3)-β-d-glucan-producing bacterium Pediococcus parvulus 2.6. Appl. Environ. Microbiol. 2009, 75, 4887–4891. [Google Scholar] [CrossRef] [PubMed]
- Foligne, B.; Dewulf, J.; Breton, J.; Claisse, O.; Lonvaud-Funel, A.; Pot, B. Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by Oenococcus oeni. Int. J. Food Microbiol. 2010, 140, 136–145. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; González de Llano, D.; Esteban-Fernández, A.; Requena, T.; Bartolomé, B.; Moreno-Arribas, M.V. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 2014, 44, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Amaretti, A.; Leonardi, A.; Raimondi, S.; Simone, M.; Quartieri, A. Potential impact of probiotic consumption on the bioactivity of dietary phytochemicals. J. Agric. Food Chem. 2013, 61, 9551–9558. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Oliver, C.M.; Weerakkody, R.; Singh, T.; Conlon, M.; Borges, G.; Sanguansri, L.; Lockett, T.; Roberts, S.A.; Crozier, A.; et al. Chronic administration of a microencapsulated probiotic enhances the bioavailability of orange juice flavanones in humans. Free Radic. Biol. Med. 2015, 84, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, I.; Espinosa-Martos, I.; Rodríguez, J.M.; Jiménez-Girón, A.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Moderate consumption of red wine can modulate human intestinal inflammatory response. J. Agric. Food Chem. 2014, 62, 10567–10575. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Cueva, C.; Muñoz-González, I.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. Studies on Modulation of Gut Microbiota by Wine Polyphenols: From Isolated Cultures to Omic Approaches. Antioxidants 2015, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Boto-Ordoñez, M.; Rothwell, J.A.; Andrés-Lacueva, C.; Manach, C.; Scalbert, A.; Urpi-Sarda, M. Prediction of the wine polyphenol metabolic space: An application of the Phenol-Explorer database. Mol. Nutr. Food Res. 2014, 58, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, I.; Jiménez-Girón, A.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Profiling of Microbial-Derived Phenolic Metabolites in Human Feces after Moderate Red Wine Intake. J. Agric. Food Chem. 2013, 61, 9470–9479. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Urpi-Sarda, M.; Sánchez-Patán, F.; Llorach, R.; Garrido, I.; Gómez-Cordovés, C.; Andrés-Lacueva, C.; Bartolomé, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233–253. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; Del Rio, D.; Crozier, A. Bioavailability, bioactivity and impact on health of dietary flavonoids and related compounds: An update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of Polyphenols and Polyphenol-Rich Dietary Sources on Gut Microbiota Composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Sánchez-Patán, F.; Monagas, M.; Walton, G.E.; Gibson, G.R.; Martín-Álvarez, P.J.; Bartolome, B.; Moreno-Arribas, M.V. In vitro fermentation of grape seed flavan-3-ol fractions by human faecal microbiota: Changes in microbial groups and phenolic metabolites. FEMS Microbiol. Ecol. 2013, 83, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macia, A.; Motilva, M.J. Metabolic and microbial modulation of the large intestine ecosystem by non-absorbed diet phenolic compounds. A review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Patán, F.; Cueva, C.; Monagas, M.; Walton, G.E.; Gibson, M.G.R.; Quintanilla-López, J.E.; Lebrón-Aguilar, R.; Marín-Álvarez, P.J.; Moreno-Arribas, M.V.; Bartolomé, B. In vitro fermentation of a red wine extract by human gut microbiota: Changes in microbial groups and formation of phenolic metabolites. J. Agric. Food Chem. 2012, 60, 2136–2147. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, M.V.; Polo, M.C. Occurrence of lactic acid bacteria and biogenic amines in biologically aged wines. Food Microbiol. 2008, 25, 875–881. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Tabasco, R.; Requena, T.; Claisse, O.; Lonvaud-Funel, A.; Bartolome, B.; Moreno-Arribas, M.V. Genetic diversity of Oenoccoccus oeni isolated from wines treated with phenolic extracts as antimicrobial agents. Food Microbiol. 2013, 36, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Langa, S.; Reviriego, C.; Jimínez, E.; Marín, M.L.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Human milk is a source of lactic acid bacteria for the infant gut. J. Pediatr. 2003, 143, 7540–7548. [Google Scholar] [CrossRef] [PubMed]
- González de Llano, D.; Gil-Sánchez, I.; Esteban-Fernández, A.; Ramos, M.A.; Fernández Díaz, M.; Cueva, C.; Moreno-Arribas, M.V.; Bartolomé, B. Reciprocal beneficial effects between wine polyphenols and probiotics: An exploratory study. Eur. Food Res. Technol. 2016. [Google Scholar] [CrossRef]
- Sánchez-Patán, F.; Monagas, M.; Moreno-Arribas, M.V.; Bartolomé, B. Determination of microbial phenolic acids in human faeces by UPLC-ESI-TQ MS. J. Agric. Food Chem. 2011, 59, 2241–2247. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Girón, A.; Queipo-Ortuño, M.I.; Boto-Ordoñez, M.; Muñoz-González, I.; Sánchez-Patan, F.; Monagas, M.; Martín-Álvarez, P.J.; Murri, M.; Tinahones, F.J.; Andrés-Lacueva, C.; et al. Comparative study of microbial-derived phenolic metabolites in human feces after intake of gin, red wine, and dealcoholized red wine. J. Agric. Food Chem. 2013, 61, 3909–3915. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bartolomé, B. Comparative study of the inhibitory effects of wine polyphenols on the growth of enological lactic acid bacteria. Int. J. Food Microbiol. 2011, 145, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.; Grimmer, S.; Naterstad, K.; Axelsson, L. In vitro testing of commercial and potential probiotic lactic acid bacteria. Int. J. Food Microbiol. 2012, 153, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Turchi, B.; Mancini, S.; Fratini, F.; Pedonese, F.; Nuvoloni, R.; Bertelloni, F.; Ebani, V.V.; Cerri, D. Preliminary evaluation of probiotic potential of Lactobacillus plantarum strains isolated from Italian food products. World J. Microbiol. Biotechnol. 2013, 29, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Van de Wiele, T.; Jiménez-Girón, A.; Muñoz-González, I.; Martín-Alvarez, P.J.; Moreno-Arribas, M.V.; Bartolomé, B.; Peláez, C.; Martínez-Cuesta, M.C.; Requena, T. Lactobacillus plantarum IFPL935 impacts colonic metabolism in a simulator of the human gut microbiota during feeding with red wine polyphenols. Appl. Microbiol. Biotechnol. 2014, 98, 6805–6815. [Google Scholar] [CrossRef] [PubMed]
- Sourabh, A.; Kanwar, S.S.; Sud, R.G.; Ghabru, A.; Sharma, O.P. Influence of phenolic compounds of Kangra tea [Camellia sinensis (L) O Kuntze] on bacterial pathogens and indigenous bacterial probiotics of Western Himalayas. Braz. J. Microbiol. 2013, 44, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Henriksson, A.; Nilsson, C.; Mitchell, H. Synergistic effect of green tea extract and probiotics on the pathogenic bacteria, Staphylococcus aureus and Streptococcus pyogenes. World J. Microbiol. Biotechnol. 2008, 24, 1837–1842. [Google Scholar] [CrossRef]
- Prisciandaro, L.D.; Geier, M.S.; Chua, A.E.; Butler, R.N.; Cummins, A.G.; Sander, G.R.; Howarth, G.S. Probiotic factors partially prevent changes to caspases 3 and 7 activation and transepithelial electrical resistance in a model of 5-fluorouracil-induced epithelial cell damage. Support. Care Cancer 2012, 20, 3205–3210. [Google Scholar] [CrossRef] [PubMed]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Likotrafiti, E.; Valavani, P.; Argiriou, A.; Rhoades, J. In vitro evaluation of potential antimicrobial synbiotics using Lactobacillus kefiri isolated from kefir grains. Int. Dairy J. 2015, 45, 23–30. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Gueimonde, M.; Jalonen, L.; He, F.; Hiramatsu, M.; Salminen, S. Adhesion and competitive inhibition and displacement of human enteropathogens by selected lactobacilli. Food Res. Int. 2006, 39, 467–471. [Google Scholar] [CrossRef]
- Bustos, I.; Garcia-Cayuela, T.; Hernández-Ledesma, B.; Pelaez, C.; Requena, T.; Martínez-Cuesta, M.C. Effect of flavan-3-ols on the adhesion of potential probiotic lactobacilli to intestinal cells. J. Agric. Food Chem. 2012, 60, 9082–9088. [Google Scholar] [CrossRef] [PubMed]
- Leahy, S.C.; Higgins, D.G.; Fitzgerald, G.F.; Sinderen, D.V. Getting better with bifidobacteria. J. Appl. Microbiol. 2005, 98, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Abedi, D.; Feizizadeh, S.; Akbari, V.; Jafarian-Dehkordi, A. In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on Escherichia coli. Res Pharm. Sci. 2013, 8, 260–268. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).