


Glyphosate-Based Herbicides and Their Potential Impact on the Microbiota of Social Bees
Abstract
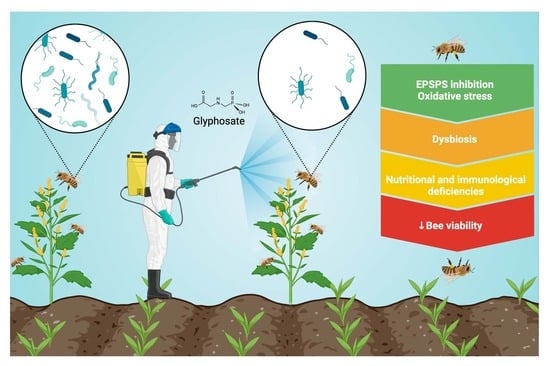
1. Introduction
2. Glyphosate: Chemistry, Usage, and Environmental Impact
3. Effects of Glyphosate Exposure on Honeybee Microbiota
4. Effects of Glyphosate Exposure on Bumblebee Microbiota
5. Functional Consequences of Glyphosate-Induced Dysbiosis in Bees
5.1. Pathogen Defense and Immune Function
5.2. Nutritional Deficiencies, Metabolic Dysregulation, and Development
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPA | Aminomethylphosphonic acid |
| AMP | Antimicrobial peptide |
| GBHs | Glyphosate-based herbicides |
| GM | Genetically modified |
| mM | Millimolar |
| SCFAs | Short-chain fatty acids |
| S. alvi | Snodgrassella alvi |
| POEA | Polyethylated tallow amine |
| EPSPS | 5-enolpyruvylshikimate-3-phosphate synthase |
| 16S rRNA | 16S ribosomal RNA |
| PEP | Phosphoenolpyruvate |
References
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Jankielsohn, A. The importance of insects in agricultural ecosystems. Adv. Entomol. 2018, 6, 62–73. [Google Scholar] [CrossRef]
- Katumo, D.M.; Liang, H.; Ochola, A.C.; Lv, M.; Wang, Q.-F.; Yang, C.-F. Pollinator diversity benefits natural and agricultural ecosystems, environmental health, and human welfare. Plant Divers. 2022, 44, 429–435. [Google Scholar] [CrossRef] [PubMed]
- James, R.R.; Pitts-Singer, T.L. Bee Pollination in Agricultural Ecosystems; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Roubik, D.W. Bees, ecological roles bees: Ecological roles. In Encyclopedia of Social Insects; Starr, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 1–6. [Google Scholar]
- Benton, T. Solitary Bees; Pelagic Publishing Ltd.: London, UK, 2017; Volume 33. [Google Scholar]
- Kevan, P.G.; Viana, B.F. The global decline of pollination services. Biodiversity 2003, 4, 3–8. [Google Scholar] [CrossRef]
- Holden, C. Report warns of looming pollination crisis in North America. Science 2006, 314, 397. [Google Scholar] [CrossRef]
- Millard, J.; Outhwaite, C.L.; Ceaușu, S.; Carvalheiro, L.G.; da Silva e Silva, F.D.; Dicks, L.V.; Ollerton, J.; Newbold, T. Key tropical crops at risk from pollinator loss due to climate change and land use. Sci. Adv. 2023, 9, eadh0756. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Vanbergen, A.J.; Insect Pollinators Initiative. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Klein, S.; Cabirol, A.; Devaud, J.-M.; Barron, A.B.; Lihoreau, M. Why Bees Are So Vulnerable to Environmental Stressors. Trends Ecol. Evol. 2017, 32, 268–278. [Google Scholar] [CrossRef]
- Walker, L.; Wu, S. Pollinators and Pesticides. In International Farm Animal, Wildlife and Food Safety Law; Steier, G., Patel, K.K., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 495–513. [Google Scholar]
- Nath, R.; Singh, H.; Mukherjee, S. Insect pollinators decline: An emerging concern of Anthropocene epoch. J. Apic. Res. 2023, 62, 23–38. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Quandahor, P.; Kim, L.; Kim, M.; Lee, K.; Kusi, F.; Jeong, I.-H. Effects of Agricultural Pesticides on Decline in Insect Species and Individual Numbers. Environments 2024, 11, 182. [Google Scholar] [CrossRef]
- Bamal, D.; Duhan, A.; Pal, A.; Beniwal, R.K.; Kumawat, P.; Dhanda, S.; Goyat, A.; Hooda, V.S.; Yadav, R. Herbicide risks to non-target species and the environment: A review. Environ. Chem. Lett. 2024, 22, 2977–3032. [Google Scholar] [CrossRef]
- Berg, C.J.; King, H.P.; Delenstarr, G.; Kumar, R.; Rubio, F.; Glaze, T. Glyphosate residue concentrations in honey attributed through geospatial analysis to proximity of large-scale agriculture and transfer off-site by bees. PLoS ONE 2018, 13, e0198876. [Google Scholar] [CrossRef]
- de Souza, A.P.F.; Rodrigues, N.R.; Reyes, F.G.R. Glyphosate and aminomethylphosphonic acid (AMPA) residues in Brazilian honey. Food Addit. Contam. Part B 2021, 14, 40–47. [Google Scholar] [CrossRef]
- Muñoz, J.P.; Araya-Osorio, R.; Mera-Adasme, R.; Calaf, G.M. Glyphosate mimics 17β-estradiol effects promoting estrogen receptor alpha activity in breast cancer cells. Chemosphere 2023, 313, 137201. [Google Scholar] [CrossRef]
- Muñoz, J.P.; Bleak, T.C.; Calaf, G.M. Glyphosate and the key characteristics of an endocrine disruptor: A review. Chemosphere 2021, 270, 128619. [Google Scholar] [CrossRef]
- Guyton, K.Z.; Loomis, D.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; Straif, K. Carcinogenicity of tetrachlorvinphos, parathion, malathion, diazinon, and glyphosate. Lancet Oncol. 2015, 16, 490–491. [Google Scholar] [CrossRef]
- Battisti, L.; Potrich, M.; Sampaio, A.R.; de Castilhos Ghisi, N.; Costa-Maia, F.M.; Abati, R.; dos Reis Martinez, C.B.; Sofia, S.H. Is glyphosate toxic to bees? A meta-analytical review. Sci. Total Environ. 2021, 767, 145397. [Google Scholar] [CrossRef]
- Lozano, V.L.; Defarge, N.; Rocque, L.M.; Mesnage, R.; Hennequin, D.; Cassier, R.; de Vendômois, J.S.; Panoff, J.M.; Séralini, G.E.; Amiel, C. Sex-dependent impact of Roundup on the rat gut microbiome. Toxicol. Rep. 2018, 5, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Bridi, D.; Altenhofen, S.; Gonzalez, J.B.; Reolon, G.K.; Bonan, C.D. Glyphosate and Roundup(®) alter morphology and behavior in zebrafish. Toxicology 2017, 392, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Farina, W.M.; Balbuena, M.S.; Herbert, L.T.; Mengoni Goñalons, C.; Vázquez, D.E. Effects of the Herbicide Glyphosate on Honey Bee Sensory and Cognitive Abilities: Individual Impairments with Implications for the Hive. Insects 2019, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Herbert, L.T.; Vázquez, D.E.; Arenas, A.; Farina, W.M. Effects of field-realistic doses of glyphosate on honeybee appetitive behaviour. J. Exp. Biol. 2014, 217, 3457–3464. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Muñoz, J.P.; Silva-Pavez, E.; Carrillo-Beltrán, D.; Calaf, G.M. Occurrence and exposure assessment of glyphosate in the environment and its impact on human beings. Environ. Res. 2023, 231, 116201. [Google Scholar] [CrossRef]
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its Environmental Persistence and Impact on Crop Health and Nutrition. Plants 2019, 8, 499. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616–617, 255–268. [Google Scholar] [CrossRef]
- Shehata, A.A.; Schrödl, W.; Aldin, A.A.; Hafez, H.M.; Krüger, M. The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr. Microbiol. 2013, 66, 350–358. [Google Scholar] [CrossRef]
- Nandula, V.K. Glyphosate Resistance in Crops and Weeds: History, Development, and Management; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Franz, J.E.; Mao, M.K.; Sikorski, J.A. Glyphosate: A Unique Global Herbicide; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Tarazona, J.V.; Court-Marques, D.; Tiramani, M.; Reich, H.; Pfeil, R.; Istace, F.; Crivellente, F. Glyphosate toxicity and carcinogenicity: A review of the scientific basis of the European Union assessment and its differences with IARC. Arch. Toxicol. 2017, 91, 2723–2743. [Google Scholar] [CrossRef] [PubMed]
- Martins-Gomes, C.; Silva, T.L.; Andreani, T.; Silva, A.M. Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity. J. Xenobiotics 2022, 12, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Howe, C.M.; Berrill, M.; Pauli, B.D.; Helbing, C.C.; Werry, K.; Veldhoen, N. Toxicity of glyphosate-based pesticides to four North American frog species. Environ. Toxicol. Chem. Int. J. 2004, 23, 1928–1938. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Silva, L.d.; Araújo, H.H.; Meireles, C.S.; Silva, L.C.d. Plant exposure to glyphosate-based herbicides and how this might alter plant physiological and structural processes. Botany 2022, 100, 473–480. [Google Scholar] [CrossRef]
- Boocock, M.R.; Coggins, J.R. Kinetics of 5-enolpyruvylshikimate-3-phosphate synthase inhibition by glyphosate. FEBS Lett. 1983, 154, 127–133. [Google Scholar] [CrossRef]
- Sikorski, J.A.; Gruys, K.J. Understanding Glyphosate’s Molecular Mode of Action with EPSP Synthase: Evidence Favoring an Allosteric Inhibitor Model. Acc. Chem. Res. 1997, 30, 2–8. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Wang, L.; Peng, R.; Tian, Y.; Han, J.; Zhao, W.; Wang, B.; Liu, M.; Yao, Q. Characterization of a class II 5-enopyruvylshikimate-3-phosphate synthase with high tolerance to glyphosate from Sinorhizobium fredii. World J. Microbiol. Biotechnol. 2014, 30, 2967–2973. [Google Scholar] [CrossRef]
- Thiour-Mauprivez, C.; Martin-Laurent, F.; Calvayrac, C.; Barthelmebs, L. Effects of herbicide on non-target microorganisms: Towards a new class of biomarkers? Sci. Total Environ. 2019, 684, 314–325. [Google Scholar] [CrossRef]
- Duke, S.O. Interaction of Chemical Pesticides and Their Formulation Ingredients with Microbes Associated with Plants and Plant Pests. J. Agric. Food Chem. 2018, 66, 7553–7561. [Google Scholar] [CrossRef] [PubMed]
- Pline-Srnic, W. Physiological Mechanisms of Glyphosate Resistance. Weed Technol. 2006, 20, 290–300. [Google Scholar] [CrossRef]
- Battaglin, W.A.; Meyer, M.T.; Kuivila, K.M.; Dietze, J.E. Glyphosate and Its Degradation Product AMPA Occur Frequently and Widely in U.S. Soils, Surface Water, Groundwater, and Precipitation. JAWRA J. Am. Water Resour. Assoc. 2014, 50, 275–290. [Google Scholar] [CrossRef]
- Panis, C.; Candiotto, L.Z.P.; Gaboardi, S.C.; Gurzenda, S.; Cruz, J.; Castro, M.; Lemos, B. Widespread pesticide contamination of drinking water and impact on cancer risk in Brazil. Environ. Int. 2022, 165, 107321. [Google Scholar] [CrossRef]
- Thompson, T.S.; van den Heever, J.P.; Limanowka, R.E. Determination of glyphosate, AMPA, and glufosinate in honey by online solid-phase extraction-liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A 2019, 36, 434–446. [Google Scholar] [CrossRef]
- Rampazzo, G.; Nobile, M.; Carpino, S.; Chiesa, L.; Ghidini, S.; Gazzotti, T.; Panseri, S. Detection of glyphosate, glufosinate, and their metabolites in multi-floral honey for food safety. Food Addit. Contam. Part A 2025, 42, 213–222. [Google Scholar] [CrossRef]
- Mesnage, R.; Teixeira, M.; Mandrioli, D.; Falcioni, L.; Ducarmon, Q.R.; Zwittink, R.D.; Mazzacuva, F.; Caldwell, A.; Halket, J.; Amiel, C.; et al. Use of Shotgun Metagenomics and Metabolomics to Evaluate the Impact of Glyphosate or Roundup MON 52276 on the Gut Microbiota and Serum Metabolome of Sprague-Dawley Rats. Environ. Health Perspect. 2021, 129, 17005. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018, 9, 372026. [Google Scholar] [CrossRef]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect. Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Wang, K.; Li, J.; Zhao, L.; Mu, X.; Wang, C.; Wang, M.; Xue, X.; Qi, S.; Wu, L. Gut microbiota protects honey bees (Apis mellifera L.) against polystyrene microplastics exposure risks. J. Hazard. Mater. 2021, 402, 123828. [Google Scholar] [CrossRef] [PubMed]
- Cornet, L.; Cleenwerck, I.; Praet, J.; Leonard, R.R.; Vereecken, N.J.; Michez, D.; Smagghe, G.; Baurain, D.; Vandamme, P. Phylogenomic analyses of Snodgrassella isolates from honeybees and bumblebees reveal taxonomic and functional diversity. Msystems 2022, 7, e01500–e01521. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey Bee Gut Microbiome Is Altered by In-Hive Pesticide Exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Raymann, K.; Moran, N.A. Glyphosate perturbs the gut microbiota of honey bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [PubMed]
- Motta, E.V.S.; Mak, M.; De Jong, T.K.; Powell, J.E.; O’Donnell, A.; Suhr, K.J.; Riddington, I.M.; Moran, N.A. Oral or Topical Exposure to Glyphosate in Herbicide Formulation Impacts the Gut Microbiota and Survival Rates of Honey Bees. Appl. Environ. Microbiol. 2020, 86, e01150-20. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Moran, N.A. Impact of Glyphosate on the Honey Bee Gut Microbiota: Effects of Intensity, Duration, and Timing of Exposure. Msystems 2020, 5, e00268-20. [Google Scholar] [CrossRef]
- Blot, N.; Veillat, L.; Rouzé, R.; Delatte, H. Glyphosate, but not its metabolite AMPA, alters the honeybee gut microbiota. PLoS ONE 2019, 14, e0215466. [Google Scholar] [CrossRef]
- Castelli, L.; Balbuena, S.; Branchiccela, B.; Zunino, P.; Liberti, J.; Engel, P.; Antúnez, K. Impact of chronic exposure to sublethal doses of glyphosate on honey bee immunity, gut microbiota and infection by pathogens. Microorganisms 2021, 9, 845. [Google Scholar] [CrossRef]
- Quinn, A.; El Chazli, Y.; Escrig, S.; Daraspe, J.; Neuschwander, N.; McNally, A.; Genoud, C.; Meibom, A.; Engel, P. Host-derived organic acids enable gut colonization of the honey bee symbiont Snodgrassella alvi. Nat. Microbiol. 2024, 9, 477–489. [Google Scholar] [CrossRef]
- Powell, J.E.; Leonard, S.P.; Kwong, W.K.; Engel, P.; Moran, N.A. Genome-wide screen identifies host colonization determinants in a bacterial gut symbiont. Proc. Natl. Acad. Sci. USA 2016, 113, 13887–13892. [Google Scholar] [CrossRef]
- Motta, E.V.S.; de Jong, T.K.; Gage, A.; Edwards, J.A.; Moran, N.A. Glyphosate effects on growth and biofilm formation in bee gut symbionts and diverse associated bacteria. Appl. Environ. Microbiol. 2024, 90, e0051524. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Yan, Z.; Ma, S.; Yang, Y.; Wang, Q.; Hou, C.; Wu, Y.; Liu, Y.; Diao, Q. The Herbicide Glyphosate Negatively Affects Midgut Bacterial Communities and Survival of Honey Bee During Larvae Reared in Vitro. J. Agric. Food Chem. 2018, 66, 7786–7793. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, D.E.; Martínez, L.E.V.; Medina, V.; Latorre-Estivalis, J.M.; Zavala, J.A.; Farina, W.M. Glyphosate affects larval gut microbiota and metamorphosis of honey bees with differences between rearing procedures. Environ. Pollut. 2023, 334, 122200. [Google Scholar] [CrossRef] [PubMed]
- Almasri, H.; Liberti, J.; Brunet, J.-L.; Engel, P.; Belzunces, L.P. Mild chronic exposure to pesticides alters physiological markers of honey bee health without perturbing the core gut microbiota. Sci. Rep. 2022, 12, 4281. [Google Scholar] [CrossRef]
- Motta, E.V.; Powell, J.E.; Moran, N.A. Glyphosate induces immune dysregulation in honey bees. Anim. Microbiome 2022, 4, 16. [Google Scholar] [CrossRef]
- Ma, C.; Gu, G.; Chen, S.; Shi, X.; Li, Z.; Li-Byarlay, H.; Bai, L. Impact of chronic exposure to field level glyphosate on the food consumption, survival, gene expression, gut microbiota, and metabolomic profiles of honeybees. Environ. Res. 2024, 250, 118509. [Google Scholar] [CrossRef]
- Abrol, D.; Mondal, A.; Shankar, U. Importance of bumble bees for crop pollination and food security. J. Palynol. 2021, 57, 9–37. [Google Scholar]
- Ana, N. European Red List of Bees; International Union for Conservation of Nature: Gland, Switzerland; European Commission, IUCN European Union Representative Office: Brussels, Belgium; IUCN Species Survival Commission (SSC), Bumblebee Specialist Group: Gland, Switzerland, 2014. [Google Scholar]
- Goulson, D. Bumblebees: Their Behaviour and Ecology; Oxford University Press: New York, NY, USA, 2003. [Google Scholar]
- Hatfield, R.; Jepsen, S.; Mader, E.; Black, S.H.; Shepherd, M. Conserving Bumble Bees; Xerces Society for Invertebrate Conservation: Portland, Oregon, 2012. [Google Scholar]
- Ahmad, M.; Bodlah, I.; Mehmood, K.; Sheikh, U.A.A.; Aziz, M.A. Pollination and foraging potential of European bumblebee, Bombus terrestris (Hymenoptera: Apidae) on tomato crop under greenhouse system. Pak. J. Zool. 2015, 47, 1279–1285. [Google Scholar]
- Hammer, T.J.; Le, E.; Martin, A.N.; Moran, N.A. The gut microbiota of bumblebees. Insectes Sociaux 2021, 68, 287–301. [Google Scholar] [CrossRef]
- Hammer, T.J.; Easton-Calabria, A.; Moran, N.A. Microbiome assembly and maintenance across the lifespan of bumble bee workers. Mol. Ecol. 2023, 32, 724–740. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Moran, N.A. The effects of glyphosate, pure or in herbicide formulation, on bumble bees and their gut microbial communities. Sci. Total Environ. 2023, 872, 162102. [Google Scholar] [CrossRef] [PubMed]
- Helander, M.; Jeevannavar, A.; Kaakinen, K.; Mathew, S.A.; Saikkonen, K.; Fuchs, B.; Puigbò, P.; Loukola, O.J.; Tamminen, M. Glyphosate and a glyphosate-based herbicide affect bumblebee gut microbiota. FEMS Microbiol. Ecol. 2023, 99, fiad065. [Google Scholar] [CrossRef] [PubMed]
- Cullen, M.G.; Bliss, L.; Stanley, D.A.; Carolan, J.C. Investigating the effects of glyphosate on the bumblebee proteome and microbiota. Sci. Total Environ. 2023, 864, 161074. [Google Scholar] [CrossRef] [PubMed]
- Straw, E.A.; Mesnage, R.; Brown, M.J.; Antoniou, M.N. No impacts of glyphosate or Crithidia bombi, or their combination, on the bumblebee microbiome. Sci. Rep. 2023, 13, 8949. [Google Scholar] [CrossRef]
- Tang, Q.-H.; Li, W.-L.; Wang, J.-P.; Li, X.-J.; Li, D.; Cao, Z.; Huang, Q.; Li, J.-L.; Zhang, J.; Wang, Z.-W. Effects of spinetoram and glyphosate on physiological biomarkers and gut microbes in Bombus terrestris. Front. Physiol. 2023, 13, 1054742. [Google Scholar] [CrossRef]
- Hotchkiss, M.Z.; Forrest, J.R.K.; Poulain, A.J. Changes in bumblebee queen gut microbiotas during and after overwintering diapause. Insect Mol. Biol. 2025, 34, 136–150. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A.; et al. Gut microbiome dysbiosis and honeybee health. J. Appl. Entomol. 2011, 135, 524–533. [Google Scholar] [CrossRef]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef]
- Steele, M.I.; Motta, E.V.S.; Gattu, T.; Martinez, D.; Moran, N.A. The Gut Microbiota Protects Bees from Invasion by a Bacterial Pathogen. Microbiol. Spectr. 2021, 9, e0039421. [Google Scholar] [CrossRef]
- Botina, L.L.; Barbosa, W.F.; Acosta, J.P.L.; Bernardes, R.C.; Cortes, J.E.Q.; Pylro, V.S.; Mendonça, A.C.; Barbosa, R.C.; Lima, M.A.P.; Martins, G.F. The impact of early-life exposure to three agrochemicals on survival, behavior, and gut microbiota of stingless bees (Partamona helleri). Environ. Sci. Pollut. Res. 2023, 30, 70143–70158. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef] [PubMed]
- Weidenmüller, A.; Meltzer, A.; Neupert, S.; Schwarz, A.; Kleineidam, C. Glyphosate impairs collective thermoregulation in bumblebees. Science 2022, 376, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Voulgari-Kokota, A.; McFrederick, Q.S.; Steffan-Dewenter, I.; Keller, A. Drivers, Diversity, and Functions of the Solitary-Bee Microbiota. Trends Microbiol. 2019, 27, 1034–1044. [Google Scholar] [CrossRef]
- Thamm, M.; Reiß, F.; Sohl, L.; Gabel, M.; Noll, M.; Scheiner, R. Solitary Bees Host More Bacteria and Fungi on Their Cuticle than Social Bees. Microorganisms 2023, 11, 2780. [Google Scholar] [CrossRef]
- Hettiarachchi, A.; Cnockaert, M.; Joossens, M.; Gekière, A.; Meeus, I.; Vereecken, N.J.; Michez, D.; Smagghe, G.; Vandamme, P. The wild solitary bees Andrena vaga, Anthophora plumipes, Colletes cunicularius, and Osmia cornuta microbiota are host specific and dominated by endosymbionts and environmental microorganisms. Microb. Ecol. 2023, 86, 3013–3026. [Google Scholar] [CrossRef]
- Gandhi, K.; Khan, S.; Patrikar, M.; Markad, A.; Kumar, N.; Choudhari, A.; Sagar, P.; Indurkar, S. Exposure risk and environmental impacts of glyphosate: Highlights on the toxicity of herbicide co-formulants. Environ. Chall. 2021, 4, 100149. [Google Scholar] [CrossRef]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent—Relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef]
- Battisti, L.; Potrich, M.; Abati, R.; Sampaio, A.R.; Libardoni, G.; Costa-Maia, F.M.; Berté, E.A.; Dos Reis Martinez, C.B.; Sofia, S.H. Toxicity of glyphosate herbicides formulated for Africanized Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae). Ecotoxicol. Environ. Saf. 2024, 287, 117247. [Google Scholar] [CrossRef]
- Vázquez, D.E.; Verellen, F.; Farina, W.M. Early exposure to glyphosate during larval development induces late behavioural effects on adult honey bees. Environ. Pollut. 2024, 360, 124674. [Google Scholar] [CrossRef]
- Tomé, H.V.V.; Schmehl, D.R.; Wedde, A.E.; Godoy, R.S.M.; Ravaiano, S.V.; Guedes, R.N.C.; Martins, G.F.; Ellis, J.D. Frequently encountered pesticides can cause multiple disorders in developing worker honey bees. Environ. Pollut. 2020, 256, 113420. [Google Scholar] [CrossRef]
- Zhi-Xiang, D.; Wan-Li, L.; Xi-Jie, L.I.; Jia-Li, L.; Jun, Z.; Chong-Hui, Z.; Qi, H.; Zhe, C.; Yuan, C.; Hong-Mu, Z.; et al. Glyphosate exposure affected longevity-related pathways and reduced survival in asian honey bees (Apis cerana). Chemosphere 2024, 351, 141199. [Google Scholar] [CrossRef]

{kind=link}
| Ref. | Compound Type | Concentration Used | Time of Exposure | Route of Exposure | Developmental Stage | Observed Effects |
|---|---|---|---|---|---|---|
| Motta et al. (2022) [71] | Glyphosate standard (≥95% purity) | (i) In vivo assays: 0, 16.91, 169.1, and 1690.7 mg/L. (ii) Ex vivo assays: 0, 16.91, 169.1, 338.1, 676.3, 1183.5, and 1690.7 mg/L. | 5 days | (i) Oral (in sucrose syrup) (ii) Ex vivo (hemolymph assays) | Newly emerged adult bees (1–5 days old); hive worker bees | Downregulation of antimicrobial peptide (AMP) genes (e.g., apidaecin, defensin, hymenoptaecin); dysbiosis (e.g., reduced S. alvi and Gilliamella); inhibition of melanization at ≥2 mM (only ex vivo); increased immune dysregulation. |
| Vázquez et al. (2023) [69] | Glyphosate (PESTANAL standard, ≥99.2% purity) | In vitro exposure: 0.07 and 2.5 mg/L | 0–144 h (chronic) 72–144 h (subchronic) | Direct ingestion of contaminated food | Larval (primarily 3rd–5th instars) | In vitro exposure: Dysbiosis with loss of core gut bacteria and increase in environmental bacteria; delayed larval development; increased teratogenesis and mortality during larval and pupal stages; surviving adults heavier but with reduced post-emergence survival. |
| In-hive exposure: 0.102 mg/L | Post-supplementation with syrup | Indirect ingestion via contaminated honey/beebread | In-hive exposure: Early larval mortality likely linked to hygienic behavior; reduced adult survival; milder dysbiosis compared to in vitro exposure; slight developmental delays. | |||
| Ma et al. (2024) [72] | Glyphosate standard (≥99.5% purity) | 5 mg/L | 10 days | Oral (via sucrose solution) | Newly emerged adult worker bees (1 day old) | Reduced sugar consumption (significant on day 10); decreased survival probability; no significant change in gut microbiota composition or diversity; significant downregulation of glucose dehydrogenase, vitellogenin, esterase FE4, and CYP6AQ1 genes. |
| Motta et al. (2020) [61] | Glyphosate standard (≥95% purity) | 169.07 mg/L | 5 days | Oral (feeding with syrup) | Newly emerged adult bees | Significant reduction in S. alvi abundance; altered gut microbiota composition; no significant mortality compared to control. |
| Roundup ProMax (48.7% glyphosate potassium salt) | (i) 169.07 mg/L a.e. (lab). (ii) 540 mg/L a.e. Roundup (field) (iii) 270–16,200 mg/L a.e. (topical) | (i) 5 days (lab) (ii) Weekly for 1 month (field) (iii) Single dose | Oral (syrup or water) and topical (spray exposure) | Newly emerged and adult bees | Stronger reduction in Snodgrassella, Gilliamella, and Bifidobacterium than with pure glyphosate; increased mortality (dose-dependent) after topical exposure; reduced hive return rates; transfer of glyphosate to honey within hive; increased susceptibility to Serratia marcescens infection. | |
| Almasri et al. (2022) [70] | Glyphosate standard. Purity not specified (NS) | 0.0001 mg/L | 5 days | Oral (in sucrose solution) | Newly emerged adult bees | No significant effect on core gut microbiota composition or total bacterial load; altered physiological markers (e.g., increased LDH activity in head, increased GST in midgut in microbiota-depleted bees); no impact on food consumption or survival; effects more pronounced in microbiota-depleted bees, suggesting gut microbiota buffers toxicity. |
| Castelli et al. (2021) [64] | Glyphosate standard (Sigma-Aldrich, Burlington, MA, USA, purity ≥95%) | 10 mg/L | 7 to 14 days (chronic exposure) | Oral (in sucrose solution) | Newly emerged adult bees (≤24 h post-emergence) | Altered gut microbiota composition: ↓ S. alvi, ↑ G. apicola, Lactobacillus kimbladii, Staphylococcus; increased alpha and beta diversity; increased expression of lysozyme and glucose oxidase (immune response genes); decreased vitellogenin expression (related to longevity and health); increased DWV replication; significantly reduced lifespan (LT50 = 13 days vs. 20 days control). |
| Blot et al. (2019) [63] | Glyphosate standard (Interchim, Montluçon, France, SS-7701, purity ≥95%) | 253.6 and 1268 mg/L | 15 days (chronic) | Oral (via sugar syrup) | Adult worker bees (interior workers, overwintering and summer) | Significant dose-independent decrease in S. alvi; decrease in G. apicola; increase in Lactobacillus spp., especially Firm-5 (at 1268 mg/L); no effect on Bifidobacterium spp. or Alphaproteobacteria; in vitro growth of S. alvi, G. apicola, Bifidobacterium spp. inhibited; no significant effect on survival or food consumption. |
| AMPA standard (Sigma-Aldrich, purity ≥95%) | 204.1 and 952.2 mg/L | 15 days (chronic) | Oral (via sugar syrup) | Adult worker bees (interior workers, summer) | No significant change in gut bacterial composition in vivo. In vitro: inhibited G. apicola at higher concentration (5 mM). No effect on S. alvi, Bifidobacterium spp. or Lactobacillus spp.; no impact on survival or food intake. | |
| Motta et al. (2024) [67] | Glyphosate standard. Purity NS | 1.69, 16.91, 169.1, 338.1, 676.3, 1014.4, 1352.5, and 1690.7 mg/L | 48 h (in vitro assays) | In vitro (in media with bacterial cultures) | Bacterial strains isolated from adult bee guts | Strain-specific, dose-dependent effects on bacterial growth and biofilm formation; inhibited biofilm formation in S. alvi, Gilliamella, and others; upregulation of EPSPS and TrpC enzymes; altered proteomic profile in S. alvi. |
| Roundup ProMax (48.7% glyphosate potassium salt) | 1.69, 16.91, 169.1, 338.1, 676.3, 1014.4, 1352.5, and 1690.7 mg/L a.e. | 48 h (in vitro assays) | In vitro (in media with bacterial cultures) | Bacterial strains isolated from adult bee guts | More pronounced effects compared to glyphosate alone; low doses often stimulated growth or biofilm formation, while high doses inhibited both; formulation co-factors likely contributed to divergent outcomes; stronger disruption observed in Snodgrassella compared to Gilliamella and Lactobacillus. | |
| Motta and Moran (2020) [62] | Glyphosate standard. Purity NS | 1.69, 6.76, 11.83, 16.91, and 169.1 mg/L | 15–20 days (chronic exposure) | Oral (in sucrose syrup) | Newly emerged bees (1-day-old) and bees with established microbiota (5-day-old) | Dose-dependent reduction in S. alvi and Gilliamella; increase in Bifidobacterium and Lactobacillus (Firm-4/5); greater mortality at ≥0.1 mM; effects observed regardless of timing of exposure (early or late microbiota acquisition); gut microbiota disruption consistent across trials; no significant alpha diversity change but altered community structure. |
| Motta et al. (2018) [60] | Glyphosate standard. Purity NS | 5 and 10 mg/L | 5 days (oral), plus 3 days post-reintroduction | Oral (in sucrose syrup) | Adult bees with established microbiota and newly emerged bees | Significant reduction in S. alvi and other species (e.g., Bifidobacterium spp., Lactobacillus Firm-4/5); increased relative abundance of G. apicola; impaired colonization during early gut development; increased mortality upon infection with S. marcescens; glyphosate-sensitive EPSPS class I associated with reduced bacterial growth. |
| Dai et al. (2018) [68] | Glyphosate standard (Aladdin, Shanghai, China, ≥99.5% purity) | 0.8, 4, and 20 mg/L | 4 days (D2–D5 post-grafting) | Oral (in artificial larval diet) | Larval stage | Survival: Significant reduction at 4 and 20 mg/L; larval weight: decreased at 0.8 and 4 mg/L; developmental rate: not significantly affected; gut microbiota: at 20 mg/L, significant alteration in midgut bacterial composition (e.g., ↑ Lachnospiraceae, Prevotellaceae, Ruminococcaceae) and reduced beta diversity; specific taxonomic shifts in response to different concentrations. |
| Ref. | Compound Type | Concentration Used | Time of Exposure | Route of Exposure | Developmental Stage During Exposure | Observed Effects |
|---|---|---|---|---|---|---|
| Tang et al. (2023) [84] | Glyphosate ammonium salt (30% glyphosate acid, purity NS) | 2.5 mg/L (sublethal) | 10 days | Oral (via sucrose syrup) | Adult workers | Increased activity of superoxide dismutase (SOD) and prophenoloxidase (PPO), suggesting oxidative stress and immune activation; significantly reduced gut α-amylase activity, suggesting impaired digestion and energy metabolism; no significant effect on glutathione-S-transferase (GST), carboxylesterase (CarE), or protease activities; no significant changes in the core gut bacterial community; significantly altered gut fungal community composition: reduced Zygosaccharomyces, increased Cladosporium, and increased fungal diversity. |
| Motta and Moran (2023) [80] | Glyphosate standard (Caisson Laboratories, Smithfield, UT, USA, purity ≥95%) | 1.69, 16.91, and 169.1 mg/L | 5–7-day post-exposure monitoring | Oral (via sucrose syrup) | Adult workers | Transient reduction in Snodgrassella abundance (in 2 of 4 colonies) after exposure, with recovery by day 7 post-exposure; no significant impact on overall bacterial load or survival; no significant effects on bee weight or syrup consumption. |
| Roundup ProMax® (48.7% glyphosate potassium salt) | 1.69, 16.91, and 169.1 mg/L a.e. | 5–7-day post-exposure monitoring | Oral (via sucrose syrup) | Adult workers | Reduced Snodgrassella abundance in some colonies, reversible by day 7; significant reduction in survival post-exposure at 1 mM concentration; increased syrup consumption at 1 mM; no major impact on total bacterial abundance or bee weight. | |
| Helander et al. (2023) [81] | Glyphosate (PESTANAL standard, ≥95% purity) | 10 and 5000 mg/L (in 60% sucrose solution) | 3 and 5 days | Oral (via colony feeding) | Adult workers | Increased gut microbiota diversity in a dose- and time-dependent manner; reduced relative abundance of Snodgrasella alvi (EPSPS Class I—sensitive); increased abundance of potentially resistant genera like Candidatus Schmidhempelia, Acinetobacter, and Weissella; no significant mortality effect. |
| Roundup Gold (glyphosate isopropylamine salt, 450 g/L a.i.) | 10 mg/L a.e. and 5000 mg/L a.e. | 3 and 5 days | Oral (via colony feeding) | Adult workers | Decreased microbiota diversity at low dose; high dose did not reduce diversity further; significant increase in mortality at high dose; disruption of gut microbial composition, with reduced Snodgrasella and increased Klebsiella, Candidatus Schmidhempelia, and Lactobacillus. | |
| Straw et al. (2023) [83] | Glyphosate standard (Sigma Aldrich, ≥95% purity) | 200 µg (acute single oral dose) equivalent to 4 mg/L (assuming ~50 µL ingestion volume) | 48 h | Oral (via sucrose solution) | Adult workers | No significant effects on survival, sucrose consumption, weight change, parasite (Crithidia bombi) intensity, or gut bacterial microbiome composition; microbiota diversity and relative abundance of major taxa (e.g., Snodgrassella, Gilliamella, Lactobacillus) remained unaffected. |
| Cullen et al. (2023) [82] | Glyphosate standard (purity NS) | 1, 10, and 100 mg/L | 5–10 days | Oral (via 40% sucrose solution) | Adult workers | No significant effects on survival, behavior, or sucrose consumption; significant changes in digestive tract proteome, particularly proteins linked to mitochondrial function, oxidative stress regulation, and structural integrity (e.g., collagen, fibrillin); altered abundance of proteins involved in immune response and detoxification; some minor changes in fungal microbiota, but no significant changes in bacterial community composition. |
| RoundUp Optima+® | 1, 10, and 100 mg/L a.e. | 5–10 days | Oral (via 40% sucrose solution) | Adult workers | No significant impact on survival, behavior, or food intake; distinct proteomic alterations from the pure glyphosate treatment: more pronounced changes in oxidative phosphorylation, lysosomal proteins, and lipid metabolism; disruption of fungal gut microbiota, especially reduced Candida abundance and increased abundance of Tomentella, Trichoderma, Filobasidium, and Archaeorhizomyces; shared effects with glyphosate on structural proteins (collagen, fibrillin), oxidative stress markers, and signaling pathways. |
| Bee Species | Core Gut Microbiota a | Glyphosate Sensitivity | Observed Microbiome Effects | Other Physiological Effects |
|---|---|---|---|---|
| A. mellifera | S. alvi, G. apicola, Lactobacillus Firm-4, Lactobacillus Firm-5, Bifidobacterium spp., Bombilactobacillus spp. | High; S. alvi and G. apicola particularly sensitive; altered biofilm formation and abundance | Dysbiosis (↓ S. alvi, G. apicola), increased alpha and beta diversity; altered immune-related gene expression; fungal dysbiosis (↑ Candida, ↓ Zygosaccharomyces) | Altered expression of AMPs (defensin, apidaecin), ↓ vitellogenin; increased susceptibility to pathogens; reduced longevity; metabolic changes |
| B. terrestris | S. alvi, G. apicola, Schmidhempelia spp., Lactobacillus spp., Bifidobacterium spp. | Moderate; dose- and strain-dependent effects on microbial diversity and core taxa | Variable shifts in microbial composition; ↓ S. alvi, ↑ Klebsiella, Acinetobacter, and Weissella; fungal diversity changes at low doses of GBHs | Proteomic alterations in digestive tissues (collagen, oxidative stress proteins); high doses of GBH increase mortality |
| B. impatiens | S. alvi, G. apicola, Lactobacillus spp., Bifidobacterium spp. | Moderate to low; microbiota more resilient with reversible effects post-exposure | Transient reduction in S. alvi, minimal effect on total bacterial abundance or community structure | Slight increase in syrup consumption at higher concentrations; reduced survival only after formulation exposure |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz, J.P.; Soto-Jiménez, D.; Brito, A.; Quezada-Romegialli, C. Glyphosate-Based Herbicides and Their Potential Impact on the Microbiota of Social Bees. Toxics 2025, 13, 551. https://doi.org/10.3390/toxics13070551
Muñoz JP, Soto-Jiménez D, Brito A, Quezada-Romegialli C. Glyphosate-Based Herbicides and Their Potential Impact on the Microbiota of Social Bees. Toxics. 2025; 13(7):551. https://doi.org/10.3390/toxics13070551
Chicago/Turabian StyleMuñoz, Juan P., Diego Soto-Jiménez, Anghel Brito, and Claudio Quezada-Romegialli. 2025. "Glyphosate-Based Herbicides and Their Potential Impact on the Microbiota of Social Bees" Toxics 13, no. 7: 551. https://doi.org/10.3390/toxics13070551
APA StyleMuñoz, J. P., Soto-Jiménez, D., Brito, A., & Quezada-Romegialli, C. (2025). Glyphosate-Based Herbicides and Their Potential Impact on the Microbiota of Social Bees. Toxics, 13(7), 551. https://doi.org/10.3390/toxics13070551