
MPTP-Treated Zebrafish Recapitulate ‘Late-Stage’ Parkinson’s-like Cognitive Decline

 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Drug Treatment
2.3. Experimental Design and Acute MPTP Toxicity (LD50) Assay
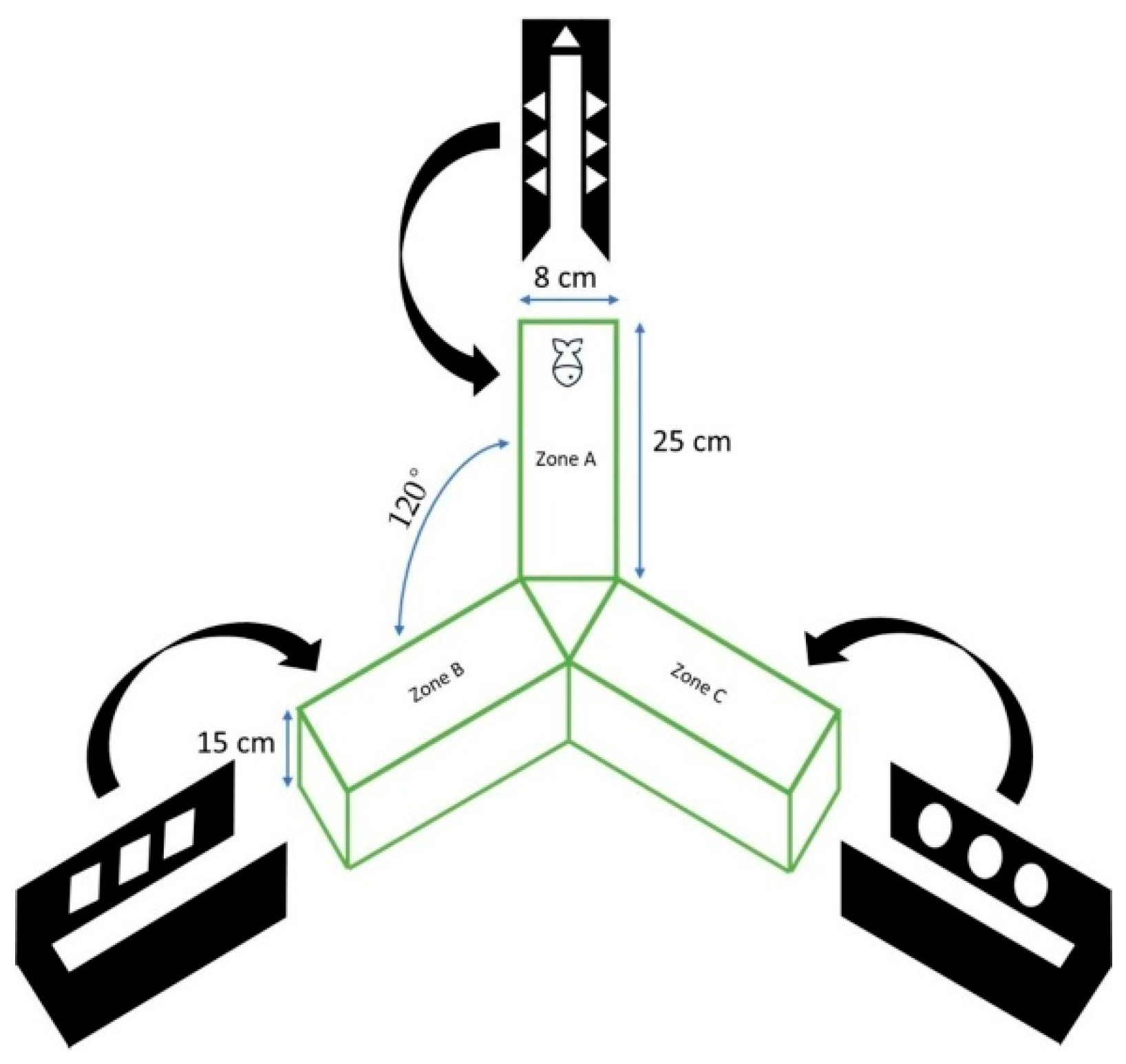
2.4. Spontaneous Alternation and Inhibitory Avoidance Assays
2.5. AI-Based Analysis
2.6. Data Analysis
3. Results
3.1. Acute MPTP Toxicity in Adult Zebrafish (LD50 Assay)
3.2. Cognitive Behaviors
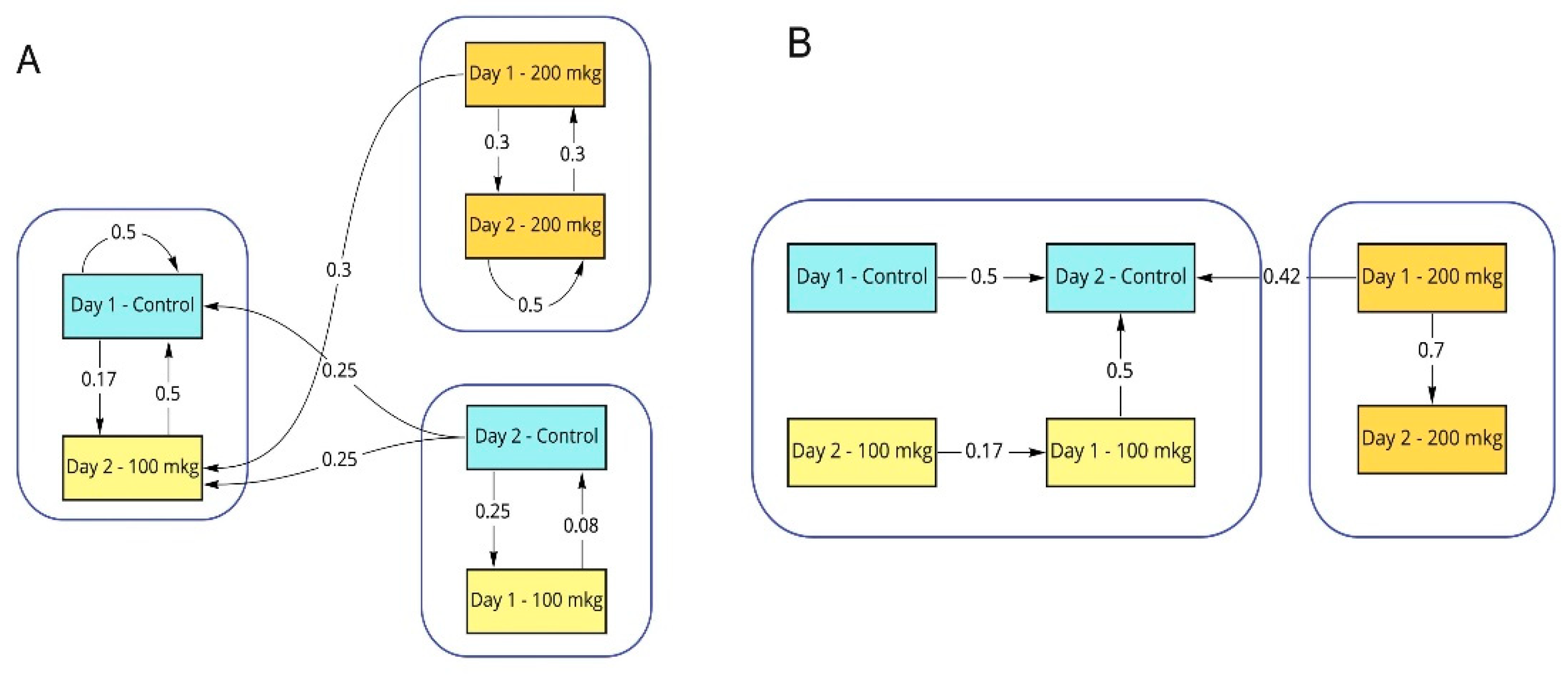
3.3. AI-Based Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, A.; Gilbert, R.M. Epidemiology of Parkinson disease. Neurol. Clin. 2016, 34, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Rodriguez, M.; Lanciego, J.L.; Artieda, J.; Gonzalo, N.; Olanow, C.W. Pathophysiology of the basal ganglia in Parkinson’s disease. Trends Neurosci. 2000, 23, S8–S19. [Google Scholar] [CrossRef]
- Mao, Q.; Qin, W.Z.; Zhang, A.; Ye, N. Recent advances in dopaminergic strategies for the treatment of Parkinson’s disease. Acta Pharmacol. Sin. 2020, 41, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Secker, D.; Brown, R. Cognitive behavioural therapy (CBT) for carers of patients with Parkinson’s disease: A preliminary randomised controlled trial. J. Neurol. Neurosurg. Psychiatry 2005, 76, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Ruiz, B.; Junque, C.; Marti, M.J.; Valldeoriola, F.; Tolosa, E. Cognitive changes in Parkinson’s disease patients with visual hallucinations. Dement. Geriatr. Cogn. Disord. 2007, 23, 281–288. [Google Scholar] [CrossRef]
- Przedborski, S.; Jackson-Lewis, V.; Yokoyama, R.; Shibata, T.; Dawson, V.L.; Dawson, T.M. Role of neuronal nitric oxide in 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP)-induced dopaminergic neurotoxicity. Proc. Natl. Acad. Sci. USA 1996, 93, 4565–4571. [Google Scholar] [CrossRef] [Green Version]
- Przedborski, S.; Jackson-Lewis, V. Mechanisms of MPTP toxicity. Mov. Disord. Off. J. Mov. Disord. Soc. 1998, 13, 35–38. [Google Scholar]
- Przedborski, S.; Vila, M. MPTP: A review of its mechanisms of neurotoxicity. Clin. Neurosci. Res. 2001, 1, 407–418. [Google Scholar] [CrossRef]
- Langston, J.W.; Forno, L.S.; Rebert, C.S.; Irwin, I. Selective nigral toxicity after systemic administration of 1-methyl-4-phenyl-1, 2, 5, 6-tetrahydropyrine (MPTP) in the squirrel monkey. Brain Res. 1984, 292, 390–394. [Google Scholar] [CrossRef]
- Ballard, P.A.; Tetrud, J.W.; Langston, J.W. Permanent human parkinsonism due to 1-methy 1-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP): Seven cases. Neurology 1985, 35, 949. [Google Scholar] [CrossRef]
- Beal, M.F. Experimental models of Parkinson’s disease. Nat. Rev. Neurosci. 2001, 2, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Sallinen, V.; Torkko, V.; Sundvik, M.; Reenilä, I.; Khrustalyov, D.; Kaslin, J.; Panula, P. MPTP and MPP+ target specific aminergic cell populations in larval zebrafish. J. Neurochem. 2009, 108, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Nicklas, W.J.; Vyas, I.; Heikkila, R.E. Inhibition of NADH-linked oxidation in brain mitochondria by 1-methyl-4-phenyl-pyridine, a metabolite of the neurotoxin, 1-methyl-4-phenyl-1, 2, 5, 6-tetrahydropyridine. Life Sci. 1985, 36, 2503–2508. [Google Scholar] [CrossRef]
- Berardo, A.; Musumeci, O.; Toscano, A. Cardiological manifestations of mitochondrial respiratory chain disorders. Acta Myol. 2011, 30, 9. [Google Scholar]
- Schmidt, D.E.; Ebert, M.H.; Lynn, J.C.; Whetsell, W.O. Attenuation of 1-methyl-4-phenylpyridinium (MPP+) neurotoxicity by deprenyl in organotypic canine substantia nigra cultures. J. Neural Transm. 1997, 104, 875–885. [Google Scholar] [CrossRef]
- Meredith, G.E.; Rademacher, D.J. MPTP mouse models of Parkinson’s disease: An update. J. Parkinson’s Dis. 2011, 1, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Razali, K.; Othman, N.; Nasir, M.H.M.; Doolaanea, A.A.; Kumar, J.; Ibrahim, W.N.; Ibrahim, N.M.; Mohamed, W.M. The Promise of the Zebrafish Model for Parkinson’s Disease: Today’s Science and Tomorrow’s Treatment. Front. Genet. 2021, 12, 655550. [Google Scholar] [CrossRef]
- Parng, C.; Roy, N.M.; Ton, C.; Lin, Y.; McGrath, P. Neurotoxicity assessment using zebrafish. J. Pharmacol. Toxicol. Methods 2007, 55, 103–112. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. The teleostean (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum). Brain Res. 2001, 889, 316–330. [Google Scholar] [CrossRef]
- Jiang, P.E.; Lang, Q.H.; Yu, Q.Y.; Tang, X.Y.; Liu, Q.Q.; Li, X.Y.; Feng, X.Z. Behavioral Assessments of Spontaneous Locomotion in a Murine MPTP-induced Parkinson’s Disease Model. J. Vis. Exp. 2019, 143, e58653. [Google Scholar] [CrossRef] [Green Version]
- Jenner, P. The MPTP-treated primate as a model of motor complications in PD: Primate model of motor complications. Neurology 2003, 61 (Suppl. 3), S4–S11. [Google Scholar] [CrossRef]
- Selvaraj, V.; Venkatasubramanian, H.; Ilango, K.; Santhakumar, K. A simple method to study motor and non-motor behaviors in adult zebrafish. J. Neurosci. Methods 2019, 320, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Sallinen, V.; Kolehmainen, J.; Priyadarshini, M.; Toleikyte, G.; Chen, Y.C.; Panula, P. Dopaminergic cell damage and vulnerability to MPTP in Pink1 knockdown zebrafish. Neurobiol. Dis. 2010, 40, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R. The neurobiological basis of spontaneous alternation. Neurosci. Biobehav. Rev. 2002, 26, 91–104. [Google Scholar] [CrossRef]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. A Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- de Abreu, M.S.; Giacomini, A.C.; Demin, K.A.; Petersen, E.V.; Kalueff, A.V. On the value of zebrafish outbred strains in neurobehavioral research. Lab Anim. 2021, 51, 5–6. [Google Scholar] [CrossRef]
- Serikuly, N.; Alpyshov, E.T.; Wang, D.M.; Wang, J.T.; Yang, L.E.; Hu, G.J.; Yan, D.N.; Demin, K.A.; Kolesnikova, T.O.; Galstyan, D.; et al. Effects of acute and chronic arecoline in adult zebrafish: Anxiolytic-like activity, elevated brain monoamines and the potential role of microglia. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2021, 104, 109977. [Google Scholar] [CrossRef]
- Randhawa, M.A. Calculation of LD50 values from the method of miller and Tainter, 1944. J. Ayub. Med. Coll. Abbottabad. 2009, 21, 184–185. [Google Scholar]
- Cognato, G.D.P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-Maze memory task in zebrafish (Danio rerio): The role of glutamatergic and cholinergic systems on the acquisition and consolidation periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef]
- Yamashita, R.; Nishio, M.; Do, R.K.G.; Togashi, K. Convolutional neural networks: An overview and application in radiology. Insights Into Imaging 2018, 9, 611–629. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, G.W. Convolutional neural networks as a model of the visual system: Past, present, and future. J. Cogn. Neurosci. 2020, 33, 2017–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Shi, J.; Li, Z.; Li, C.; Zhu, J.; Liu, S. Towards better analysis of deep convolutional neural networks. IEEE Trans. Vis. Comput. Graph. 2016, 23, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozhko, D.V.; Myrov, V.O.; Kolchanova, S.M.; Polovian, A.I.; Galumov, G.K.; Demin, K.A.; Zabegalov, K.N.; Strekalova, S.T.; de Abreu, M.S.; Petersen, E.V.; et al. Artificial intelligence-driven phenotyping of zebrafish psychoactive drug responses. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2022, 112, 110405. [Google Scholar] [CrossRef]
- Flach, P. Performance evaluation in machine learning: The good, the bad, the ugly, and the way forward. In Proceedings of the AAAI Conference on Artificial Intelligence, Honolulu, HI, USA, 24 January–1 February 2019. [Google Scholar]
- Que, X.; Checconi, F.; Petrini, F.; Gunnels, J.A. Scalable community detection with the louvain algorithm. In Proceedings of the 2015 IEEE International Parallel and Distributed Processing Symposium, Hyderabad, India, 25–29 May 2015. [Google Scholar]
- Anichtchik, O.V.; Kaslin, J.; Peitsaro, N.; Scheinin, M.; Panula, P. Neurochemical and behavioural changes in zebrafish Danio rerio after systemic administration of 6-hydroxydopamine and 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine. J. Neurochem. 2004, 88, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Bretaud, S.; Lee, S.; Guo, S. Sensitivity of zebrafish to environmental toxins implicated in Parkinson’s disease. Neurotox. Teratol. 2004, 26, 857–864. [Google Scholar] [CrossRef]
- Sarath Babu, N.; Murthy, C.L.N.; Kakara, S.; Sharma, R.; Brahmendra Swamy, C.V.; Idris, M.M. 1-Methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine induced Parkinson’s disease in zebrafish. Proteomics 2016, 16, 1407–1420. [Google Scholar] [CrossRef]
- Lal, P.; Tanabe, H.; Suster, M.L.; Ailani, D.; Kotani, Y.; Muto, A.; Itoh, M.; Iwasaki, M.; Wada, H.; Yaksi, E.; et al. Identification of a neuronal population in the telencephalon essential for fear conditioning in zebrafish. BMC Biol 2018, 16, 45. [Google Scholar] [CrossRef]
- Owen, A.M.; Iddon, J.L.; Hodges, J.R.; Summers, B.A.; Robbins, T.W. Spatial and non-spatial working memory at different stages of Parkinson’s disease. Neuropsychologia 1997, 35, 519–532. [Google Scholar] [CrossRef]
- Freedman, M.; Oscar-Berman, M. Selective delayed response deficits in Parkinson’s and Alzheimer’s disease. Arch. Neurol. 1986, 43, 886–890. [Google Scholar] [CrossRef]
- Cooper, J.A.; Sagar, H.J.; Jordan, N.; Harvey, N.S.; Sullivan, E. Cognitive impairment in early, untreated Parkinson’s disease and its relationship to motor disability. Brain 1991, 114 Pt 5, 2095–2122. [Google Scholar] [CrossRef]
- Schneider, J.S.; Kovelowski, C.J. II. Chronic exposure to low doses of MPTP. I. Cognitive deficits in motor asymptomatic monkeys. Brain Res. 1990, 519, 122–128. [Google Scholar] [CrossRef]
- Roeltgen, D.P.; Schneider, J.S. Chronic low-dose MPTP in nonhuman primates: A possible model for attention deficit disorder. J. Child Neurol. 1991, 6, S82–S89. [Google Scholar] [CrossRef] [PubMed]
- Decamp, E.; Tinker, J.P.; Schneider, J.S. Attentional cueing reverses deficits in spatial working memory task performance in chronic low dose MPTP-treated monkeys. Behav. Brain Res. 2004, 152, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, S.; Yabuki, Y.; Fukunaga, K. Reduced calcium/calmodulin-dependent protein kinase II activity in the hippocampus is associated with impaired cognitive function in MPTP-treated mice. J. Neurochem. 2012, 120, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Deguil, J.; Chavant, F.; Lafay-Chebassier, C.; Pérault-Pochat, M.C.; Fauconneau, B.; Pain, S. Neuroprotective Effect of PACAP on Translational Control Alteration and Cognitive Decline in MPTP Parkinsonian Mice. Neurotox. Res. 2010, 17, 142–155. [Google Scholar] [CrossRef]
- Kumar, P.; Kaundal, R.K.; More, S.; Sharma, S.S. Beneficial effects of pioglitazone on cognitive impairment in MPTP model of Parkinson’s disease. Behav. Brain Res. 2009, 197, 398–403. [Google Scholar] [CrossRef]
- Roeltgen, D.P.; Schneider, J.S. Task persistence and learning ability in normal and chronic low dose MPTP-treated monkeys. Behav. Brain Res. 1994, 60, 115–124. [Google Scholar] [CrossRef]
- Yan, J.; Liu, A.; Fan, H.; Qiao, L.; Wu, J.; Shen, M.; Lai, X.; Huang, J. Simvastatin improves behavioral disorders and hippocampal inflammatory reaction by NMDA-mediated anti-inflammatory function in MPTP-treated mice. Cell. Mol. Neurobiol. 2020, 40, 1155–1164. [Google Scholar] [CrossRef]
- Xu, Z.; Cheng, X.E. Zebrafish tracking using convolutional neural networks. Sci. Rep. 2017, 7, 42815. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.; Haurum, J.B.; Bengtson, S.H.; Moeslund, T.B. 3D-ZeF: A 3D Zebrafish Tracking Benchmark Dataset. In Proceedings of the 2020 IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR), Seattle, WA, USA, 13–19 June 2020. [Google Scholar]
- Wang, X.; Cheng, E.; Burnett, I.S.; Huang, Y.; Wlodkowic, D. Automatic multiple zebrafish larvae tracking in unconstrained microscopic video conditions. Sci. Rep. 2017, 7, 17596. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Peterson, R.T. Social behavioral profiling by unsupervised deep learning reveals a stimulative effect of dopamine D3 agonists on zebrafish sociality. bioRxiv 2021. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Datasets | Experiment 1 | Experiment 2A | Experiment 2B | Experiment 3A | Experiment 3B |
|---|---|---|---|---|---|
| Day 1—Control | Training and testing | Training and testing | Training | Testing | |
| Day 1—100 μg | |||||
| Day 1—200 μg | |||||
| Day 2—Control | Training and testing | Testing | Training | ||
| Day 2—100 μg | |||||
| Day 2—200 μg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashirzade, A.A.O.; Cheresiz, S.V.; Belova, A.S.; Drobkov, A.V.; Korotaeva, A.D.; Azizi-Arani, S.; Azimirad, A.; Odle, E.; Gild, E.-Y.V.; Ardashov, O.V.; et al. MPTP-Treated Zebrafish Recapitulate ‘Late-Stage’ Parkinson’s-like Cognitive Decline. Toxics 2022, 10, 69. https://doi.org/10.3390/toxics10020069
Bashirzade AAO, Cheresiz SV, Belova AS, Drobkov AV, Korotaeva AD, Azizi-Arani S, Azimirad A, Odle E, Gild E-YV, Ardashov OV, et al. MPTP-Treated Zebrafish Recapitulate ‘Late-Stage’ Parkinson’s-like Cognitive Decline. Toxics. 2022; 10(2):69. https://doi.org/10.3390/toxics10020069
Chicago/Turabian StyleBashirzade, Alim A. O., Sergey V. Cheresiz, Alisa S. Belova, Alexey V. Drobkov, Anastasiia D. Korotaeva, Soheil Azizi-Arani, Amirhossein Azimirad, Eric Odle, Emma-Yanina V. Gild, Oleg V. Ardashov, and et al. 2022. "MPTP-Treated Zebrafish Recapitulate ‘Late-Stage’ Parkinson’s-like Cognitive Decline" Toxics 10, no. 2: 69. https://doi.org/10.3390/toxics10020069
APA StyleBashirzade, A. A. O., Cheresiz, S. V., Belova, A. S., Drobkov, A. V., Korotaeva, A. D., Azizi-Arani, S., Azimirad, A., Odle, E., Gild, E.-Y. V., Ardashov, O. V., Volcho, K. P., Bozhko, D. V., Myrov, V. O., Kolchanova, S. M., Polovian, A. I., Galumov, G. K., Salakhutdinov, N. F., Amstislavskaya, T. G., & Kalueff, A. V. (2022). MPTP-Treated Zebrafish Recapitulate ‘Late-Stage’ Parkinson’s-like Cognitive Decline. Toxics, 10(2), 69. https://doi.org/10.3390/toxics10020069