High-Definition Transcranial Direct Current Stimulation in the Right Ventrolateral Prefrontal Cortex Lengthens Sustained Attention in Virtual Reality

 ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
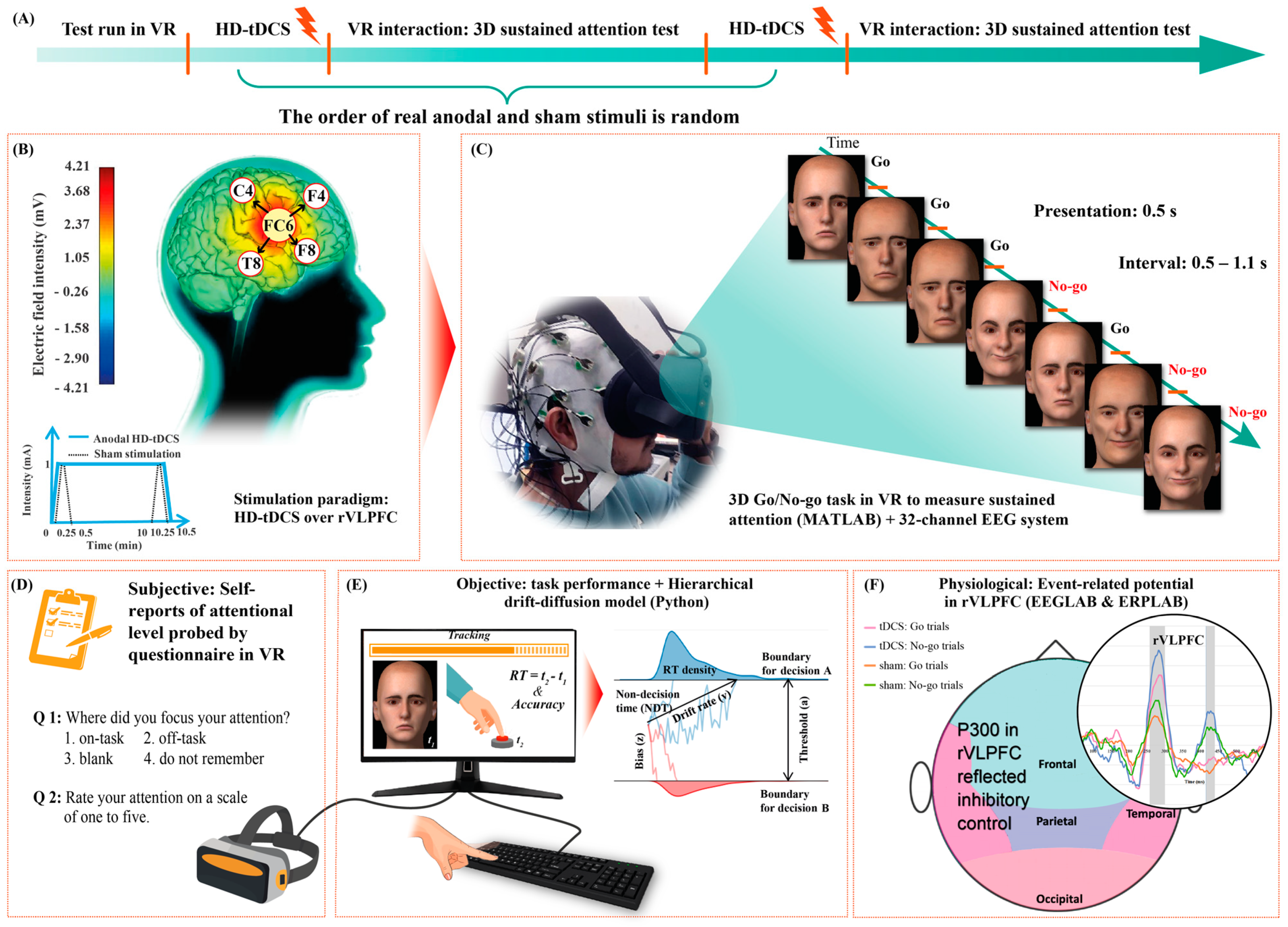
2. Materials and Methods
2.1. Participants
2.2. Procedure and Stimulation
2.3. VR and 3D GNG Task
2.4. Behavioral Analyses
2.5. EEG and ERP
2.6. Statistical Analyses
3. Results
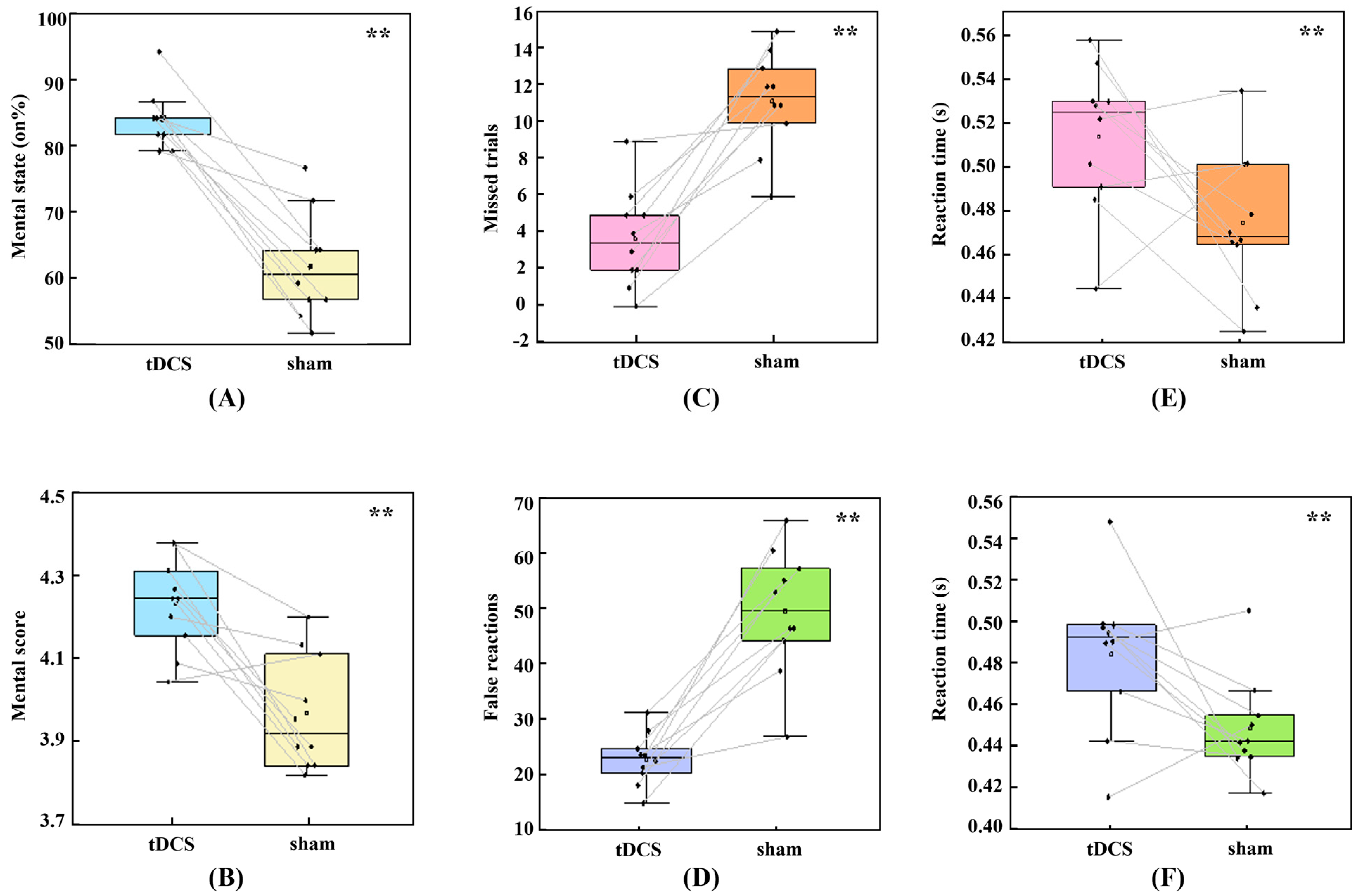
3.1. Subjective Experience
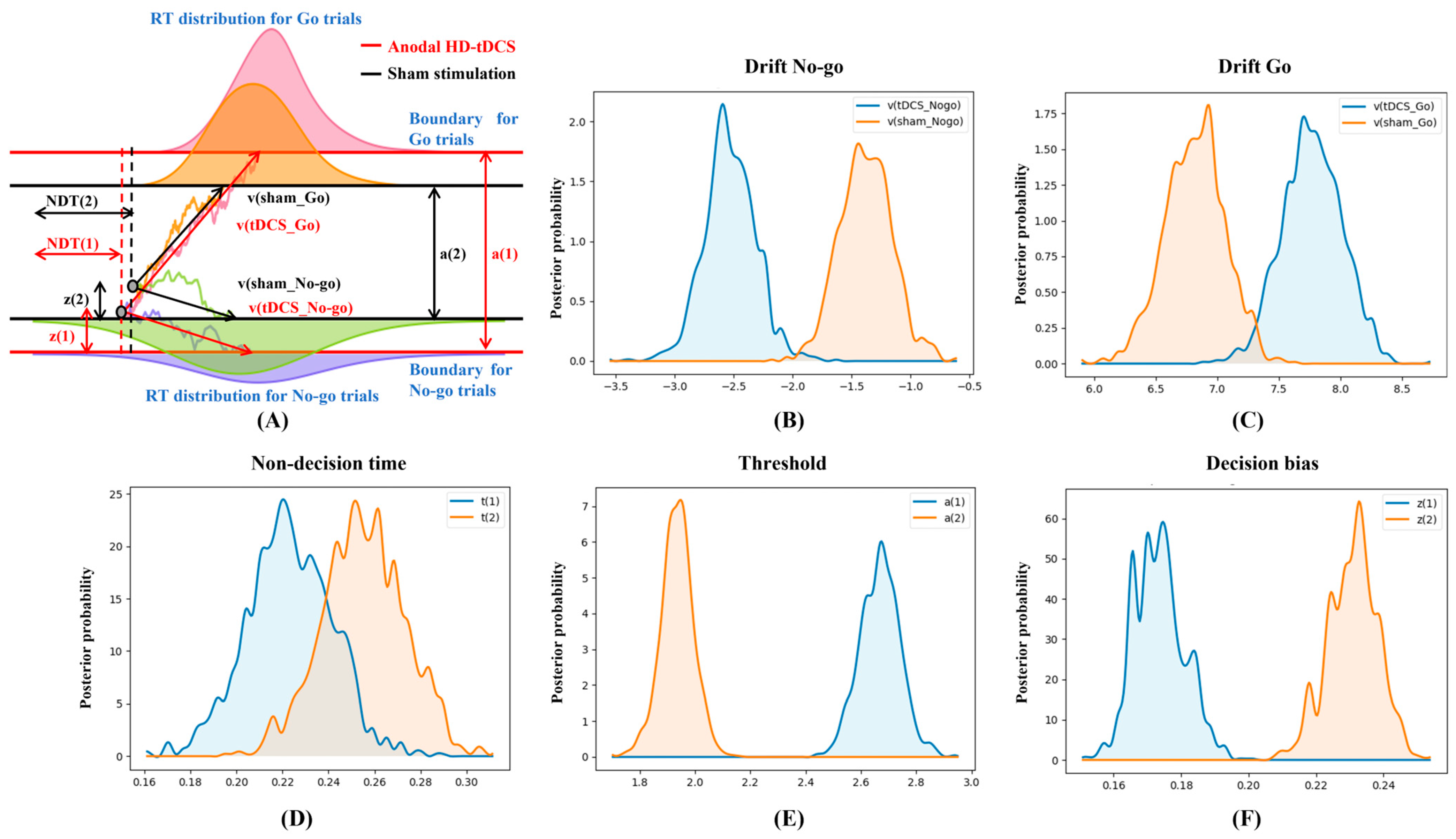
3.2. Behavioral Performance and HDDM Metrics
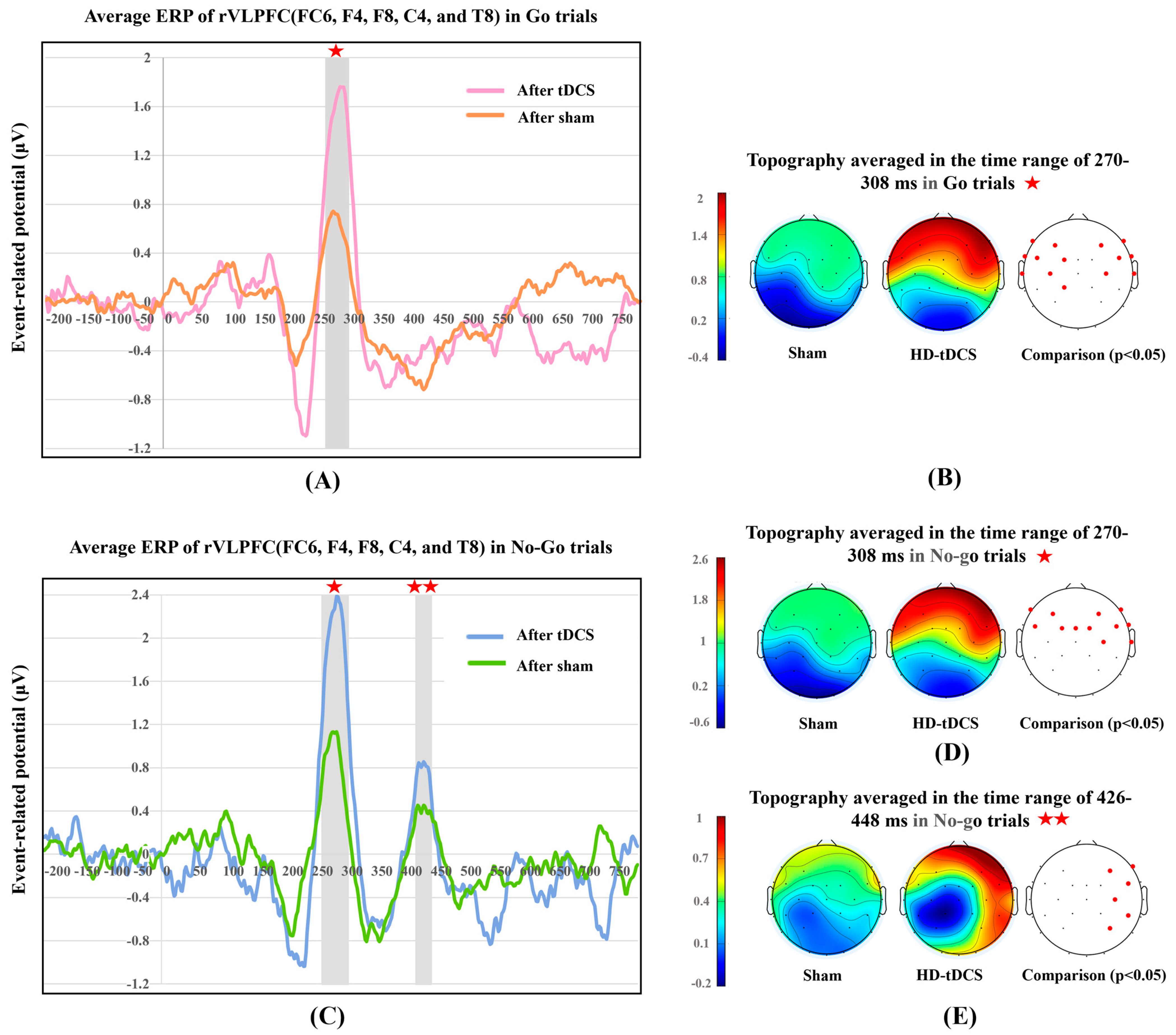
3.3. ERP Components
3.3.1. P300 in Go and No-Go Trials
3.3.2. P400 in No-Go Trials
3.4. Hierarchical Multiple Regression
4. Discussion
4.1. Methodology of Embodying and Motivating 3D Sustained Attention
4.2. Comprehensive Evidence for Extended Attention in VR
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiong, J.; Hsiang, E.-L.; He, Z.; Zhan, T.; Wu, S.-T. Augmented reality and virtual reality displays: Emerging technologies and future perspectives. Light Sci. Appl. 2021, 10, 216. [Google Scholar] [CrossRef]
- Zhan, T.; Yin, K.; Xiong, J.; He, Z.; Wu, S.-T. Augmented reality and virtual reality displays: Perspectives and challenges. iScience 2020, 23, 101397. [Google Scholar] [CrossRef]
- Qiao, J.; Li, X.; Wang, Y.; Wang, Y.; Li, G.; Lu, P.; Wang, S. The infraslow frequency oscillatory transcranial direct current stimulation over the left dorsolateral prefrontal cortex enhances sustained attention. Front. Aging Neurosci. 2022, 14, 879006. [Google Scholar] [CrossRef]
- Andrillon, T.; Burns, A.; Mackay, T.; Windt, J.; Tsuchiya, N. Predicting lapses of attention with sleep-like slow waves. Nat. Commun. 2021, 12, 3657. [Google Scholar] [CrossRef]
- Litleskare, S. The relationship between postural stability and cybersickness: It’s complicated–an experimental trial assessing practical implications of cybersickness etiology. Physiol. Behav. 2021, 236, 113422. [Google Scholar] [CrossRef]
- Nie, G.-Y.; Duh, H.B.-L.; Liu, Y.; Wang, Y. Analysis on mitigation of visually induced motion sickness by applying dynamical blurring on a user’s retina. IEEE Trans. Vis. Comput. Graph. 2019, 26, 2535–2545. [Google Scholar] [CrossRef]
- Fernandes, A.S.; Feiner, S.K. Combating VR sickness through subtle dynamic field-of-view modification. In Proceedings of the 2016 IEEE Symposium on 3D User Interfaces (3DUI), Greenville, SC, USA, 19–20 March 2016; pp. 201–210. [Google Scholar] [CrossRef]
- Lin, J.-W.; Duh, H.B.-L.; Parker, D.E.; Abi-Rached, H.; Furness, T.A. Effects of field of view on presence, enjoyment, memory, and simulator sickness in a virtual environment. In Proceedings of the IEEE Virtual Reality 2002, Orlando, FL, USA, 24–28 March 2002; pp. 164–171. [Google Scholar]
- Thair, H.; Holloway, A.L.; Newport, R.; Smith, A.D. Transcranial direct current stimulation (tDCS): A beginner’s guide for design and implementation. Front. Neurosci. 2017, 11, 641. [Google Scholar] [CrossRef]
- Zhao, H.; Qiao, L.; Fan, D.; Zhang, S.; Turel, O.; Li, Y.; Li, J.; Xue, G.; Chen, A.; He, Q. Modulation of brain activity with noninvasive transcranial direct current stimulation (tDCS): Clinical applications and safety concerns. Front. Psychol. 2017, 8, 685. [Google Scholar] [CrossRef]
- Mashour, G.A.; Pal, D.; Brown, E.N. Prefrontal cortex as a key node in arousal circuitry. Trends Neurosci. 2022, 45, 722–732. [Google Scholar] [CrossRef]
- Takahashi, M.; Ikegami, M. Differential Frontal Activation during Exogenous and Endogenous Orientation of Visuospatial Attention. Neuropsychobiology 2008, 58, 55–64. [Google Scholar] [CrossRef]
- Levy, B.J.; Wagner, A.D. Cognitive control and right ventrolateral prefrontal cortex: Reflexive reorienting, motor inhibition, and action updating. Ann. N. Y. Acad. Sci. 2011, 1224, 40–62. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.C.; Mullins, T.S.; Heinrich, M.D.; Witkiewitz, K.; Alfred, B.Y.; Hansberger, J.T.; Clark, V.P. Transcranial direct current stimulation facilitates category learning. Brain Stimul. 2020, 13, 393–400. [Google Scholar] [CrossRef]
- Coffman, B.A.; Trumbo, M.C.; Clark, V.P. Enhancement of object detection with transcranial direct current stimulation is associated with increased attention. BMC Neurosci. 2012, 13, 108. [Google Scholar] [CrossRef]
- Campanella, S.; Schroder, E.; Monnart, A.; Vanderhasselt, M.-A.; Duprat, R.; Rabijns, M.; Kornreich, C.; Verbanck, P.; Baeken, C. Transcranial direct current stimulation over the right frontal inferior cortex decreases neural activity needed to achieve inhibition: A double-blind ERP study in a male population. Clin. EEG Neurosci. 2017, 48, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Hampshire, A.; Chamberlain, S.R.; Monti, M.M.; Duncan, J.; Owen, A.M. The role of the right inferior frontal gyrus: Inhibition and attentional control. Neuroimage 2010, 50, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Ghodratitoostani, I.; Delbem, A.C.; Ali, A.; Datta, A. Influence of gender-related differences in transcranial direct current stimulation: A Computational Study. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and biology Society (EMBC), Berlin, Germany, 23–27 July 2019; pp. 5196–5199. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar] [CrossRef]
- Villamar, M.F.; Wivatvongvana, P.; Patumanond, J.; Bikson, M.; Truong, D.Q.; Datta, A.; Fregni, F. Focal modulation of the primary motor cortex in fibromyalgia using 4 × 1-ring high-definition transcranial direct current stimulation (HD-tDCS): Immediate and delayed analgesic effects of cathodal and anodal stimulation. J. Pain 2013, 14, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Ruch, S.; Fehér, K.; Homan, S.; Morishima, Y.; Mueller, S.M.; Mueller, S.V.; Dierks, T.; Grieder, M. Bi-temporal anodal transcranial direct current stimulation during slow-wave sleep boosts slow-wave density but not memory consolidation. Brain Sci. 2021, 11, 410. [Google Scholar] [CrossRef] [PubMed]
- Sreeraj, V.S.; Dinakaran, D.; Parlikar, R.; Chhabra, H.; Selvaraj, S.; Shivakumar, V.; Bose, A.; Narayanaswamy, J.C.; Venkatasubramanian, G. High-definition transcranial direct current simulation (HD-tDCS) for persistent auditory hallucinations in schizophrenia. Asian J. Psychiatry 2018, 37, 46–50. [Google Scholar] [CrossRef]
- Friedman, N.P.; Robbins, T.W. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef]
- Kessler, S.K.; Turkeltaub, P.E.; Benson, J.G.; Hamilton, R.H. Differences in the experience of active and sham transcranial direct current stimulation. Brain Stimul. 2012, 5, 155–162. [Google Scholar] [CrossRef]
- Nitsche, M.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef] [PubMed]
- Hays, J.; Wong, C.; Soto, F.A. FaReT: A free and open-source toolkit of three-dimensional models and software to study face perception. Behav. Res. Methods 2020, 52, 2604–2622. [Google Scholar] [CrossRef]
- O’Callaghan, C.; Hall, J.M.; Tomassini, A.; Muller, A.J.; Walpola, I.C.; Moustafa, A.A.; Shine, J.M.; Lewis, S.J. Visual hallucinations are characterized by impaired sensory evidence accumulation: Insights from hierarchical drift diffusion modeling in Parkinson’s disease. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 680–688. [Google Scholar] [CrossRef]
- Turner, B.M.; Van Maanen, L.; Forstmann, B.U. Informing cognitive abstractions through neuroimaging: The neural drift diffusion model. Psychol. Rev. 2015, 122, 312. [Google Scholar] [CrossRef] [PubMed]
- Wiecki, T.V.; Sofer, I.; Frank, M.J. HDDM: Hierarchical Bayesian estimation of the drift-diffusion model in Python. Front. Neuroinformatics 2013, 7, 14. [Google Scholar] [CrossRef]
- Zhang, J.; Rowe, J.B. Dissociable mechanisms of speed-accuracy tradeoff during visual perceptual learning are revealed by a hierarchical drift-diffusion model. Front. Neurosci. 2014, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-J.; Kim, E.-S.; Noh, B.H.; Liang, J.-G.; Lee, D.; Hur, Y.J.; Kim, N.-Y.; Kim, H.-D. Alteration in brain connectivity in patients with Dravet syndrome after vagus nerve stimulation (VNS): Exploration of its effectiveness using graph theory analysis with electroencephalography. J. Neural Eng. 2020, 17, 036014. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Lopez-Calderon, J.; Luck, S.J. ERPLAB: An open-source toolbox for the analysis of event-related potentials. Front. Hum. Neurosci. 2014, 8, 213. [Google Scholar] [CrossRef] [PubMed]
- Read, G.L.; Innis, I.J. Electroencephalography (Eeg). Int. Encycl. Commun. Res. Methods 2017, 1–18. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Zhang, B.; Wang, Y.; Zhou, X. Electrophysiological responses to expectancy violations in semantic and gambling tasks: A comparison of different EEG reference approaches. Front. Neurosci. 2018, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Sander, T.; Bock, A.; Leistner, S.; Kühn, A.; Trahms, L. Coherence and imaginary part of coherency identifies cortico-muscular and cortico-thalamic coupling. In Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology, Buenos Aires, Argentina, 31 August–4 September 2010; pp. 1714–1717. [Google Scholar] [CrossRef]
- Pavone, E.F.; Tieri, G.; Rizza, G.; Tidoni, E.; Grisoni, L.; Aglioti, S.M. Embodying others in immersive virtual reality: Electro-cortical signatures of monitoring the errors in the actions of an avatar seen from a first-person perspective. J. Neurosci. 2016, 36, 268–279. [Google Scholar] [CrossRef]
- Wang, Z.J.; Noh, B.H.; Kim, E.S.; Yang, D.; Yang, S.; Kim, N.Y.; Hur, Y.J.; Kim, H.D. Brain network analysis of interictal epileptiform discharges from ECoG to identify epileptogenic zone in pediatric patients with epilepsy and focal cortical dysplasia type II: A retrospective study. Front. Neurol. 2022, 13, 901633. [Google Scholar] [CrossRef]
- Makeig, S.; Bell, A.; Jung, T.-P.; Sejnowski, T.J. Independent component analysis of electroencephalographic data. Adv. Neural Inf. Process. Syst. 1995, 8, 145–151. [Google Scholar]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage 2019, 198, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-T.; Chung, I.-F.; Ko, L.-W.; Chen, Y.-C.; Liang, S.-F.; Duann, J.-R. EEG-based assessment of driver cognitive responses in a dynamic virtual-reality driving environment. IEEE Trans. Biomed. Eng. 2007, 54, 1349–1352. [Google Scholar] [CrossRef]
- Deng, Z.; Zhang, Z. Event-related complexity analysis and its application in the detection of facial attractiveness. Int. J. Neural Syst. 2014, 24, 1450026. [Google Scholar] [CrossRef]
- Logan, G.D.; Cowan, W.B. On the ability to inhibit thought and action: A theory of an act of control. Psychol. Rev. 1984, 91, 295. [Google Scholar] [CrossRef]
- Friedrich, J.; Beste, C. Paradoxical, causal effects of sensory gain modulation on motor inhibitory control—A tDCS, EEG-source localization study. Sci. Rep. 2018, 8, 17486. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil-Vall, L.; Gomez-Bernal, F.; Villegas, A.C.; Cirillo, P.; Surman, C.; Ruffini, G.; Widge, A.S.; Camprodon, J.A. Transcranial direct current stimulation to the left dorsolateral prefrontal cortex improves cognitive control in patients with attention-deficit/hyperactivity disorder: A randomized behavioral and neurophysiological study. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Gaspelin, N. How to get statistically significant effects in any ERP experiment (and why you shouldn’t). Psychophysiology 2017, 54, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Clayson, P.E.; Baldwin, S.A.; Larson, M.J. How does noise affect amplitude and latency measurement of event-related potentials (ERPs)? A methodological critique and simulation study. Psychophysiology 2013, 50, 174–186. [Google Scholar] [CrossRef]
- Huynh-The, T.; Pham, Q.-V.; Pham, X.-Q.; Nguyen, T.T.; Han, Z.; Kim, D.-S. Artificial intelligence for the metaverse: A survey. Eng. Appl. Artif. Intell. 2023, 117, 105581. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, H.; Geng, F.; Song, X.; Hu, Y. Internet gaming disorder increases mind-wandering in young adults. Front. Psychol. 2021, 11, 619072. [Google Scholar] [CrossRef]
- Garavan, H.; Ross, T.J.; Murphy, K.; Roche, R.A.; Stein, E.A. Dissociable executive functions in the dynamic control of behavior: Inhibition, error detection, and correction. Neuroimage 2002, 17, 1820–1829. [Google Scholar] [CrossRef]
- Aron, A.R.; Robbins, T.W.; Poldrack, R.A. Inhibition and the right inferior frontal cortex: One decade on. Trends Cogn. Sci. 2014, 18, 177–185. [Google Scholar] [CrossRef]
- Nieuwenhuis, S.; Yeung, N.; Van Den Wildenberg, W.; Ridderinkhof, K.R. Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency. Cogn. Affect. Behav. Neurosci. 2003, 3, 17–26. [Google Scholar] [CrossRef]
- Koizumi, A.; Lau, H.; Shimada, Y.; Kondo, H.M. The effects of neurochemical balance in the anterior cingulate cortex and dorsolateral prefrontal cortex on volitional control under irrelevant distraction. Conscious. Cogn. 2018, 59, 104–111. [Google Scholar] [CrossRef]
- Schulz, K.P.; Bédard, A.C.V.; Czarnecki, R.; Fan, J. Preparatory activity and connectivity in dorsal anterior cingulate cortex for cognitive control. Neuroimage 2011, 57, 242–250. [Google Scholar] [CrossRef]
- Rodrigo, A.H.; Di Domenico, S.I.; Ayaz, H.; Gulrajani, S.; Lam, J.; Ruocco, A.C. Differentiating functions of the lateral and medial prefrontal cortex in motor response inhibition. Neuroimage 2014, 85, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Chikazoe, J. Localizing performance of go/no-go tasks to prefrontal cortical subregions. Curr. Opin. Psychiatry 2010, 23, 267–272. [Google Scholar] [CrossRef]
- Liddle, P.F.; Kiehl, K.A.; Smith, A.M. Event-related fMRI study of response inhibition. Hum. Brain Mapp. 2001, 12, 100–109. [Google Scholar] [CrossRef]
- Aron, A.R.; Fletcher, P.C.; Bullmore, E.T.; Sahakian, B.J.; Robbins, T.W. Stop-signal inhibition disrupted by damage to right inferior frontal gyrus in humans. Nat. Neurosci. 2003, 6, 115–116. [Google Scholar] [CrossRef]
- Rubia, K.; Smith, A.B.; Brammer, M.J.; Taylor, E. Right inferior prefrontal cortex mediates response inhibition while mesial prefrontal cortex is responsible for error detection. Neuroimage 2003, 20, 351–358. [Google Scholar] [CrossRef]
- Lapenta, O.M.; Di Sierve, K.; de Macedo, E.C.; Fregni, F.; Boggio, P.S. Transcranial direct current stimulation modulates ERP-indexed inhibitory control and reduces food consumption. Appetite 2014, 83, 42–48. [Google Scholar] [CrossRef]
- Reinhart, R.M.; Cosman, J.D.; Fukuda, K.; Woodman, G.F. Using transcranial direct-current stimulation (tDCS) to understand cognitive processing. Atten. Percept. Psychophys. 2017, 79, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Hupfeld, K.E.; Ketcham, C.J.; Schneider, H.D. Transcranial direct current stimulation (tDCS) to Broca’s area: Persisting effects on non-verbal motor behaviors. Neurol. Disord. Ther. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Brunyé, T.T.; Holmes, A.; Cantelon, J.; Eddy, M.D.; Gardony, A.L.; Mahoney, C.R.; Taylor, H.A. Direct current brain stimulation enhances navigation efficiency in individuals with low spatial sense of direction. Neuroreport 2014, 25, 1175–1179. [Google Scholar] [CrossRef]
- Kogler, W.; Wood, G.; Kober, S.E. Effects of electrical brain stimulation on brain indices and presence experience in immersive, interactive virtual reality. Virtual Real. 2021, 26, 1019–10291. [Google Scholar] [CrossRef]
- Fox, J. Regression Diagnostics: An Introduction; Sage Publications: London, UK, 2019. [Google Scholar]
- Pamplona, G.S.; Heldner, J.; Langner, R.; Koush, Y.; Michels, L.; Ionta, S.; Scharnowski, F.; Salmon, C.E. Network-based fMRI-neurofeedback training of sustained attention. Neuroimage 2020, 221, 117194. [Google Scholar] [CrossRef] [PubMed]
- Posner, M.I.; Petersen, S.E. The attention system of the human brain. Annu. Rev. Neurosci. 1990, 13, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Posner, M.I.; Rothbart, M.K.; Voelker, P. Developing brain networks of attention. Curr. Opin. Pediatr. 2016, 28, 720. [Google Scholar] [CrossRef]
- Wu, T.; Liu, X.; Cheng, F.; Wang, S.; Li, C.; Zhou, D.; Zhang, W. Dorsolateral prefrontal cortex dysfunction caused by a go/no-go task in children with attention-deficit hyperactivity disorder: A functional near-infrared spectroscopy study. Front. Neurosci. 2023, 17, 1145485. [Google Scholar] [CrossRef]
- Mason, M.F.; Norton, M.I.; Van Horn, J.D.; Wegner, D.M.; Grafton, S.T.; Macrae, C.N. Wandering minds: The default net-work and stimulus-independent thought. Science 2007, 315, 393–395. [Google Scholar] [CrossRef]
- Keil, A.; Debener, S.; Gratton, G.; Junghöfer, M.; Kappenman, E.S.; Luck, S.J.; Luu, P.; Miller, G.A.; Yee, C.M. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology 2014, 51, 1–21. [Google Scholar] [CrossRef]
- Wessel, J.R.; Aron, A.R. It’s not too late: The onset of the frontocentral P 3 indexes successful response inhibition in the stop-signal paradigm. Psychophysiology 2015, 52, 472–480. [Google Scholar] [CrossRef]
- Huster, R.J.; Enriquez-Geppert, S.; Lavallee, C.F.; Falkenstein, M.; Herrmann, C.S. Electroencephalography of response inhibition tasks: Functional networks and cognitive contributions. Int. J. Psychophysiol. 2013, 87, 217–233. [Google Scholar] [CrossRef]
- Osimo, S.A.; Piretti, L.; Ionta, S.; Rumiati, R.I.; Aiello, M. The neural substrates of subliminal attentional bias and reduced inhibition in individuals with a higher BMI: A VBM and resting state connectivity study. NeuroImage 2021, 229, 117725. [Google Scholar] [CrossRef]
- Silva, A.F.; Zortea, M.; Carvalho, S.; Leite, J.; Torres, I.L.D.S.; Fregni, F.; Caumo, W. Anodal transcranial direct current stimulation over the left dorsolateral prefrontal cortex modulates attention and pain in fibromyalgia: Randomized clinical trial. Sci. Rep. 2017, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Khoshnoud, S.; Shamsi, M.; Nazari, M.A.; Makeig, S. Different cortical source activation patterns in children with attention deficit hyperactivity disorder during a time reproduction task. J. Clin. Exp. Neuropsychol. 2018, 40, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Bhavnani, S.; Lockwood Estrin, G.; Haartsen, R.; Jensen, S.K.; Gliga, T.; Patel, V.; Johnson, M.H. EEG signatures of cognitive and social development of preschool children—A systematic review. PLoS ONE 2021, 16, e0247223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Tian, F.; Wu, X.; Liao, S.; Qiu, J. The neural correlates of insight in Chinese verbal problems: An event related-potential study. Brain Res. Bull. 2011, 84, 210–214. [Google Scholar] [CrossRef]
- Lisanby, S.H.; Luber, B.; Schlaepfer, T.E.; Sackeim, H.A. Safety and feasibility of magnetic seizure therapy (MST) in major depression: Randomized within-subject comparison with electroconvulsive therapy. Neuropsychopharmacology 2003, 28, 1852–1865. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Model 1 (Stimulation Type, P300 in No-Go, P300 in Go) | Model 2 (P400 in No-Go Added) | ||
|---|---|---|---|---|
| Adjusted R2 | Regression Prediction Equation | Variation of Adjusted R2 | Regression Prediction Equation | |
| Mental state | 0.755 ** | y = 67 + 25.43x1 − 16.29x2 + 17x3 ** | −0.011 | y = 69.52 + 30.04x1 − 29.1x2 + 21.34x3 + 16.06x4 ** |
| Mental score | 0.569 ** | y = 3.68 − 0.16x1 − 0.26x2 + 0.8x3 ** | −0.026 | y = 3.7 − 0.1x1 − 0.41x2 + 0.86x3 + 0.19x4 * |
| Accuracy | 0.825 ** | y = 0.84 + 0.05x1 − 0.05x2 + 0.09x3 ** | −0.011 | y = 0.84 + 0.05x1 − 0.05x2 + 0.09x3 + 0.01x4 ** |
| RT | 0.255 | y = 0.54 + 0.15x1 − 0.01x2 − 0.11x3 | 0.049 | y = 0.54 + 0.15x1 − 0.01x2 − 0.11x3 − 0.01x4 |
| vgo | 0.520 * | y = 8.69 + 2.95x1 − 3.68x2 + 2.47x3 * | 0.026 | y = 9.34 + 4.16x1 − 7.04x2 + 3.61x3 + 4.21x4 * |
| vnogo | 0.865 ** | y = −1.81 − 1.61x1 + 0.73x2 − 0.41x3 ** | −0.009 | y = −1.83 − 1.66x1 + 0.86x2 − 0.46x3 − 0.16x4 ** |
| NDT | 0.134 | y = 0.45 + 0.23x1 − 0.14x2 − 0.11x3 | 0.110 | y = 0.48 + 0.28x1 − 0.26x2 − 0.07x3 + 0.16x4 |
| a | 0.918 ** | y = 1.74 + 0.44x1 + 0.02x2 + 0.29x3 ** | 0.001 | y = 1.66 + 0.29x1 + 0.43x2 + 0.15x3 − 0.52x4 ** |
| z | 1.00 ** | y = 0.23 − 0.06x1 + 0.01x2 − 0.01x3 ** | 0 | y = 0.23 − 0.06x1 + 0.01x2 − 0.01x3 − 0.01x4 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Enkhzaya, G.; Zhu, B.-H.; Chen, J.; Wang, Z.-J.; Kim, E.-S.; Kim, N.-Y. High-Definition Transcranial Direct Current Stimulation in the Right Ventrolateral Prefrontal Cortex Lengthens Sustained Attention in Virtual Reality. Bioengineering 2023, 10, 721. https://doi.org/10.3390/bioengineering10060721
Yang S, Enkhzaya G, Zhu B-H, Chen J, Wang Z-J, Kim E-S, Kim N-Y. High-Definition Transcranial Direct Current Stimulation in the Right Ventrolateral Prefrontal Cortex Lengthens Sustained Attention in Virtual Reality. Bioengineering. 2023; 10(6):721. https://doi.org/10.3390/bioengineering10060721
Chicago/Turabian StyleYang, Shan, Ganbold Enkhzaya, Bao-Hua Zhu, Jian Chen, Zhi-Ji Wang, Eun-Seong Kim, and Nam-Young Kim. 2023. "High-Definition Transcranial Direct Current Stimulation in the Right Ventrolateral Prefrontal Cortex Lengthens Sustained Attention in Virtual Reality" Bioengineering 10, no. 6: 721. https://doi.org/10.3390/bioengineering10060721
APA StyleYang, S., Enkhzaya, G., Zhu, B.-H., Chen, J., Wang, Z.-J., Kim, E.-S., & Kim, N.-Y. (2023). High-Definition Transcranial Direct Current Stimulation in the Right Ventrolateral Prefrontal Cortex Lengthens Sustained Attention in Virtual Reality. Bioengineering, 10(6), 721. https://doi.org/10.3390/bioengineering10060721