
The Levels of Polycyclic Aromatic Hydrocarbons and Their Derivatives in Plasma and Their Effect on Mitochondrial DNA Methylation in the Oilfield Workers



 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. Determination of PAHs and PAH Derivatives in Plasma
2.2.1. Pretreatment
2.2.2. GC–MS Analysis
2.3. DNA Methylation Analysis
2.4. Quality Assurance and Quality Control (QA and QC)
2.5. Statistical Analysis
3. Results
3.1. PAH, MPAH, and OPAH Concentration Levels
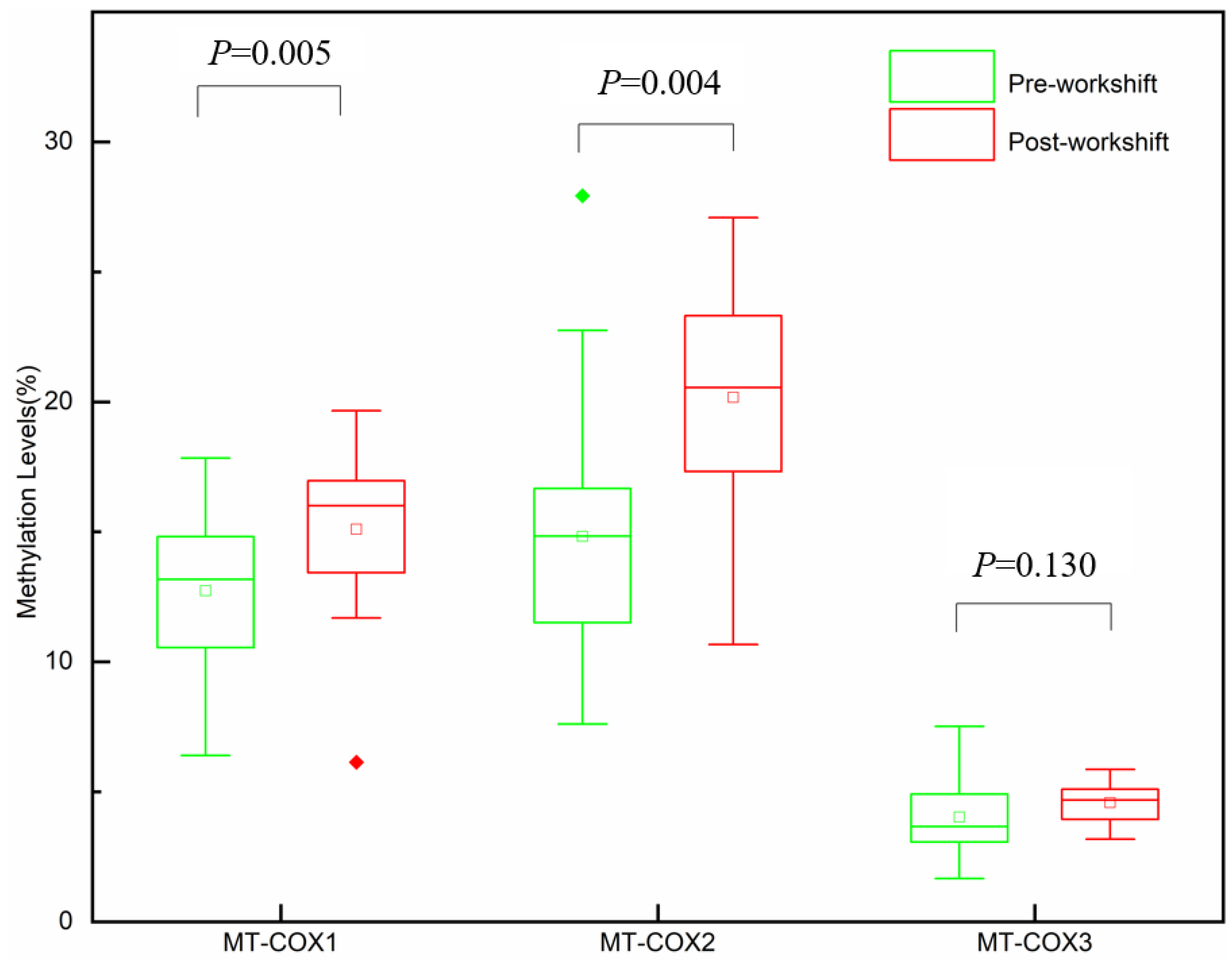
3.2. mtDNA Methylation Levels
3.3. Association between mtDNA Methylation and Pollutant Exposure Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nadal, M.; Schuhmacher, M.; Domingo, J. Levels of PAHs in soil and vegetation samples from Tarragona County, Spain. Environ. Pollut. 2004, 132, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Wu, S.L.; Zhou, H.; Wang, B.; Li, H.; Chen, Y.; Yu, Y.; Shi, Y. Polycyclic Aromatic Hydrocarbons in Soils from Urban to Rural Areas in Nanjing: Concentration, Source, Spatial Distribution, and Potential Human Health Risk. Sci. Total Environ. 2015, 527–528, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Lokeshkumar, B.; Rajendran, P.; Nishigaki, I. Exposure to polycyclic aromatic hydrocarbons with special focus on cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 182–189. [Google Scholar] [CrossRef]
- Orecchio, S.; Amorello, D.; Indelicato, R.; Barreca, S.; Orecchio, S. A Short Review of Simple Analytical Methods for the Evaluation of PAHs and PAEs as Indoor Pollutants in House Dust Samples. Atmosphere 2022, 13, 1799. [Google Scholar] [CrossRef]
- Orecchio, S.; Bianchini, F.; Bonsignore, R.; Blandino, P.; Barreca, S.; Amorello, D. Profiles and Sources of PAHs in Sediments from an Open-Pit Mining Area in the Peruvian Andes. Polycycl. Aromat. Compd. 2016, 36, 429–451. [Google Scholar] [CrossRef]
- Ravindra, K.; Sokhi, R.; Van Grieken, R. Atmospheric Polycyclic Aromatic Hydrocarbons: Source Attribution, Emission Factors and Regulation. Atmos. Environ. 2008, 42, 2895–2921. [Google Scholar] [CrossRef]
- Pickering, R.W. A Toxicological Review of Polycyclic Aromatic Hydrocarbons. J. Toxicol. Cutan. Ocul. Toxicol. 1999, 18, 101–135. [Google Scholar] [CrossRef]
- Blasco, M.; Domeño, C.; Nerín, C. Use of Lichens as Pollution Biomonitors in Remote Areas: Comparison of PAHs Extracted from Lichens and Atmospheric Particles Sampled in and Around the Somport Tunnel (Pyrenees). Environ. Sci. Technol. 2006, 40, 6384–6391. [Google Scholar] [CrossRef]
- Oleszczuk, P.; Baran, S. Polycyclic Aromatic Hydrocarbons Content in Shoots and Leaves of Willow (Salix viminalis) Cultivated on the Sewage Sludge-Amended Soil. Water, Air, Soil Pollut. 2005, 168, 91–111. [Google Scholar] [CrossRef]
- Tidwell, L.G.; Allan, S.E.; O’Connell, S.G.; Hobbie, K.A.; Smith, B.W.; Anderson, K.A. PAH and OPAH Flux during the Deepwater Horizon Incident. Environ. Sci. Technol. 2016, 50, 7489–7749. [Google Scholar] [CrossRef]
- Forsberg, N.D.; Wilson, G.R.; Anderson, K.A. Determination of Parent and Substituted Polycyclic Aromatic Hydrocarbons in High-Fat Salmon Using a Modified Quechers Extraction, Dispersive Spe and Gc-Ms. J. Agric. Food Chem. 2011, 59, 8108–8116. [Google Scholar] [CrossRef] [PubMed]
- Øvrevik, J. Oxidative Potential Versus Biological Effects: A Review on the Relevance of Cell-Free/Abiotic Assays as Predictors of Toxicity from Airborne Particulate Matter. Int. J. Mol. Sci. 2019, 20, 4772. [Google Scholar] [CrossRef] [PubMed]
- Binelli, A.; Provini, A. POPs in edible clams from different Italian and European markets and possible human health risk. Mar. Pollut. Bull. 2003, 46, 879–886. [Google Scholar] [CrossRef]
- Radmacher, P.; Myers, S.; Looney, S.; Adamkin, D. A Pilot Study of Polycyclic Aromatic Hydrocarbons (Pah) in Maternal and Cord Blood Plasma. J. Investig. Med. 2008, 56, 476–477. [Google Scholar]
- Song, X.F.; Chen, Z.Y.; Zang, Z.J.; Zhang, Y.N.; Zeng, F.; Peng, Y.P.; Yang, C. Investigation of Polycyclic Aromatic Hydrocarbon Level in Blood and Semen Quality for Residents in Pearl River Delta Region in China. Environ. Int. 2013, 60, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Geier, M.C.; Chlebowski, A.C.; Truong, L.; Simonich, S.L.M.; Anderson, K.A.; Tanguay, R.L. Comparative developmental toxicity of a comprehensive suite of polycyclic aromatic hydrocarbons. Arch. Toxicol. 2018, 92, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Zama, A.M.; Uzumcu, M. Targeted Genome-Wide Methylation and Gene Expression Analyses Reveal Signaling Pathways Involved in Ovarian Dysfunction after Developmental EDC Exposure in Rats. Biol. Reprod. 2013, 88, 1–13. [Google Scholar] [CrossRef]
- Zeinomar, N.; Wu, H.-C.; Ma, X.; Herbstman, J.B.; Perera, F.P.; Miller, R.L.; Terry, M.B. Abstract C082: Prenatal exposure to polycyclic aromatic hydrocarbons and altered DNA methylation in breast cancer-related genes. Cancer Epidemiol. Biomark. Prev. 2020, 29, C082. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, T.; Xu, M.; Yu, H.; Ding, C.; Wang, Z.; Pan, X.; Li, Y.; Niu, Y.; Yan, R.; et al. Independent effect of main components in particulate matter on DNA methylation and DNA methyltransferase: A molecular epidemiology study. Environ. Int. 2019, 134, 105296. [Google Scholar] [CrossRef]
- Alegría-Torres, J.A.; Barretta, F.; Batres-Esquivel, L.E.; Carrizales-Yáñez, L.; Pérez-Maldonado, I.N.; Baccarelli, A.; Bertazzi, P.A. Epigenetic markers of exposure to polycyclic aromatic hydrocarbons in Mexican brickmakers: A pilot study. Chemosphere 2013, 91, 475–480. [Google Scholar] [CrossRef]
- Hou, L.; Zhu, Z.-Z.; Zhang, X.; Nordio, F.; Bonzini, M.; Schwartz, J.; Hoxha, M.; Dioni, L.; Marinelli, B.; Pegoraro, V.; et al. Airborne particulate matter and mitochondrial damage: A cross-sectional study. Environ. Health 2010, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, X.; Zhong, M.; Hotchkiss, I.P.; Lewandowski, R.P.; Wagner, J.G.; Bramble, L.A.; Yang, Y.; Wang, A.; Harkema, J.R.; et al. Ambient particulate air pollution induces oxidative stress and alterations of mitochondria and gene expression in brown and white adipose tissues. Part. Fibre Toxicol. 2011, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, C.; Xu, Z.; Tzan, K.; Zhong, M.; Wang, A.; Lippmann, M.; Chen, L.C.; Rajagopalan, S.; Sun, Q. Long-term Exposure to Ambient Fine Particulate Pollution Induces Insulin Resistance and Mitochondrial Alteration in Adipose Tissue. Toxicol. Sci. 2011, 124, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, M.; Ovrevik, J.; Mollerup, S.; Asare, N.; Longhin, E.; Dahlman, H.J.; Camatini, M.; Holme, J.A. Airborne Urban Particles (Milan Winter-PM2.5) Cause Mitotic Arrest and Cell Death: Effects on DNA, Mitochondria, Ahr Binding and Spindle Organization. Mol. Mech. Mutagen. 2011, 713, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Bergamasco, A.; Culotta, L.; De Stefano, C.; Orecchio, S.; Sammartano, S.; Barreca, S. Composition, Distribution, and Sources of Polycyclic Aromatic Hydrocarbons in Sediments of the Gulf of Milazzo (Mediterranean Sea, Italy). Polycycl. Aromat. Compd. 2014, 34, 397–424. [Google Scholar] [CrossRef]
- Singh, V.K.; Patel, D.K.; Ram, S.; Mathur, N.; Siddiqui, M.K.J.; Behari, J.R. Blood Levels of Polycyclic Aromatic Hydrocarbons in Children of Lucknow, India. Arch. Environ. Contam. Toxicol. 2008, 54, 348–354. [Google Scholar] [CrossRef]
- Pleil, J.; Stiegel, M.; Sobus, J.; Tabucchi, S.; Ghio, A.; Madden, M. Cumulative exposure assessment for trace-level polycyclic aromatic hydrocarbons (PAHs) using human blood and plasma analysis. J. Chromatogr. B 2010, 878, 1753–1760. [Google Scholar] [CrossRef]
- Baccarelli, A.A.; Byun, H.-M. Platelet mitochondrial DNA methylation: A potential new marker of cardiovascular disease. Clin. Epigenet. 2015, 7, 44. [Google Scholar] [CrossRef]
- Jiao, H.; Rui, X.; Wu, S.; Bai, Z.; Zhuang, X. Polycyclic Aromatic Hydrocarbons in the Dagang Oilfield (China): Distribution, Sources, and Risk Assessment. Int. J. Environ. Res. Public Health 2015, 12, 5775–5791. [Google Scholar] [CrossRef]
- Chandru, K.; Zakaria, M.P.; Anita, S.; Shahbazi, A.; Sakari, M.; Bahry, P.S.; Mohamed, C.A.R. Characterization of alkanes, hopanes, and polycyclic aromatic hydrocarbons (PAHs) in tar-balls collected from the East Coast of Peninsular Malaysia. Mar. Pollut. Bull. 2008, 56, 950–962. [Google Scholar] [CrossRef]
- Yin, S.; Tang, M.; Chen, F.; Li, T.; Liu, W. Environmental exposure to polycyclic aromatic hydrocarbons (PAHs): The correlation with and impact on reproductive hormones in umbilical cord serum. Environ. Pollut. 2017, 220, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhang, L.; Mesaros, C.; Zhang, S.; Blaha, M.A.; Blair, I.A.; Penning, T.M. Metabolism of a Representative Oxygenated Polycyclic Aromatic Hydrocarbon (Pah) Phenanthrene-9,10-Quinone in Human Hepatoma (Hepg2) Cells. Chem. Res. Toxicol. 2014, 27, 852–863. [Google Scholar] [CrossRef] [PubMed]
- Koukoulakis, K.; Kanellopoulos, P.; Chrysochou, E.; Koukoulas, V.; Minaidis, M.; Maropoulos, G.; Nikoleli, G.-P.; Bakeas, E. Leukemia and PAHs levels in human blood serum: Preliminary results from an adult cohort in Greece. Atmospheric Pollut. Res. 2020, 11, 1552–1565. [Google Scholar] [CrossRef]
- Barreca, S.; Bastone, S.; Caponetti, E.; Martino, D.F.C.; Orecchio, S. Determination of selected polyaromatic hydrocarbons by gas chromatography–mass spectrometry for the analysis of wood to establish the cause of sinking of an old vessel (Scauri wreck) by fire. Microchem. J. 2014, 117, 116–121. [Google Scholar] [CrossRef]
- Barreca, S.; Mazzola, A.; Orecchio, S.; Tuzzolino, N. Polychlorinated Biphenyls in Sediments from Sicilian Coastal Area (Scoglitti) Using Automated Soxhlet, Gc-Ms, and Principal Component Analysis. Polycycl. Aromat. Compd. 2014, 34, 237–262. [Google Scholar] [CrossRef]
- Bandowe, B.A.M.; Shukurov, N.; Leimer, S.; Kersten, M.; Steinberger, Y.; Wilcke, W. Polycyclic aromatic hydrocarbons (PAHs) in soils of an industrial area in semi-arid Uzbekistan: Spatial distribution, relationship with trace metals and risk assessment. Environ. Geochem. Health 2021, 43, 4847–4861. [Google Scholar] [CrossRef]
- Wang, W.; Jariyasopit, N.; Schrlau, J.; Jia, Y.; Tao, S.; Yu, T.-W.; Dashwood, R.H.; Zhang, W.; Wang, X.; Simonich, S.L.M. Concentration and Photochemistry of PAHs, NPAHs, and OPAHs and Toxicity of PM2.5 during the Beijing Olympic Games. Environ. Sci. Technol. 2011, 45, 6887–6895. [Google Scholar] [CrossRef]
- Yun, Y.; Zhang, Y.; Li, G.; Chen, S.; Sang, N. Embryonic exposure to oxy-polycyclic aromatic hydrocarbon interfere with pancreatic β-cell development in zebrafish via altering DNA methylation and gene expression. Sci. Total. Environ. 2019, 660, 1602–1609. [Google Scholar] [CrossRef]
- Bordoni, L.; Smerilli, V.; Nasuti, C.; Gabbianelli, R. Mitochondrial DNA methylation and copy number predict body composition in a young female population. J. Transl. Med. 2019, 17, 399. [Google Scholar] [CrossRef]
- Vos, S.; Nawrot, T.S.; Martens, D.S.; Byun, H.-M.; Janssen, B.G. Mitochondrial DNA methylation in placental tissue: A proof of concept study by means of prenatal environmental stressors. Epigenetics 2021, 16, 121–131. [Google Scholar] [CrossRef]
- Breton, C.V.; Song, A.Y.; Xiao, J.; Kim, S.-J.; Mehta, H.H.; Wan, J.; Yen, K.; Sioutas, C.; Lurmann, F.; Xue, S.; et al. Effects of air pollution on mitochondrial function, mitochondrial DNA methylation, and mitochondrial peptide expression. Mitochondrion 2019, 46, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Graff, C.; Bui, T.-H.; Larsson, N.-G. Mitochondrial diseases. Best Pract. Res. Clin. Obstet. Gynaecol. 2002, 16, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Valinluck, V.; Tsai, H.H.; Rogstad, D.K.; Burdzy, A.; Bird, A.; Sowers, L.C. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res. 2004, 32, 4100–4108. [Google Scholar] [CrossRef]
- Berdanier, C.D. Linking Mitochondrial Function to Diabetes Mellitus: An Animal’s Tale. Am. J. Physiol. Cell Physiol. 2007, 293, C830–C836. [Google Scholar] [CrossRef] [PubMed]
- Schrauwen-Hinderling, V.B.; Kooi, M.E.; Schrauwen, P. Mitochondrial Function and Diabetes: Consequences for Skeletal and Cardiac Muscle Metabolism. Antioxid. Redox Signal. 2016, 24, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Pecina, P.; Houstkova, H.; Hansikova, H.; Zeman, J.; Houstek, J. Genetic Defects of Cytochrome C Oxidase Assembly. Physiol. Res. 2004, 53, S213–S223. [Google Scholar] [CrossRef]
- Sun, X.; Vaghjiani, V.; Jayasekara, W.S.N.; Cain, J.E.; John, J.C.S. The degree of mitochondrial DNA methylation in tumor models of glioblastoma and osteosarcoma. Clin. Epigenetics 2018, 10, 157. [Google Scholar] [CrossRef]
- Dzitoyeva, S.; Chen, H.; Manev, H. Effect of aging on 5-hydroxymethylcytosine in brain mitochondria. Neurobiol. Aging 2012, 33, 2881–2891. [Google Scholar] [CrossRef]
- Pirola, C.J.; Gianotti, T.F.; Burgueño, A.L.; Rey-Funes, M.; Loidl, C.F.; Mallardi, P.; Martino, J.S.; Castaño, G.O.; Sookoian, S. Epigenetic modification of liver mitochondrial DNA is associated with histological severity of nonalcoholic fatty liver disease. Gut 2013, 62, 1356–1363. [Google Scholar] [CrossRef]
- Bauereisen, E.; Hauck, G.; Jacob, R.; Peiper, U. Enddiastolische Druck-Volumen-Relationen Und Arbeitsdiagramme Des Intakten Herzens Im Natürlichen Kreislauf in Abhängigkeit Von Herzfrequenz, Adrenalin Wirkung Und Vagusreiz. Pflüger’s Arch. Für Die Gesamte Physiol. Des Menschen Und Der Tiere 1964, 281, 216–230. [Google Scholar] [CrossRef]
- Byun, H.-M.; Panni, T.; Motta, V.; Hou, L.; Nordio, F.; Apostoli, P.; Bertazzi, P.A.; Baccarelli, A.A. Effects of airborne pollutants on mitochondrial DNA Methylation. Part. Fibre Toxicol. 2013, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-C.; Song, Y.-S.; Wang, H.; Zhang, J.; Yu, S.-F.; Gu, Y.-E.; Chen, T.; Wang, Y.; Shen, H.-Q.; Jia, G. Oxidative DNA damage and global DNA hypomethylation are related to folate deficiency in chromate manufacturing workers. J. Hazard. Mater. 2012, 213–214, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Salles, T.; Mendez, M.A.; Pessoa, V.; Guxens, M.; Aguilera, I.; Kogevinas, M.; Sunyer, J. Smoking during pregnancy is associated with higher dietary intake of polycyclic aromatic hydrocarbons and poor diet quality. Public Health Nutr. 2010, 13, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, L.; Zhou, M.; Li, J.; Liu, Q.; Wang, B.; Deng, K.; Zhou, P.; Wu, Y. Polycyclic aromatic hydrocarbons in the Chinese diet: Contamination characteristics, indicator screening, and health risk assessment. Food Addit. Contam. Part A 2023, 40, 625–640. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene Name | Primer | Sequence |
|---|---|---|
| MT-COX1 | Forward primer (5′ to 3′) | TATTAATTGGTTTTTTAGGGTTTAT |
| Reverse biotin primer (5′ to 3′) | CAACAAATCATTTCATATTACTTCC | |
| Sequencing primer (5′ to 3′) | TATTTATAGTAGGAAT | |
| MT-COX2 | Forward primer (5′ to 3′) | TTTATGAGTTGTTTTTATATTAGGTTTAAA |
| Reverse biotin primer (5′ to 3′) | ACTCCACAAATTTCAAAACATTAAC | |
| Sequencing primer (5′ to 3′) | TAAAAATAGATGTAAT | |
| MT-COX3 | Forward primer (5′ to 3′) | TATATTATTTGTTTAAAAAGGTTTT |
| Reverse biotin primer (5′ to 3′) | AATAAAAAACTCAAAAAAATCCTAC | |
| Sequencing primer (5′ to 3′) | TATATTATTTGTTTAAAAAGGTTTT |
| PAHs | Pre-Workshift | Post-Workshift | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Mean | Median (P25–P75) | Detection Frequency | Mean | Median, (P25–P75) | Detection Frequency | ||
| Nap | 0 | 0 | 0% | 1.8 | 1.7 (1.4, 2.1) | 44.4% | / |
| Flo | 4.1 | 3.9 (2.6, 5.7) | 94.4% | 6.6 | 6.0 (3.7, 9.0) | 83.3% | 0.0304 * |
| Phe | 14.6 | 13.3 (11.3, 19.2) | 100% | 23.4 | 22.1 (13.4, 29.1) | 83.3% | 0.0077 * |
| Ant | 1.7 | 1.6 (1.5, 2.0) | 44.4% | 2.7 | 2.9 (1.9, 3.2) | 55.5% | 0.0013 * |
| Flu | 3.3 | 3.5 (2.2, 3.8) | 94.4% | 6.0 | 5.2 (4.2, 7.7) | 72.2% | 0.0430 * |
| Pyr | 7.7 | 6.1 (3.97, 7.8) | 100% | 8.2 | 7.6 (3.6, 11.6) | 94.4% | 0.9617 |
| Nap-1 | 1.3 | 1.3 (1.3, 1.3) | 5.5% | 2.0 | 1.8 (1.4, 2.3) | 33.3% | / |
| Nap-2 | 1.4 | 1.1 (1.1, 1.6) | 44.4% | 2.5 | 2.2 (1.7, 2.9) | 61.1% | 0.2151 |
| Dibf | 7.2 | 5.4 (3.3, 8.0) | 94.4% | 8.7 | 6.0 (4.9, 14.1) | 83.3% | 0.8130 |
| The total of all components | 41.3 | 35.2 (24.0, 48.5) | / | 61.8 | 44.0 (19.7, 66.8) | / | 0.2948 |
| Variables | MT−COX1 | MT−COX2 | MT−COX3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| β | SD | p-Value | β | SD | p-Value | β | SD | p-Value | |
| Nap−2 | 0.688 | 0.555 | 0.243 | 0.680 | 0.817 | 0.424 | 0.089 | 0.142 | 0.213 |
| Dibf | 0.008 | 0.095 | 0.934 | −0.255 | 0.147 | 0.105 | 0.058 | 0.028 | 0.545 |
| Flo | 0.126 | 0.185 | 0.508 | −0.083 | 0.251 | 0.745 | 0.115 | 0.042 | 0.016 * |
| Phe | 0.088 | 0.063 | 0.188 | 0.091 | 0.092 | 0.337 | 0.036 | 0.015 | 0.037 * |
| Ant | 0.831 | 0.105 | 0.000 * | 1.893 | 1.364 | 0.199 | 0.192 | 0.232 | 0.430 |
| Flu | 0.270 | 0.208 | 0.215 | 0.365 | 0.342 | 0.303 | 0.064 | 0.070 | 0.372 |
| Pyr | −0.022 | 0.067 | 0.744 | 0.046 | 0.122 | 0.711 | 0.018 | 0.023 | 0.457 |
| The total of all components | 0.011 | 0.020 | 0.590 | 0.015 | 0.027 | 0.606 | 0.006 | 0.005 | 0.245 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Li, W.; Li, Y.; Zhao, L.; Li, C.; Wang, L.; Fang, J.; Song, S.; Ji, Y.; Fang, T.; et al. The Levels of Polycyclic Aromatic Hydrocarbons and Their Derivatives in Plasma and Their Effect on Mitochondrial DNA Methylation in the Oilfield Workers. Toxics 2023, 11, 466. https://doi.org/10.3390/toxics11050466
Jia Y, Li W, Li Y, Zhao L, Li C, Wang L, Fang J, Song S, Ji Y, Fang T, et al. The Levels of Polycyclic Aromatic Hydrocarbons and Their Derivatives in Plasma and Their Effect on Mitochondrial DNA Methylation in the Oilfield Workers. Toxics. 2023; 11(5):466. https://doi.org/10.3390/toxics11050466
Chicago/Turabian StyleJia, Yaning, Weixia Li, Yanlin Li, Lei Zhao, Chenguang Li, Lei Wang, Junkai Fang, Shanjun Song, Yaqin Ji, Tao Fang, and et al. 2023. "The Levels of Polycyclic Aromatic Hydrocarbons and Their Derivatives in Plasma and Their Effect on Mitochondrial DNA Methylation in the Oilfield Workers" Toxics 11, no. 5: 466. https://doi.org/10.3390/toxics11050466
APA StyleJia, Y., Li, W., Li, Y., Zhao, L., Li, C., Wang, L., Fang, J., Song, S., Ji, Y., Fang, T., Zhang, J., Guo, L., & Li, P. (2023). The Levels of Polycyclic Aromatic Hydrocarbons and Their Derivatives in Plasma and Their Effect on Mitochondrial DNA Methylation in the Oilfield Workers. Toxics, 11(5), 466. https://doi.org/10.3390/toxics11050466