1. Introduction
Oral inflammatory disease is a general term for the inflammatory lesions developed in oral mucosa. The pathogenesis of oral inflammatory diseases is non-uniform due to the involvement of various factors—such as external and mechanical stimuli, the presence of microorganisms, and the overall physical conditions—that play a role in the onset of inflammation. There is a wide range of variations in the aspect of oral inflammatory diseases, and the aspect is unequal. Therefore, we construed the oral inflammatory diseases as a symptom of inflammation, and categorized them into acute and chronic inflammatory diseases. In the oral region, the representative example of acute inflammatory diseases is stomatitis (also named as oral mucositis), and that of chronic inflammatory diseases is periodontitis. Several Japanese herbal medicines (also known as kampo medicines) are clinically used for the treatment of inflammatory diseases. Recent reviews have summarized the therapeutic application of herbal medicines for oral diseases such as stomatitis and periodontitis [
1]. For example, hangeshashinto (TJ-14) is used for inflammatory diseases such as acute or chronic gastrointestinal catarrh, nervous gastritis and stomatitis.
In this review, we aim to summarize the biological efficacy of herbal medicine, natural products, and their active ingredients against acute and chronic inflammatory diseases in the oral region, especially stomatitis and periodontitis.
3. Biological Efficacy of Natural Products against Chronic Inflammatory Disease; Periodontitis
3.1. Periodontitis
Periodontal disease (periodontitis) comprises a group of infections that leads to inflammation of the gingiva and destruction of periodontal tissues, and is accompanied by alveolar bone loss in severe clinical cases. The tissue destruction is the result of activation of the host’s immuno-inflammatory response to virulent factors such as LPS and peptidoglycan. In inflammatory responses and tissue degradation, prostaglandin E
(PGE
), interleukin (IL)-6, and IL-8 play important roles. As PGE
has several functions in vasodilation, the enhancement of vascular permeability and pain, and osteoclastogenesis induction, PGE
participates in inflammatory responses and alveolar bone resorption in periodontitis [
24].
Generally, periodontitis is a chronic inflammation, and the elimination of these virulent factors by initial preparation is very important for the treatment of periodontitis. However, during the acute advanced stage, non-steroidal anti-inflammatory drugs (NSAIDs) are administrated to improve gingival inflammation. In fact, many studies have demonstrated that systemic administration of acid NSAIDs prevented gingival inflammation and alveolar bone resorption in animals and humans [
25]. However, acid NSAIDs are well known to have side effects such as gastrointestinal dysfunction and bronchoconstriction. Therefore, the usage of alternative agents is necessary for patients with gastrointestinal ulcer or bronchial asthma. Previously, we suggested that several herbal medicines are effective for the improvement of periodontitis. In this review, we focused on the anti-inflammatory effects of herbal medicines on mainly periodontitis —in particular, about the effects on human gingival fibroblasts (HGFs). In addition, we summarized the effects of ingredients in herbs and their mechanism against arachidonic acid cascade.
Here, we will explain the importance of HGFs in the study of periodontitis. (1) HGFs are the most prominent cells in periodontal tissue. LPS-treated HGFs produce inflammatory chemical mediators, such as PGE
, and inflammatory cytokines such as IL-6 and IL-8. (2) More importantly, unlike macrophages, HGFs continue to produce PGE
[
26], IL-6, and IL-8 [
27] in the presence of LPS. From these findings, the large amount of chemical mediators and cytokines derived from HGFs may be contained in periodontal tissues. Therefore, we believe that examining the effects of pharmaceuticals on HGFs is needed in the study of periodontitis.
3.2. Brief Summary of Arachidonic Acid Cascade
At first, we explain arachidonic acid cascade briefly, focusing on sites of action for herbs and ingredients. PGE
is produced by arachidonic acid cascade (
Figure 2). Phospholipids in plasma membrane are digested by phospholipase A
(PLA
), producing arachidonic acid. Cyclooxygenases (COXs) convert arachidonic acid into PGH
, and thereafter PGE synthase converts into PGE
.
PLA
is the most upstream enzyme in the arachidonic acid cascade and releases arachidonic acid from the plasma membrane. PLA
forms a superfamily and is classified into cytosolic PLA
(cPLA
), calcium-independent PLA
(iPLA
), secretory PLA
(sPLA
), and others [
28]. Among these isoforms, cPLA
is the primary isoform in HGFs from the results using PLA
inhibitors [
29]. cPLA
activity is directly regulated by extracellular signal-regulated kinase (ERK). The active form of ERK (phosphorylated ERK) phosphorylates Ser505 of cPLA
and activates cPLA
[
30,
31,
32]. Therefore, the suppression of ERK phosphorylation leads to the suppression of cPLA
activation and the reduction of PGE
production [
30,
31,
32]. In contrast, annexin1, also named as lipocortin, is an anti-inflammatory mediator produced by steroidal anti-inflammatory drugs (SAIDs) that inhibits PLA
activity [
33,
34].
COX is classified into COX-1 and COX-2. COX-1 is constitutive expressed at low level, and is involved in normal functions such as protection of gastric mucosa. In contrast, COX-2 is induced by the various stimuli such as LPS and peptidoglycan, and involved in inflammatory response. The expression of COX-2 is upregulated by NF-B. The reduction of PGE by anti-inflammatory drugs is one of the important mechanisms. Acid NSAIDs inhibit both COX-1 and COX-2 activities. The inhibition of COX-2 improve inflammatory response, while the inhibition of COX-1 causes gastric irritation. SAIDs also have powerful anti-inflammatory effects, and inhibit NF-B activity and suppress COX-2 expression.
Recently, protein kinase A (PKA) pathway is reported to regulate LPS-induce PGE
production in HGFs [
35]. PKA inhibitor (H-89) reduced LPS-induced PGE
production in a concentration-dependent manner. In contrast, PKA activator (dibutyryl cAMP; dbcAMP) and drugs which increased intracellular cAMP (adrenaline and aminophylline) increased LPS-induced PGE
production in a concentration-dependent manner. However, the effects of PKA pathway on arachidonic acid cascade have not been examined in this report [
35].
3.3. Effect of Herbal Medicines on Periosontal Disease
Similar to NSAIDs, several herbal medicines also reduce PGE
production. Examples of herbal medicine which have been reported to reduce PGE
production in in vitro and/or animal models are shown in
Table 1. In particular, we reported that kakkonto (TJ-1), shosaikoto (TJ-9), hangeshashinto (TJ-14), shinbuto (TJ-30), ninjinto (TJ-32), and orento (TJ-120) reduced LPS-induced PGE
production using HGFs [
36,
37,
38,
39,
40]. Other groups have also demonstrated that several herbal medicines reduced PGE
production using human periodontal ligament cells [
41], human monocytes [
42], mouse macrophage RAW264.7 cells [
43,
44], human oral keratinocytes [
7], and animals [
45,
46,
47,
48].
We introduce briefly the effects and mechanisms of herbal medicines on periodontitis in clinical, animal, and/or in vitro studies. Moreover, in this section, we will demonstrate the effects of herbal medicines on the reduction of PGE in HGFs. From our data, the mechanisms of these herbal medicines on arachidonic acid cascade are divided into three groups as follows.
Shosaikoto (TJ-9) inhibited COX-2 activity and suppressed COX-2 expression, but did not alter cPLA
expression (the effects on annexin1 expression and ERK phosphorylation were not examined) [
37]. Hangeshashinto (TJ-14) inhibited both COX-1 and COX-2 activities, and suppressed cPLA
and COX-2 expressions and ERK phosphorylation [
38]. Therefore, these herbal medicines are suggested to inhibit arachidonic acid cascade at multiple points.
Shinbuto (TJ-30) and ninjinto (TJ-32) enhanced annexin1 expression, but did not alter ERK phosphorylation and COX activity [
39]. However, the contribution of enhancement of annexin1 expression is considered to be small because shokyo, which is the main herb in shinbuto to reduce PGE
production, did not affect annexin1 expression.
Kakkonto (TJ-1) suppressed ERK phosphorylation, but neither inhibited COXs activities nor suppressed the expression of molecules in arachidonic acid cascade [
36]. In addition, orento (TJ-120) suppressed ERK phosphorylation, but neither inhibited COXs activities nor suppressed the expression of molecules in arachidonic acid cascade, but rather increased COX-2 expression [
40]. However, its contribution in the suppression of ERK phosphorylation is considered to be small as described at keihi (
Cinnamomi Cortex). Indeed, we did not examine the direct effect of herbal medicines on cPLA
activity. Nevertheless, we consider that these herbal medicines inhibit cPLA
activity and that this effect is due to shokyo (
Zingiberis Rhizoma) and kankyo (
Zingiberis Processum Rhizoma) as described below.
3.4. Effect of Herbs on Arachidonic Acid Cascade
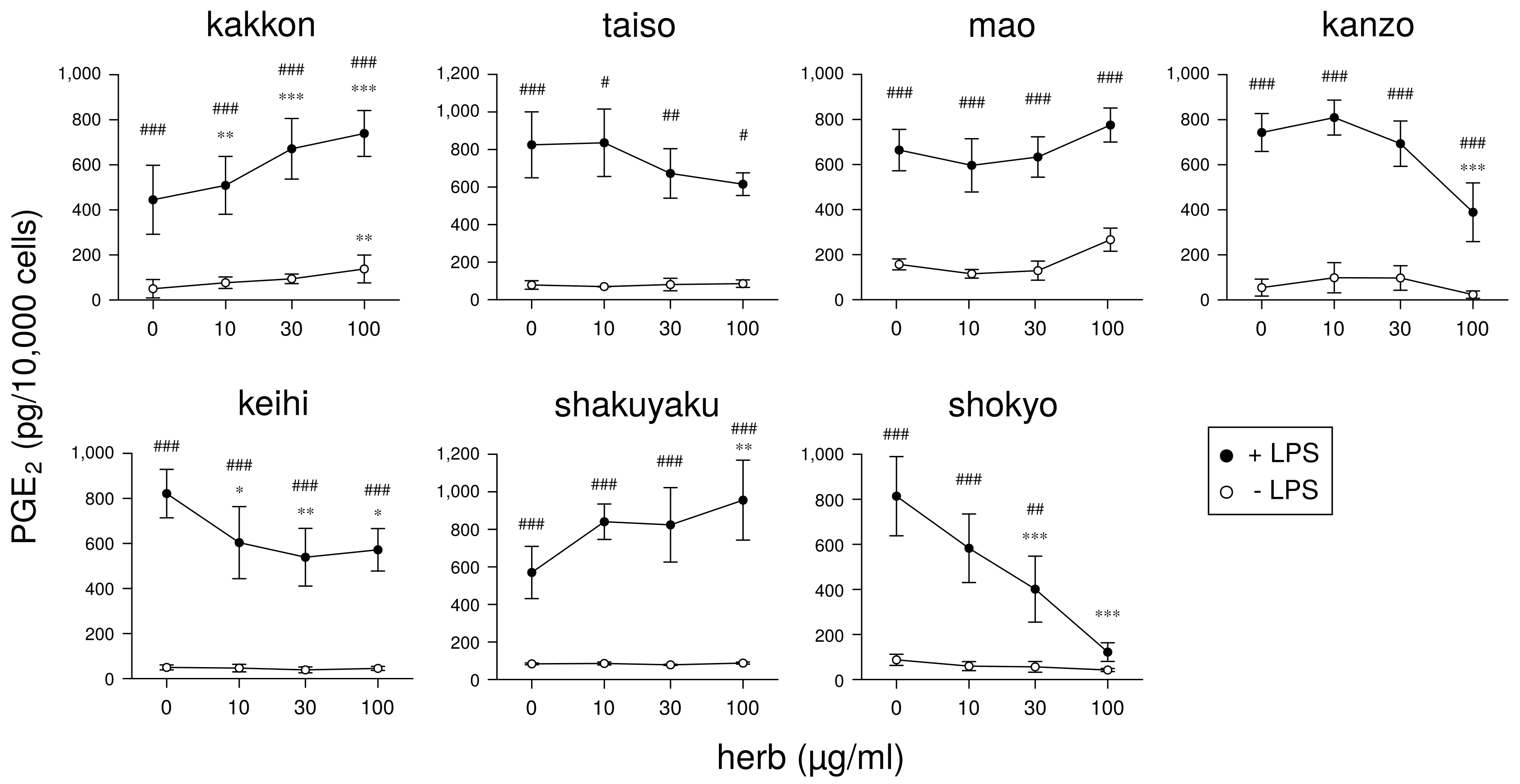
Next, we will demonstrate the experimental results at the herb level. The ingredients in the formula of herbal medicines that were used are shown in
Table 2,
Table 3,
Table 4,
Table 5,
Table 6 and
Table 7. In our experiments at the herb level, shokyo (
Zingiberis Rhizoma), kankyo (
Zingiberis Processum Rhizoma), kanzo (
Glycyrrhizae Radix), and keihi (
Cinnamomi Cortex) reduced PGE
production (
Figure 3 and
Figure 4) [
29,
39]. We summarized major ingredients in herbs and their mechanism against arachidonic acid cascade in
Table 8. In addition to these four herbs, ogon (
Scutellariae Radix), and oren (
Coptidis Rhizoma) are shown in
Table 8 because ogon (included in shosaikoto and hangeshashinto) and oren (included in hangeshashinto and orento) also have several bioactive ingredients such as flavonoids, saponin, and chalcones. We will describe the effects and mechanisms of these herbs, particularly shokyo and kankyo, on arachidonic cascade.
3.4.1. Shokyo (Zingiberis Rhizoma)/Kankyo (Zingiberis Processum Rhizoma)
Shokyo is the powdered rhizome of
Zingiber offinale Roscoe (ginger), and kankyo is the steamed and powdered rhizome of ginger. Both shokyo and kankyo are the aqueous extracts of ginger. Among the herbal medicines shown in
Table 1, shokyo is included in kakkonto (TJ-1), shosaikoto (TJ-9), shinbuto (TJ-30), saireito (TJ-114), and orento (TJ-120), and kankyo is included in hangeshashinto (TJ-14) and ninjinto (TJ-32). Many reports have demonstrated that ginger possesses anti-inflammatory effects in human [
80,
81] and animal models [
82,
83,
84], and in vitro [
85]. Ginger has been widely used in diet and as a treatment for rheumatoid arthritis, fever, emesis, nausea, and migraine headache [
80]. A recent systematic review shows that the extracts of ginger including turmeric, ginger, Javanese ginger, and galangal are clinically effective as hypoanalgesic agents [
81]. In an animal model, the aqueous extract of ginger significantly reduced serum PGE
level by oral or intraperitoneal administration in rats [
82]. Moreover, crude hydroalcoholic extract of ginger reduced the serum level of PGE
, and improved tracheal hyperreactivity and lung inflammation induced by LPS in rats [
83]. Ethanol extract of ginger reduced the tissue level of PGE
, and improved acetic acid-induced ulcerative colitis in rats [
84].
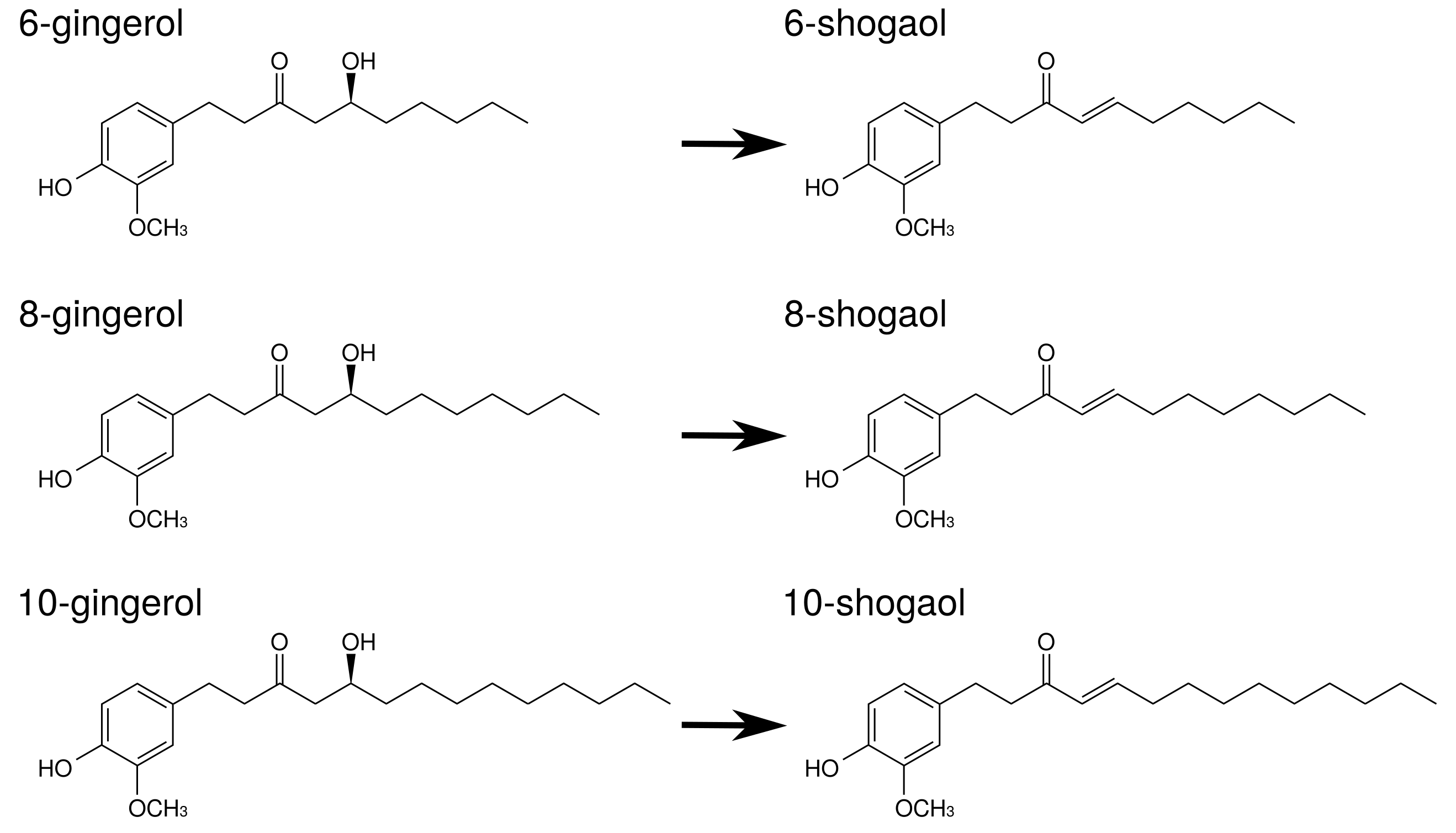
Gingerols and shogaols are the major ingredients in ginger. Their structures are indicated in
Figure 5. With prolonged storage or heat-treatment of ginger, gingerols are converted to shogaols, which are the dehydrated form of the gingerols [
80] (
Figure 5). Therefore, kankyo contains a larger amount of shogaols than shokyo although both shokyo and kankyo contain gingerols and shogaols. In in vitro models, gingerols and shogaols have been reported to reduce PGE
production by several mechanisms. The effects of gingerols and shogaols on arachidonic acid cascade are briefly summarized in
Table 8.
Gingerols and shogaols inhibit COX-2 activity. Their IC
values were
M order: 6-gingerol (>50
M), 8-gingerol (10.0
M), 10-gingerol (3.7
M), 6-shogaol (2.1
M), and 8-shogaol (7.2
M) in human lung adenocarcinoma A549 cells [
49], and 10-gingerol (32.0
M), 8-shogaol (17.5
M), 10-shogaol (7.5
M) in a cell-free assay [
50].
Gingerols and shogaols suppress COX-2 expression. For example, 6-, 8-, and 10-gingerol suppressed COX-2 expression in LPS-treated human leukemic monocyte lymphoma U937 cells [
51]. Similarly, 6-gingerol and 6-shogaol suppressed LPS-induced COX-2 expression in mouse macrophage RAW264.7 cells [
52], mouse microglial BV-2 cells [
53], and primary rat astrocytes [
86]. 6-Gingerol suppressed COX-2 expression in TPA-treated mouse skin in vivo [
54].
As aforementioned, the expression of COX-2 is regulated by NF-
B. Gingerols and shogaols are reported to suppress NF-
B activation, and to downregulate COX-2 expression. For example, 6-shogaol suppressed LPS-induced NF-
B activation in RAW264.7 cells [
52], mouse primary cultured microglia cells [
53], and human breast cancer MDA-MB-231 cells [
56]. 6-Shogaol suppressed TPA-induced NF-
B activation in mouse skin [
54]. Similarly, 6-gingerol suppressed
Vibrio cholerae-induced NF-
B activation in human intestinal epithelial cells [
55]. These results suggest that gingerols and shogaols suppress NF-
B activation directly or indirectly, leading to the inhibition of COX-2 expression.
Gingerols and shogaols inhibit PLA
activities [
57]. In more detail, iPLA
activity was inhibited by 6-, 8-, and 10-gingerol and 6-, 8-, and 10-shogaol, whereas cPLA
activity was inhibited by 6-gingerol and 6-, 8-, and 10-shogaol. In particular, IC
values of 10-shogaol against iPLA
and cPLA
were 0.7
M and 3
M, respectively, in U937 cells.
As aforementioned, many reports have examined the effects of ginger. However, there is little report using ginger as “shokyo” and “kankyo.” For this reason, we examined the mechanism of the actions of shokyo and kankyo on the reduction of PGE
production in HGFs. Shokyo and kankyo concentration-dependently reduced LPS-induced PGE
production by HGFs, and the effects of kankyo were slightly stronger than those of shokyo (
Figure 4) [
39]. The effects of shokyo and kankyo on arachidonic cascade in HGFs are described as follows.
Both shokyo and kankyo only slightly increased cPLA
expression, and did not alter annexin1 expression [
39].
Shokyo did not alter LPS-induced ERK phosphorylation in HGFs [
29] (but we have not examined the effect of kankyo). Therefore, shokyo (and perhaps kankyo) may have little to no effect on cPLA
activation, and the subsequent arachidonic acid production.
Both shokyo and kankyo did not inhibit COX-2 and PGE synthase activities, and did not alter LPS-induced COX-2 expression in HGFs [
29,
39]. These findings suggest shokyo and kankyo primarily inhibit cPLA
activity in HGFs. Although we have no direct data to show that shokyo and kankyo inhibit cPLA
activity, this assumption is consistent with the results that ginger (and gingerols/shogaols) inhibits both iPLA
and cPLA
activities [
57].
As described above, our data that shokyo did not alter COX-2 activity and COX-2 expression are different from those of gingerols and shogaols in
Table 8. Although there is no obvious evidence, the reason may be the preparation method of shokyo and kankyo. Gingerols and shogaols are extremely hydrophobic by their structures. These ingredients were extracted from hydrophobic phase, whereas shokyo and kankyo were prepared by decoction. Therefore, hydrophobic ingredients such as gingerol and shogaol are unlikely to be extracted, and their concentration might be lower than those in previous reports. Quantification of these ingredients is needed to explain these discrepancies.
3.4.2. Kanzo (Glycyrrhizae Radix)
Kanzo is the powdered root or stolon of
Glycyrrhiza uralensis Fischer (licorice). Among the herbal medicines shown in
Table 1, kanzo is included in kakkonto (TJ-1), shosaikoto (TJ-9), hangeshashinto (TJ-14), ninjinto (TJ-32), rikkosan (TJ-110), saireito (TJ-114), and orento (TJ-120). Licorice is also known to have anti-inflammatory effects [
87] such as inhibition of COX-2 activity [
46].
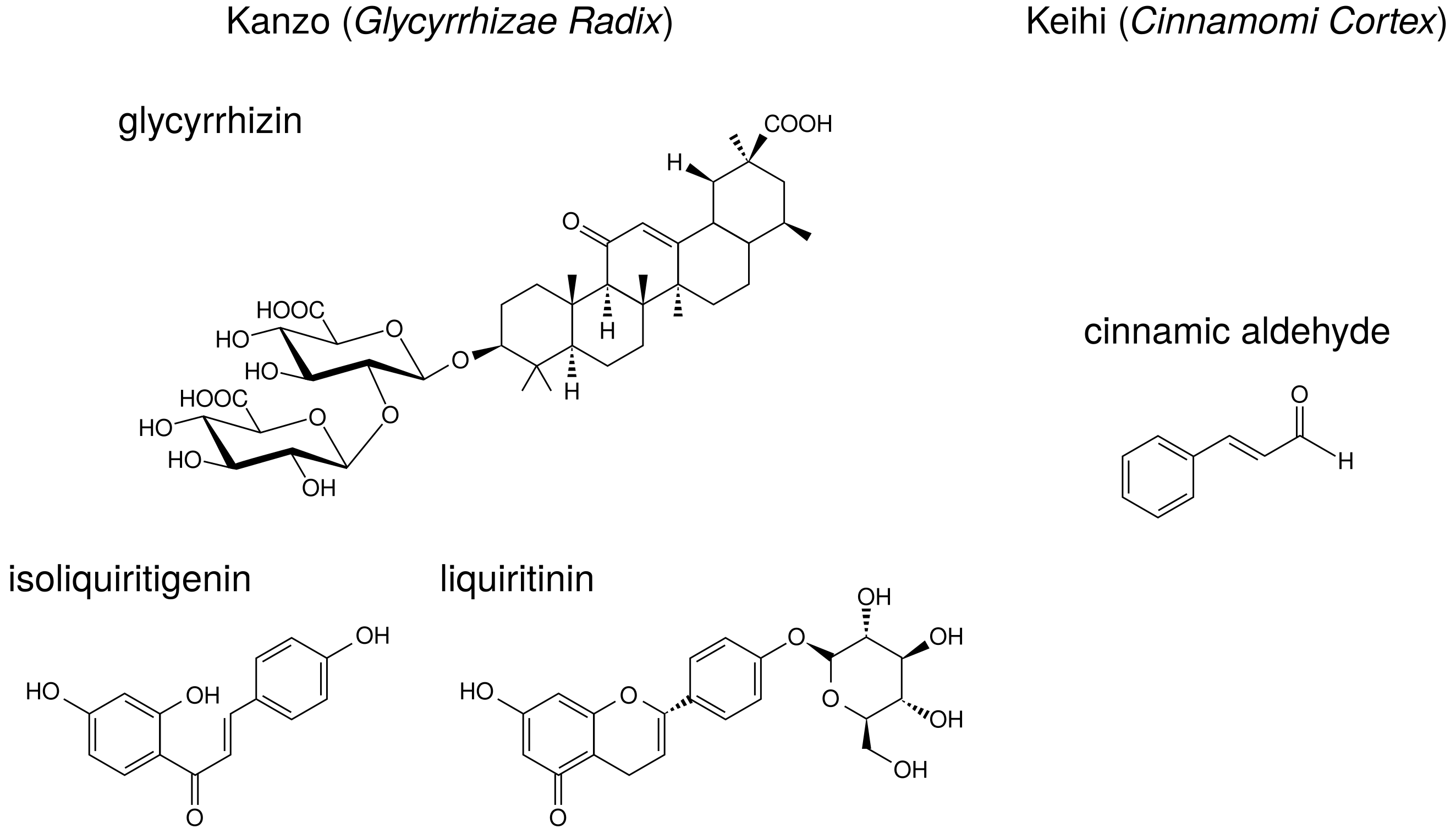
Licorice contains triperpene saponin such as glycyrrhizin (glycyrrhizin acid), and chalcones such as liquiritin and isoliquiritigenin. Their structures are indicated in
Figure 6. Glycyrrhizin, liquiritin, and isoliquiritigenin are reported to reduce PGE
production. The effects of these ingredients on arachidonic acid cascade are briefly summarized in
Table 8.
Glycyrrhizin suppressed COX-2 expression in LPS-treated mouse microglial BV2 cells [
58] and uterus of ovariectominezed mice [
59]. Moreover, orally administrated glycyrrhizin suppressed COX-2 expression in the cerebral cortex of LPS-treated mice [
60]. Liquiritin and isoliquiritigenin also suppressed LPS-induced COX-2 expression in RAW264.7 cells [
63] and BV2 cells [
58].
Glycyrrhizin suppressed TNF-
or IL-1
-induced NF-
B activation in human lung epithelial A549 cells [
61]. Isoliquiritigenin also suppressed NF-
B activity and suppressed LPS-induced COX-2 expression in RAW264.7 cells [
64].
Glycyrrhizin and isoliquiritigenin inhibited TLR4 (receptor of LPS) homodimerization and downstream signal pathway [
62], resulting in the suppression of COX-2 expression.
Indeed, although glycyrrhizin has anti-inflammatory effects, glycyrrhizin is known to show a serious adverse effect, pseudohyperaldosteronism. Excessive dietary intake of licorice can cause a syndrome mimicking hypermineralocorticoidism, characterized by hypertension, hypokalemia, alkalosis, and reduced plasma renin [
88,
89,
90,
91]. Glycyrrhizin inhibits 11
-hydroxysteroid dehydrogenase type 2 (11
-HSD2), which converts active glucocorticoid cortisol to inactive cortisone [
92]. This inhibition results in the activation of renal mineralocorticoid receptors by cortisol, inducing Na
reabsorption, K
excretion, hypertension, hypokalemia, and metabolic alkalosis. These phenotypes are similar to that of apparent mineralocorticoid excess syndrome. [
91,
93].
We examined the mechanism of the action of kanzo on the reduction of PGE production in HGFs. However, the effects of kanzo on arachidonic acid cascade in HGFs cannot be explained by those of glycyrrhizin, liquiritin, and isoliquiritigenin.
As reported previously [
46], kanzo inhibited COX-2 activity because kanzo decreased LPS-induced PGE
production when arachidonic acid was added [
29]. In contrast, kanzo did not inhibit PGE synthase activity because kanzo did not alter LPS-induced PGE
production when PGH
was added [
29].
Kanzo increased both cPLA
and annexin1 expressions [
29], thus leaving the effect of kanzo on PLA
unconcluded.
Kanzo increased LPS-induced COX-2 expression [
29] although glycyrrhizin, liquiritin, and isoliquiritigenin suppressed COX-2 expression [
58,
59,
60,
63,
64].
This result is the same as those observed using orento [
40] and saireito [
44], which contain kanzo.
Therefore, these effects of kanzo were different from those of glycyrrhizin, liquiritin, and isoliquiritigenin, suggesting that other ingredients may contribute to our findings. In addition, not all herbal medicines which contain kanzo increased annexin1 as kakkonto, hangeshashinto, and orento did not alter annexin1 expression.
3.4.3. Keihi (Cinnamomi Cortex)
Keihi is the powdered bark of
Cinnamomum cassia (cinnamon). Among the herbal medicines shown in
Table 1, keihi is included in kakkonto (TJ-1), saireito (TJ-114), and orento (TJ-120). Cinnamon has been widely used for the treatment of fever and inflammation [
28]. Cinnamon improves nephritis, purulent dermatitis, and hypertension, and it also enhances wound healing. Cinnamon extracts have been used for the improvement or prevention of common cold, diarrhea, and pain [
28]. Ethanol-extract of
C. cassia reduced LPS-induced PGE
production by RAW264.7 cells, and it suppressed NF-
B activity and the following COX-2 expression [
66].
Keihi contains the ingredients such as cinnamic aldehyde, cinnamic alcohol, cinnamic acid, and coumarin. The structure of cinnamic aldehyde is indicated in
Figure 6. Cinnamic aldehyde is reported to reduce PGE
production. The effects of cinnamic aldehyde on arachidonic acid cascade are briefly summarized in
Table 8.
Cinnamic aldehyde suppressed carrageenan-induced COX-2 expression and improved footpad edema in mice [
65]. Cinnamic aldehyde, but not others, suppressed LPS-induced COX-2 expression and decreased PGE
production by RAW264.7 cells [
65,
66].
Cinnamic aldehyde suppressed LPS-induced NF-
B activity in RAW264.7 cells and TLR4- expressing HEK293 cells [
67].
Cinnamic aldehyde inhibited IL-1
-induced COX-2 activity in rat cerebral microvascular endothelial cells although its effect is weak [
68].
Cinnamic aldehyde inhibited TLR4 oligomerization and downstream signal pathway, which include NF-
B. Sulfhydryl modification is suggested to be an important contributing factor for the regulation of TLR4 activation [
69].
We examined the mechanism of action of keihi on the reduction of PGE production in HGFs. However, the effects of keihi on arachidonic acid cascade in HGFs cannot be explained by that of cinnamic aldehyde.
Therefore, these effects of keihi are different from that of cinnamic aldehyde, suggesting that other ingredients may contribute to our findings.
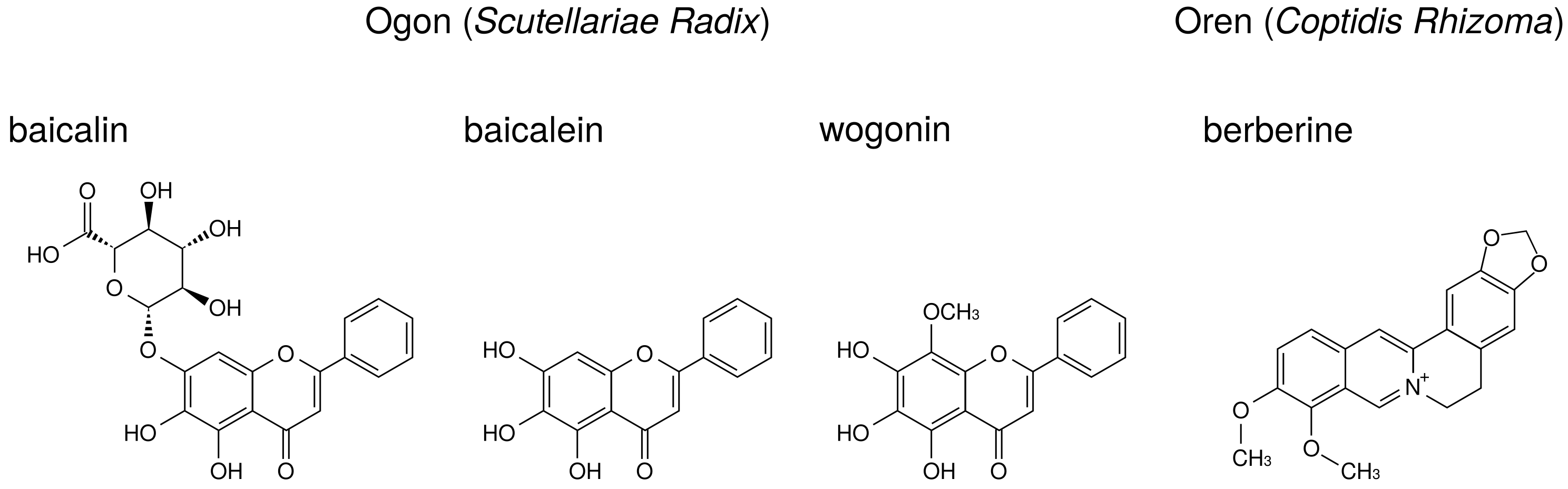
3.4.4. Ogon (Scutellariae Radix)
Ogon is the powdered root of
Scutellaria baicalensis Georgi. Among the herbal medicines shown in
Table 1, ogon is included in shosaikoto (TJ-9), hangeshashinto (TJ-14), and saireito (TJ-114). Among the herbs constituting saireito, ogon is reported to reduce PGE
production by LPS-treated RAW264.7 cells [
44].
The major ingredients of ogon are flavonoids such as baicalin, baicalein, and wogonin. Their structures are indicated in
Figure 7. Baicalin is the glucuronide of baicalein and is an inactive form. Administered baicalein is metabolized to baicalin, which is an active form. Baicalin, baicalein, and wogonin reduce PGE
production in human oral keratinocytes [
7] and RAW264.7 cells [
94].
Wogonin suppressed LPS-induced COX-2 expression in RAW264.7 cells [
73,
74], whereas baicalin and baicalein did not [
73]. Other group demonstrated that baicalein (but not baicalin) suppressed LPS-induced COX-2 expression in RAW264.7 cells [
71]. This discrepancy may be due to the concentrations of LPS and flavonoids among these reports. Moreover, baicalein and wogonin suppressed COX-2 expression in human oral keratinocytes [
7].
Baicalin [
70], baicalein [
72], and wogonin [
7] suppressed NF-
B activity.
Baicalin, baicalein, and wogonin did not inhibit COX-2 activity in RAW264.7 cells [
73].
Our data indicate that shosaikoto and hangeshashinto, which include ogon, suppressed LPS- induced COX-2 expression in HGFs [
37,
38]. This mechanism is accounted for by those of baicalin, baicalein, and wogonin.
3.4.5. Oren (Coptidis Rhizoma)
Oren is the powdered rhizome of
Coptis japonica Makino,
Coptis chinensis Franchet,
Coptis deltoidea C. Y. Cheng et Hsiao, or
Coptis teeta Wallich (Ranunculaceae). Among the herbal medicines shown in
Table 1, oren is included in hangeshashinto (TJ-14) and orento (TJ-120).
Berberine, one of benzylisoquinoline alkaloid, is the major ingredient of oren. The structure of berberine is indicated in
Figure 7. Berberine is reported to reduce PGE
production. The effects of berberine on arachidonic acid cascade are briefly summarized in
Table 8.
Berberine suppressed NF-
B activation and COX-2 expression in human leukemia Jurkat cells [
75] and oral cancer OC2 and KB cells [
95,
96].
Berberine suppressed MAP kinases phosphorylation (including ERK) and activated AMP-activated protein kinase (AMPK) in peritoneal macrophages and RAW 264.7 cells [
76], BV-2 cells [
77], and melanoma cells [
78]. Therefore, berberine is considered to inhibit cPLA
activation through suppression of ERK phosphorylation. In addition, because AMPK is reported to suppress NF-
B activation [
97], berberine suppressed COX-2 expression due to activation of AMPK.
3.5. Conclusion about Herbal Medicines and Herbs
We have described the effects of herbal medicines, herbs, and their ingredients on arachidonic acid cascade in this review. Several herbal medicines show reduced LPS-induced PGE production by HGFs. These results suggest that these herbal medicines may be effective in the improvement of the inflammatory symptoms in periodontitis. Herbal medicines must be properly selected by the patterns of each patient —excess patterns, medium patterns, or deficiency patterns. Among the herbal medicines in our studies, kakkonto (TJ-1) and orento (TJ-120) are used for the patients with excess patterns. Shosaikoto (TJ-9), hangeshashinto (TJ-14), and orento are used for the patients with medium patterns. Shinbuto (TJ-30) and ninjinto (TJ-32) are used for the patients with deficiency patterns. Therefore, it may be possible to use appropriate herbal medicines to patients with any pattern.
As shown in the above-mentioned descriptions, not all effects of herbal medicines are explainable by the effects of herbs constituting herbal medicines. Similarly, not all effects of herbs are explainable by the effects of ingredients contained in herbs. Experiments using “herbal medicines” or “herbs” themselves may be important rather than those using ingredients. The concentrations of these hydrophobic ingredients may also be low because the herbs that we used are water-soluble fractions. Therefore, it is considered that the concentrations of their ingredients need to be measured. Moreover, the unanalyzed ingredients other than those explained in this review are likely to be present. It is to be desired that further analyses reveal the novel ingredients and their action of mechanisms.
4. Anti-Osteoclastogenic Effects of Natural Products
Like periodontitis (PD), rheumatoid arthritis (RA) is a disease associated with inflammation and bone destruction. Although therapeutics of RA have recently advanced with the development of antibody drugs, natural substances displaying anti-inflammatory and anti-osteoclast characteristics against RA are still being used as widely as they have been in the past.
Some studies have revealed a relationship between PD and RA. RA prevalence is increased in patients with PD [
98,
99]. The presence of PD may contribute to the progression of RA; that is, RA patients with PD receiving non-surgical periodontal treatment resulted in a noteworthy improvement in the clinical outcome for RA [
100]. From the aspect of the clinical marker, RA and PD are similar in cytokines and mediators involved in inflammation and bone destruction [
101]. For example, TNF-
, receptor activator of nuclear factor-
B ligand (RANKL), and matrix metalloproteinase (MMP) family increase in production in RA and PD [
102,
103,
104,
105,
106]. Due to these similarities, natural products used for RA are probably effective for PD.
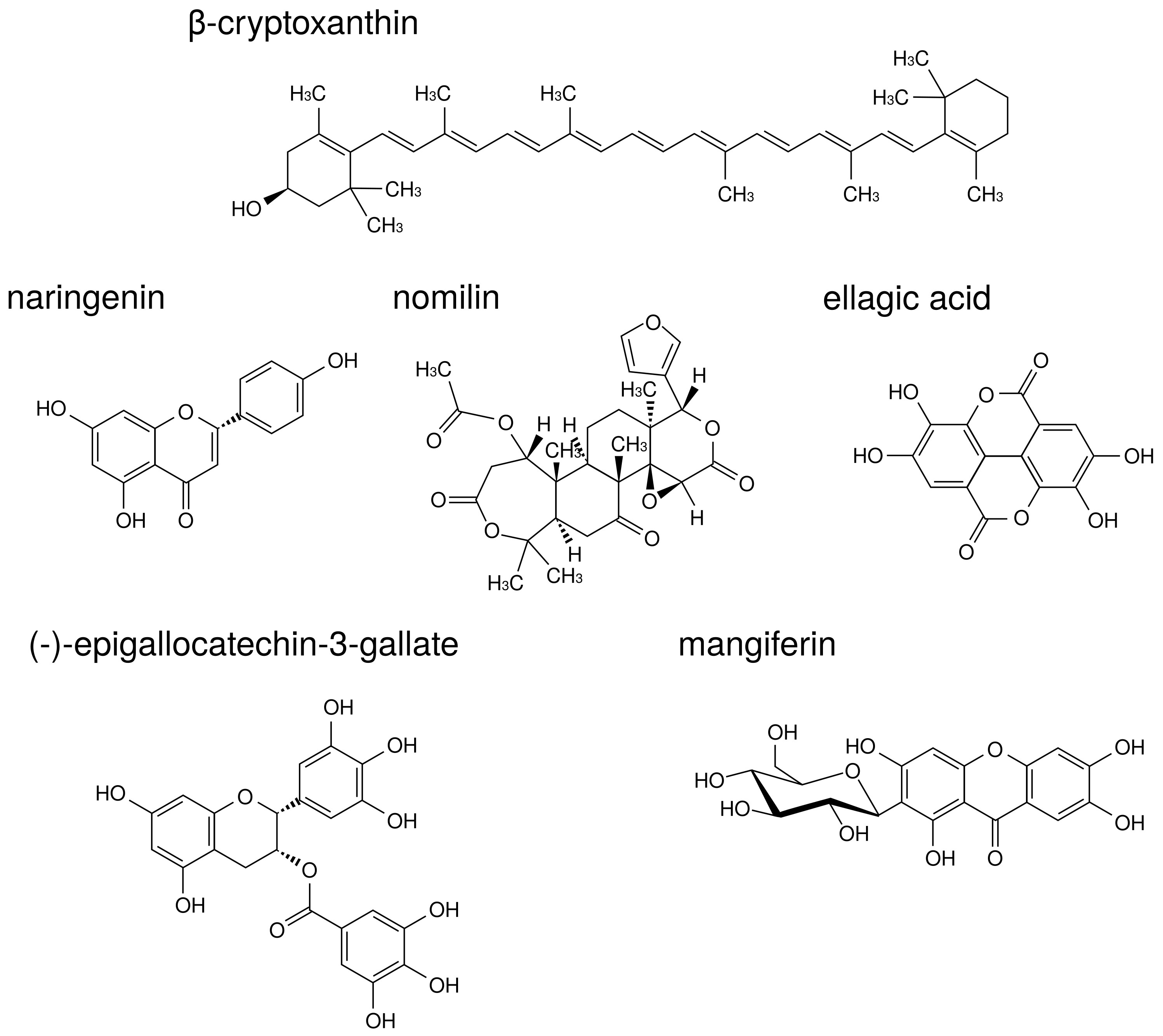
The structures of natural products described in this review are indicated in
Figure 8.
Epidemiological studies have revealed a positive correlation between bone health and increased consumption of fruits and vegetables [
107,
108]. Some fruits and vegetables contain components that inhibit both inflammation and osteoclast activity.
-Cryptoxanthin is a carotenoid present in a wide range of citrus fruits and in
Diospyros kaki Thunb.,
Physalis alkekengi L., etc.
-Cryptoxanthin has a potent inhibitory effect on osteoclast-like cell formation in mouse marrow culture [
109]. Moreover, in a mouse model of PD,
-cryptoxanthin suppressed bone resorption in the mandibular alveolar bone in vitro and restored alveolar bone loss induced by LPS in vivo [
110].
Naringenin is a flavonoid contained in citrus fruits such as oranges and grapefruits. Accumulating evidence has suggested that naringenin modulates chronic inflammation [
111]. In a murine model of collagen-induced arthritis, naringenin inhibited pro-inflammatory cytokine production by decreasing MAPK and NF-
B signaling activation [
112]. La et al. showed naringenin thus holds promise as a therapeutic and preventive agent for bone-related diseases such as PD [
113]. Thus, there are cases in which components demonstrating anti-osteoclast behavior are demonstrated to be effective against PD. In addition to naringenin, citrus fruits contain components that suppress osteoclast activity via MAPK. Nomilin, a limonoid present in citrus fruits, displays inhibitory effects on osteoclastic differentiation through the suppression of MAPK signaling pathways [
114].
Ellagic acid is a polyphenol contained in berries, pomegranates, nuts, etc. Ellagic acid has an anti-inflammatory effect in various organs such as the liver, stomach, small intestine, and skin [
115,
116,
117,
118]. Moreover, ellagic acid has anti-osteoclast activity and significantly reduced serum levels of pro-inflammatory cytokines, TNF-
, IL-1
, and IL-17 in RA model mice [
119]. A recent study supported the traditional use of
Geum urbanum L. root contained ellagic acid derivatives in cavity inflammation including mucositis, gingivitis, and PD [
120].
Additional useful components against both RA and PD have been found in tea. (-)-Epigallocatechin-3-gallate (EGCG) is a major catechin derivative present in green tea. Previous studies have also suggested that EGCG decreases MMP-1, MMP-2, and MMP-3 production by RA synovial fibroblasts, thereby preventing further cartilage and bone destruction [
121,
122]. Moreover, it has been reported that EGCG selectively inhibited IL-1
-induced IL-6 synthesis in RA synovial fibroblasts and suppressed IL-6 trans-signaling via upregulation of an endogenous inhibitor, a soluble gp130 [
123]. Clinical study of EGCG suggested that local drug delivery utilizing green tea extract could be used as an adjunct in the treatment of chronic PD [
124].
Traditional medicine in Ayurveda also presents useful teas against RA and PD.
Salacia reticulata Wight is a plant native to Sri Lanka that has been used for the prevention of RA, gonorrhea, and skin disease. We previously reported that leaf of
S. reticulata alleviates collagen antibody-induced arthritis in RA model mice [
125].
S. reticulata contains a polyphenol known as mangiferin that inhibits osteoclastic bone resorption by promoting ER
mRNA expression in mouse bone marrow macrophage cells [
126].
In conclusion, natural products displaying both anti-inflammation and anti-osteoclast characteristics are suggested to be useful for the prevention and treatment of PD.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}