
Differential Uptake and Translocation of Cadmium and Lead by Quinoa: A Multivariate Comparison of Physiological and Oxidative Stress Responses


Abstract
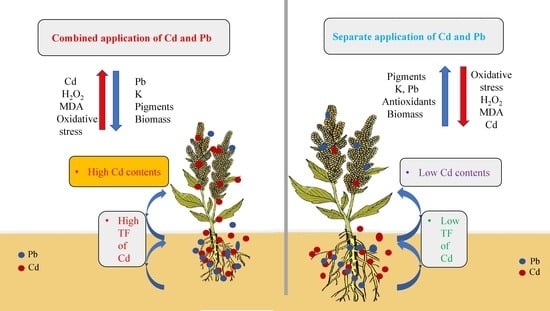
:
1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Treatment Application
2.2. Harvesting of Plants and Growth Measurement
2.3. Ionic Analysis of Plant Samples
2.4. Metal Tolerance and Translocation
2.5. Pigment Contents and Stomatal Conductance
2.6. Oxidative Stress Attributes
2.7. Enzymatic Activities
2.8. Statistical Analysis
3. Results
3.1. Plant Growth
3.2. Pigment Contents and Stomatal Conductance
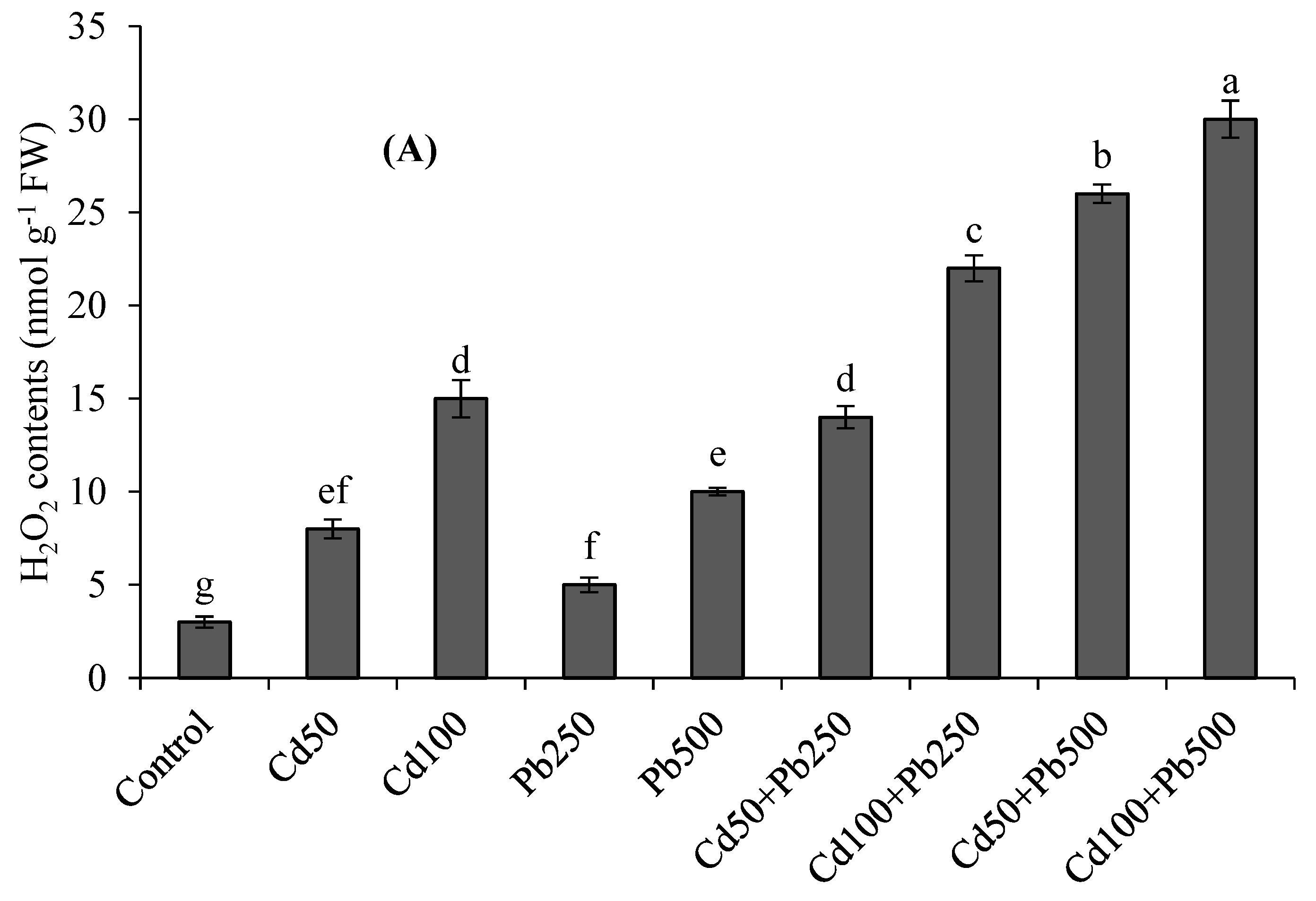
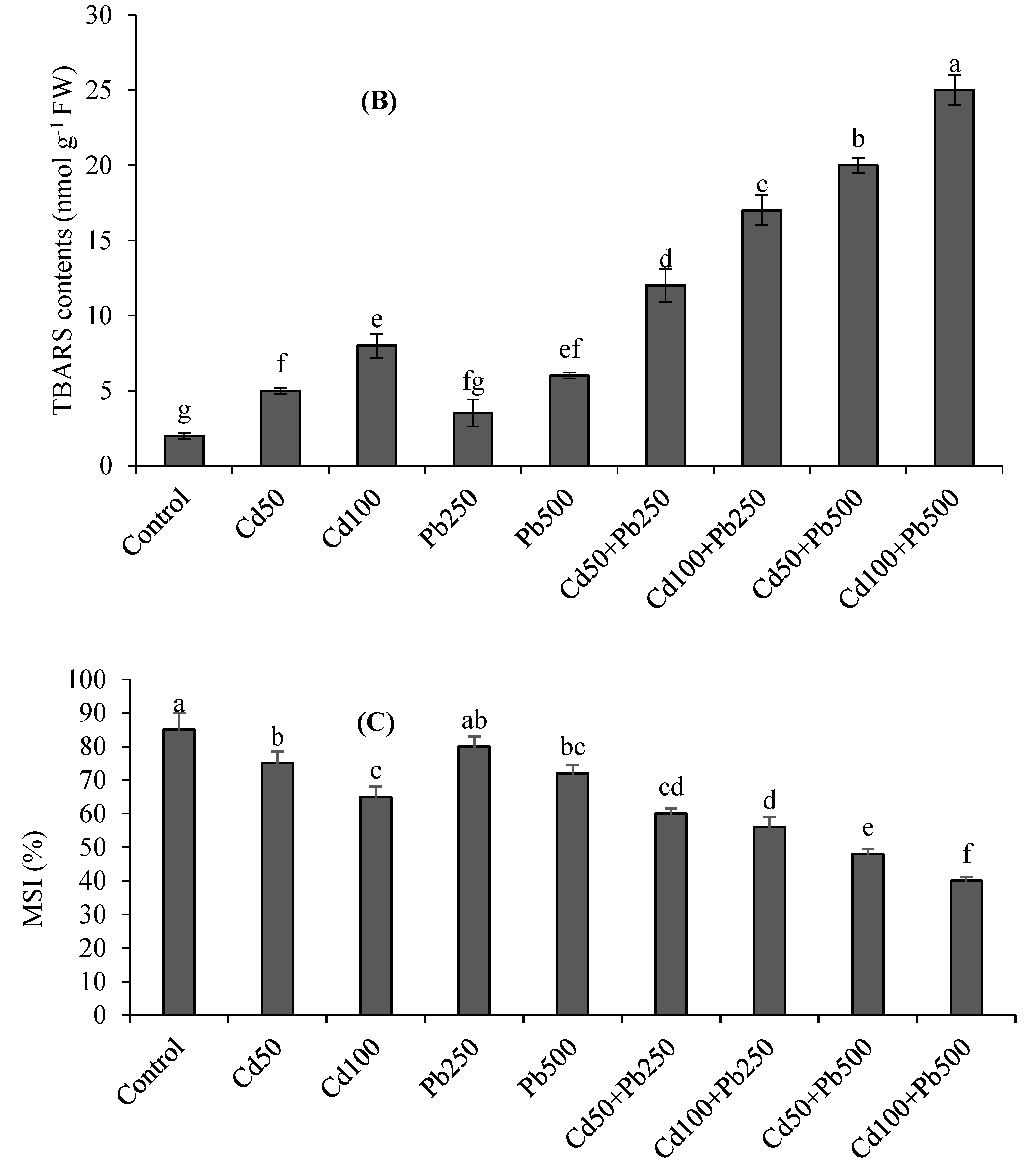
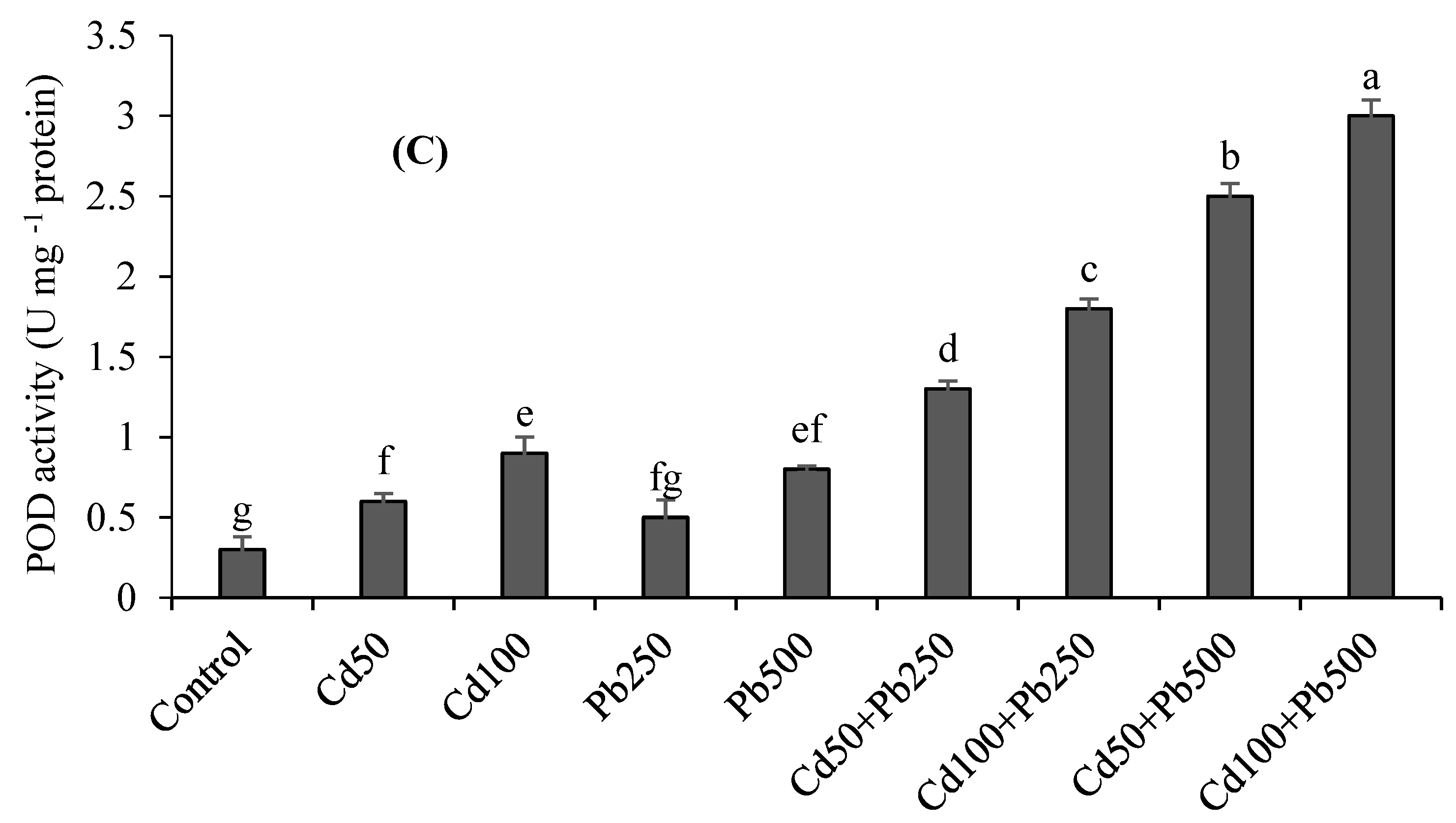
3.3. Oxidative Stress Attributes
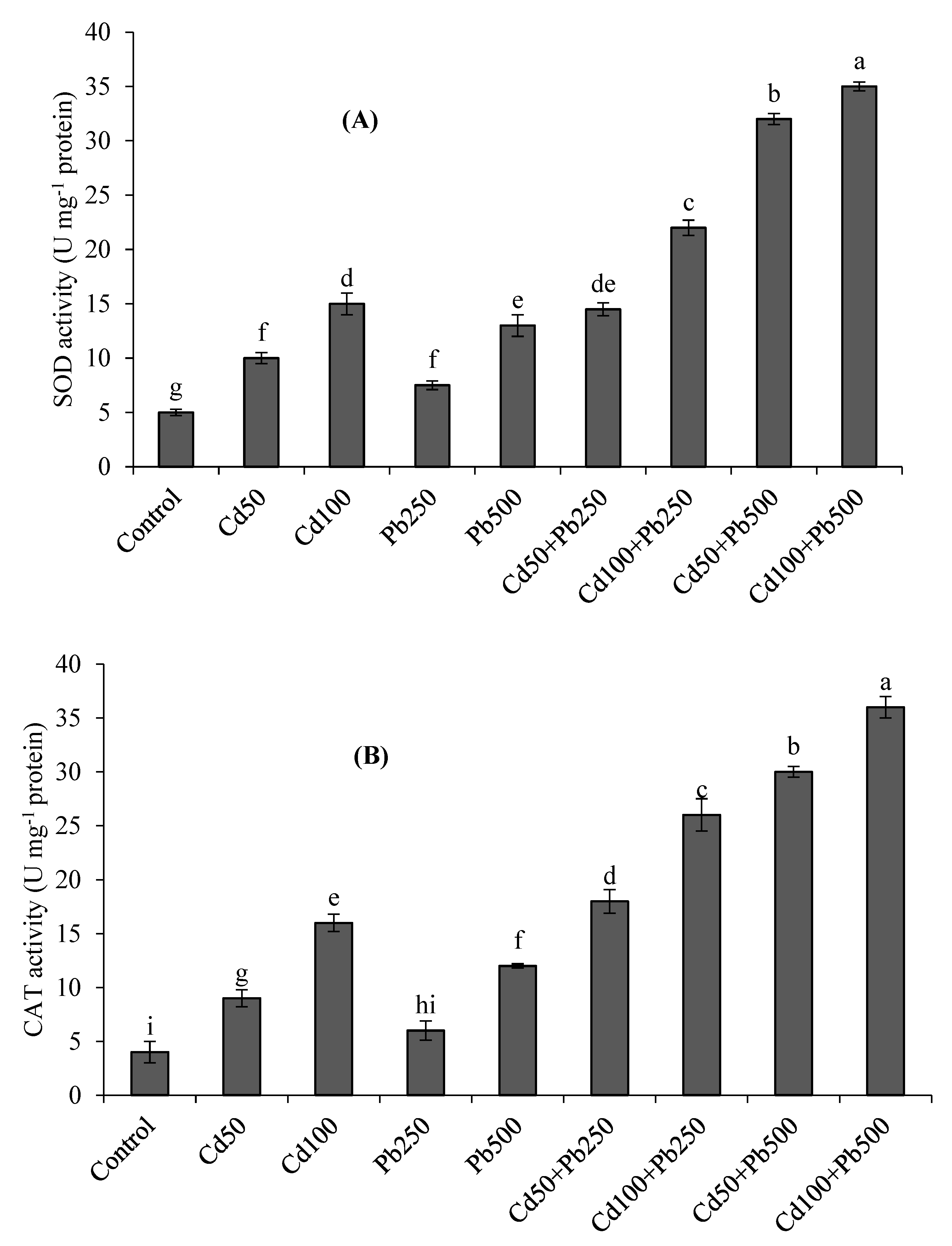
3.4. Antioxidant Enzymes
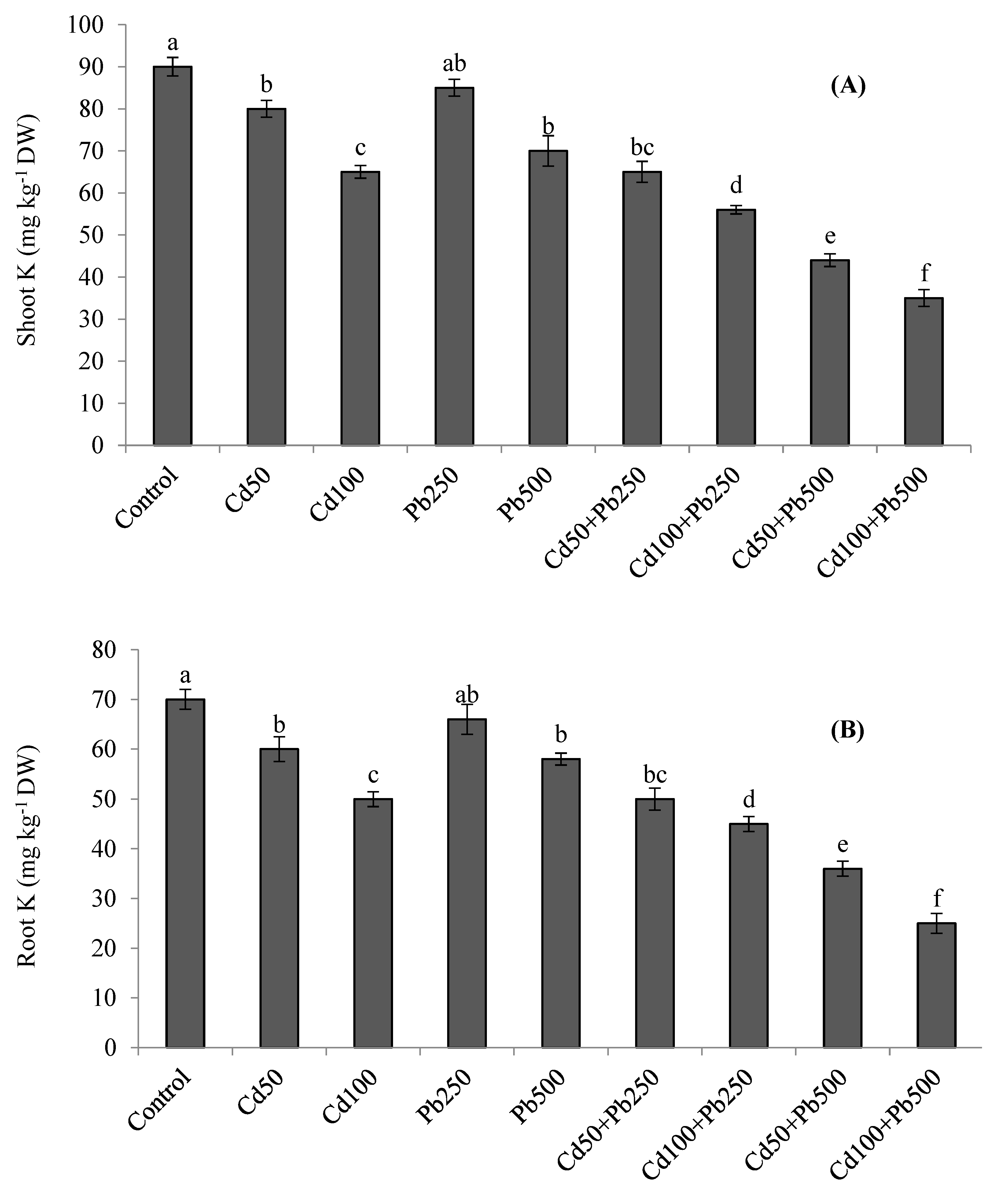
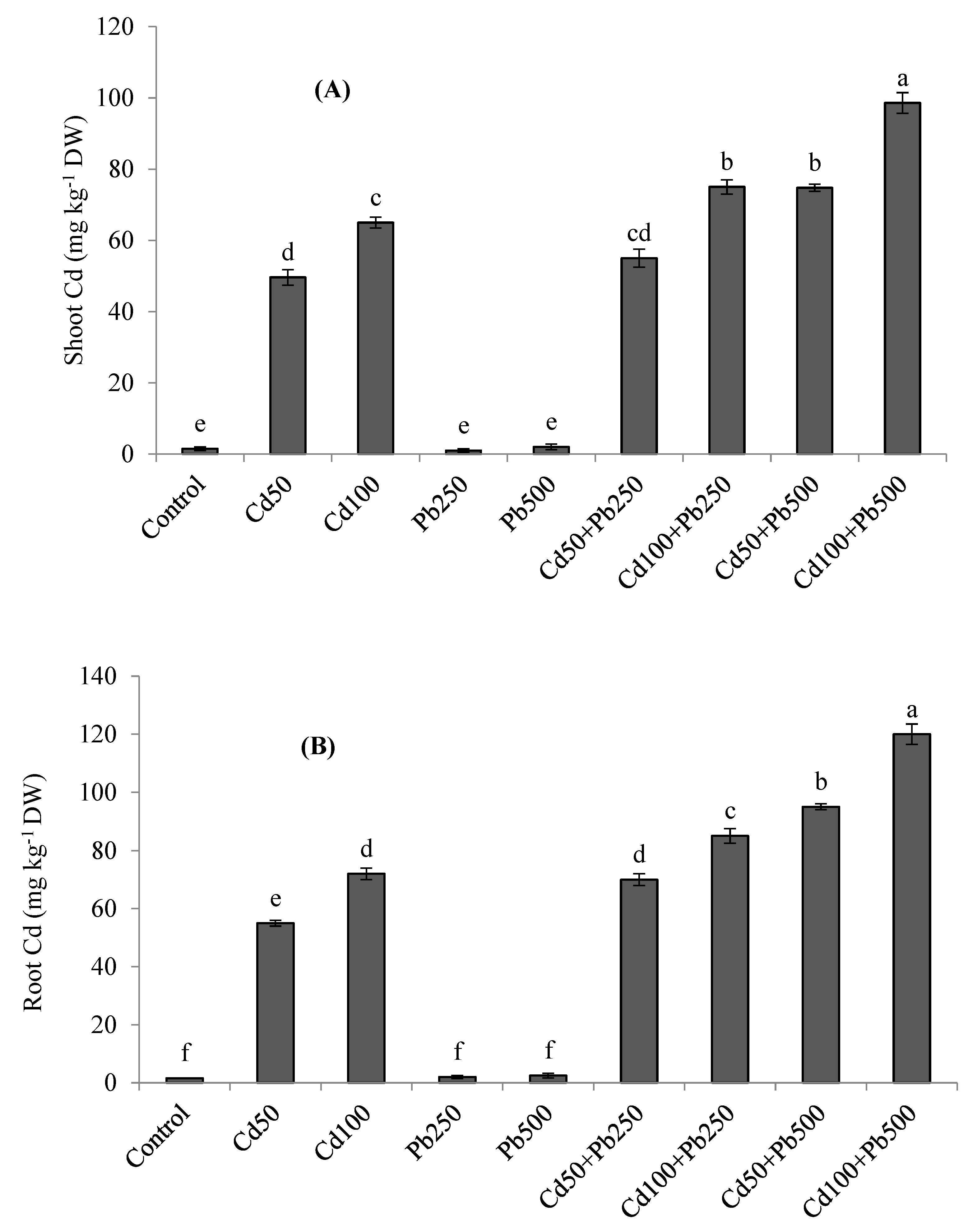
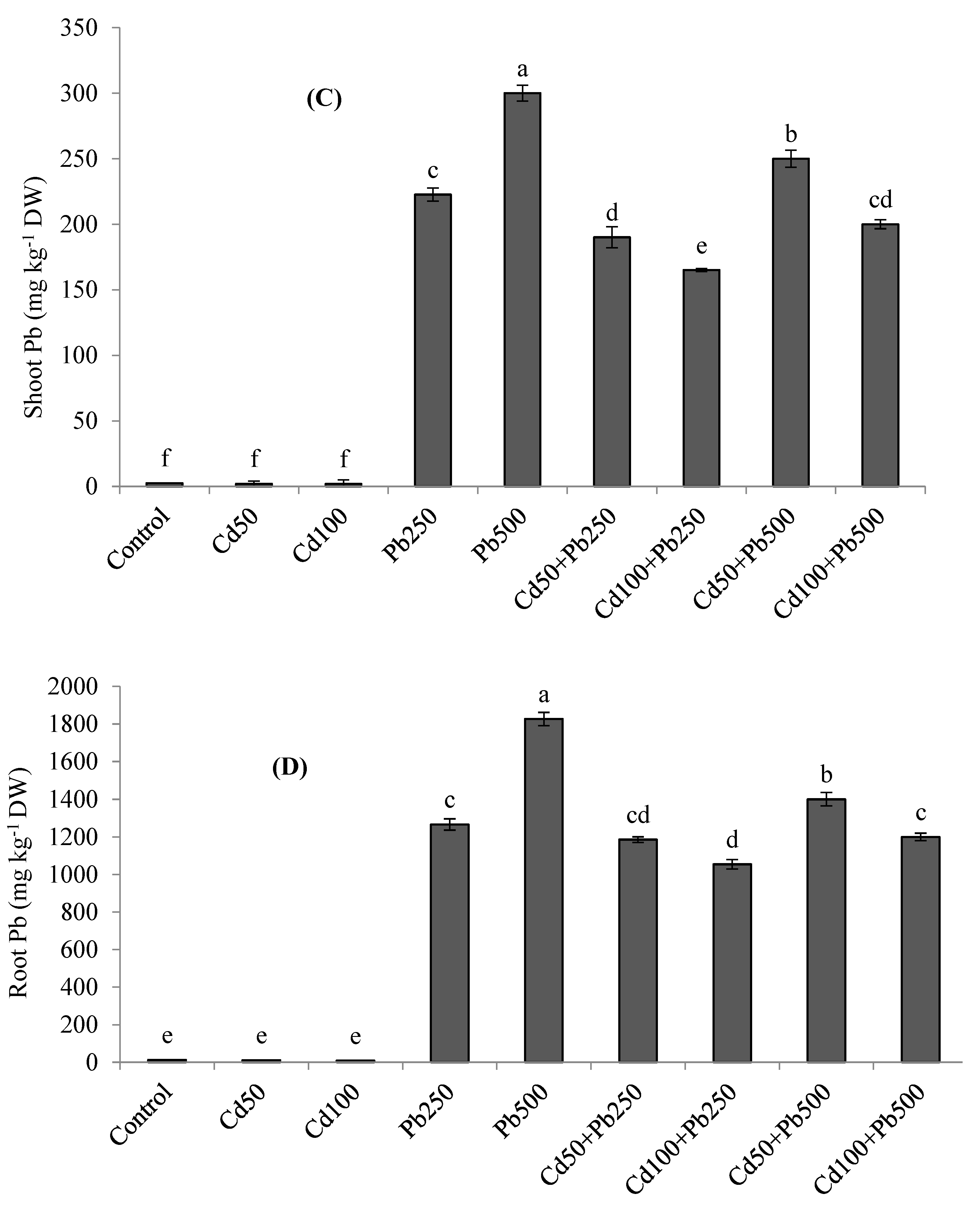
3.5. Potassium and Heavy Metals Accumulation
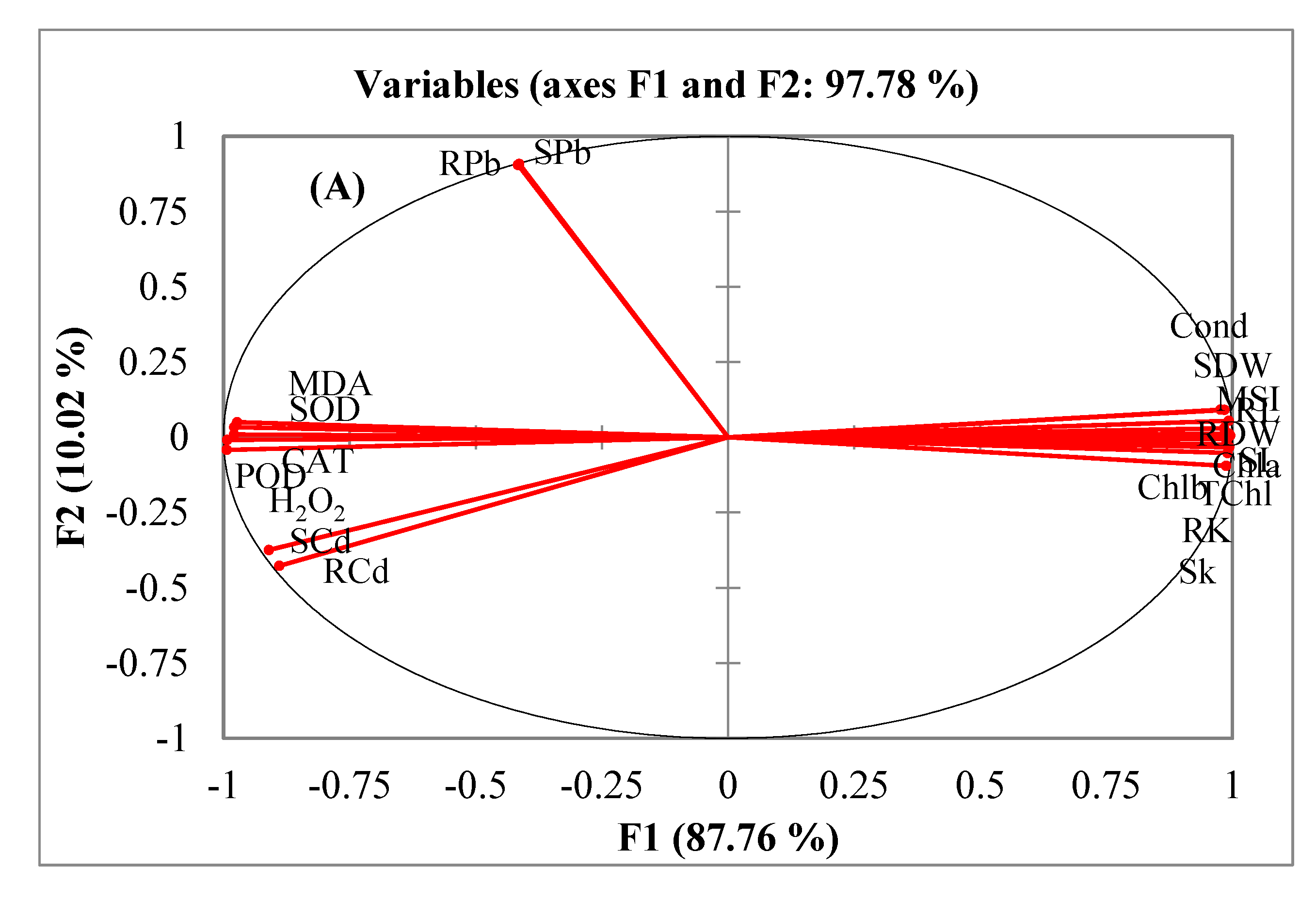
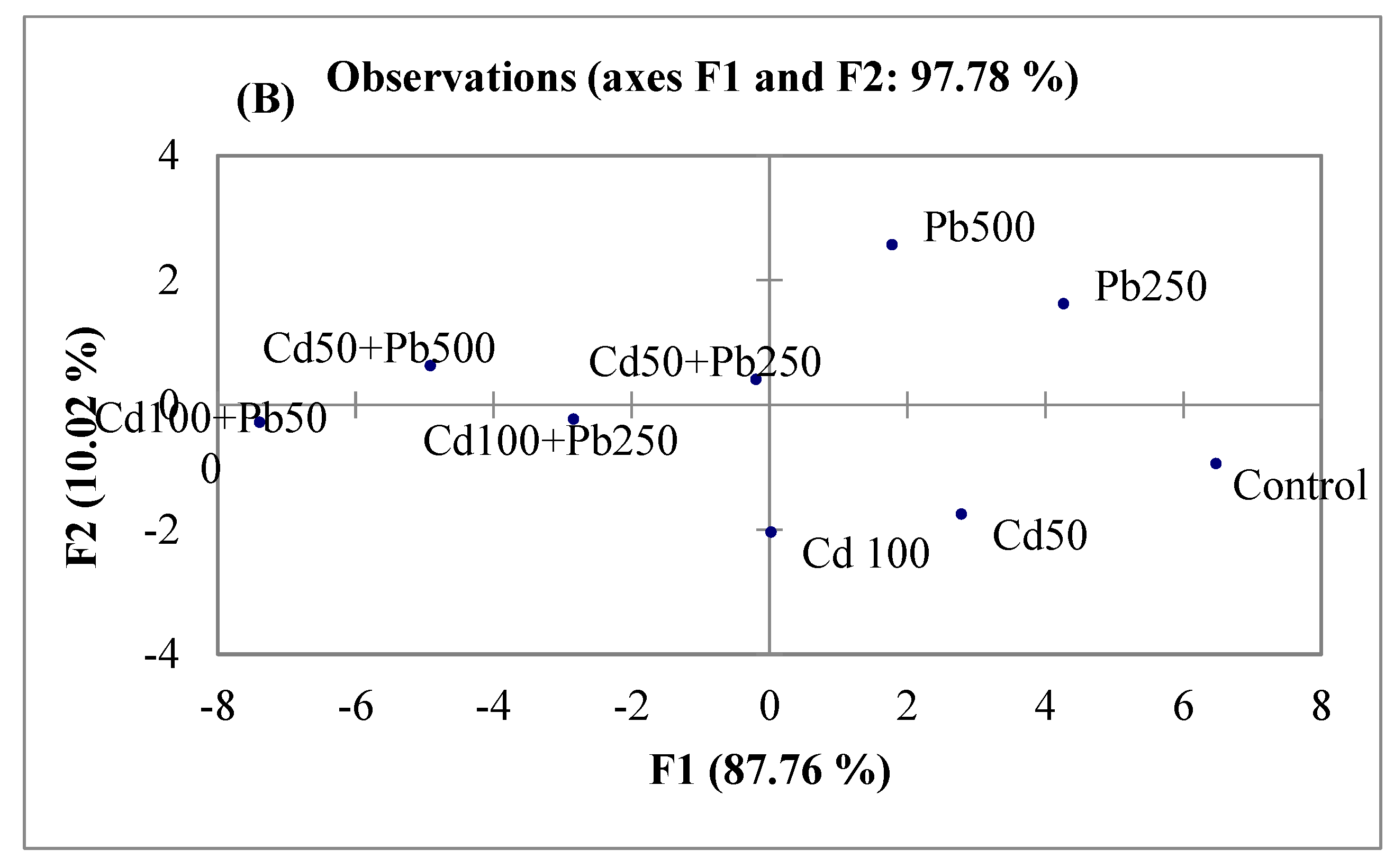
3.6. Multivariate Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qayyum, M.F.; Rehman, M.Z.U.; Ali, S.; Rizwan, M.; Naeem, A.; Maqsood, M.A.; Khalid, H.; Rinklebe, J.; Ok, Y.S. Residual effects of monoammonium phosphate, gypsum and elemental sulfur on cadmium phytoavailability and translocation from soil to wheat in an effluent irrigated field. Chemosphere 2017, 174, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Anwar, H.; Shahid, M.; Niazi, N.K.; Khalid, S.; Tariq, T.Z.; Ahmad, S.; Nadeem, M.; Abbas, G. Risk as-sessment of potentially toxic metal (loid) in Vigna radiata L. under wastewater and freshwater irrigation. Chemosphere 2021, 265, 129124. [Google Scholar] [CrossRef]
- Abdal, N.; Abbas, G.; Asad, S.A.; Ghfar, A.A.; Shah, G.M.; Rizwan, M.; Ali, S.; Shahbaz, M. Salinity mitigates cadmi-um-induced phytotoxicity in quinoa (Chenopodium quinoa Willd) by limiting the Cd uptake and improved responses to ox-idative stress: Implications for phytoremediation. Environ. Geochem. Health 2021, 1–15. [Google Scholar] [CrossRef]
- Waseem, A.; Arshad, J.; Iqbal, F.; Sajjad, A.; Mehmood, Z.; Murtaza, G. Pollution Status of Pakistan: A Retrospective Review on Heavy Metal Contamination of Water, Soil, and Vegetables. BioMed Res. Int. 2014, 2014, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M.C. Cadmium Bioavailability, Uptake, Toxicity and Detoxification in Soil-Plant System. Rev. Environ. Contam. Toxicol. 2016, 241, 73–137. [Google Scholar] [CrossRef]
- Zhang, S.; Ni, X.; Arif, M.; Zheng, J.; Stubbs, A.; Li, C. NaCl improved Cd tolerance of the euhalophyte Suaeda glauca but not the recretohalophyte Limonium aureum. Plant Soil 2020, 449, 303–318. [Google Scholar] [CrossRef]
- Shanying, H.E.; Xiaoe, Y.A.N.G.; Zhenli, H.E.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A review. Pedosphere 2017, 27, 421–443. [Google Scholar]
- Amjad, M.; Iqbal, M.M.; Abbas, G.; Farooq, A.B.U.; Naeem, M.A.; Imran, M.; Murtaza, B.; Nadeem, M.; Jacobsen, S.-E. Assessment of cadmium and lead tolerance potential of quinoa (Chenopodium quinoa Willd) and its implications for phy-toremediation and human health. Environ. Geochem. Health 2021, 1–14. [Google Scholar] [CrossRef]
- Rehman, S.; Abbas, G.; Shahid, M.; Saqib, M.; Farooq, A.B.U.; Hussain, M.; Farooq, A. Effect of salinity on cadmium tolerance, ionic homeostasis and oxidative stress responses in conocarpus exposed to cadmium stresss. Ecotoxicol. Environ. 2019, 171, 146–153. [Google Scholar] [CrossRef]
- Shabbir, A.; Saqib, M.; Murtaza, G.; Abbas, G.; Imran, M.; Rizwan, M.; Naeem, M.A.; Ali, S.; Javeed, H.M.R. Biochar mitigates arsenic-induced human health risks and phytotoxicity in quinoa under saline conditions by modulating ionic and oxidative stress responses. Environ. Pollut. 2021, 287, 117348. [Google Scholar] [CrossRef]
- Shahid, M.; Farooq, A.B.U.; Rabbani, F.; Khalid, S.; Dumat, C. Risk assessment and biophysiochemical responses of spinach to foliar application of lead oxide nanoparticles: A multivariate analysis. Chemosphere 2019, 245, 125605. [Google Scholar] [CrossRef]
- Kushwaha, A.; Hans, N.; Kumar, S.; Rani, R. A critical review on speciation, mobilization and toxicity of lead in soil-microbe-plant system and bioremediation strategies. Ecotoxicol. Environ. Saf. 2018, 147, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, A.; Abbas, G.; Saqib, M.; Shabbir, A.; Amjad, M.; Shahid, M.; Ahmad, I.; Iqbal, S.; Qaisrani, S.A. Salinity modulates lead (Pb) tolerance and phytoremediation potential of quinoa: A multivariate comparison of physiological and biochemical attributes. Environ. Geochem. Health 2021, 44, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Arias, J.A.; Peralta-Videa, J.R.; Ellzey, J.T.; Ren, M.; Viveros, M.N.; Gardea-Torresdey, J.L. Effects of Glomus deserticola inoculation on Prosopis: Enhancing chromium and lead uptake and translocation as confirmed by X-ray mapping, ICP-OES and TEM techniques. Environ. Exp. Bot. 2010, 68, 139–148. [Google Scholar] [CrossRef]
- Shahid, M.; Pinelli, E.; Pourrut, B.; Silvestre, J.; Dumat, C. Lead-induced genotoxicity to Vicia faba L. roots in relation with metal cell uptake and initial speciation. Ecotoxicol. Environ. Saf. 2011, 74, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, H.; Corso, M.; Verbruggen, N.; Clemens, S. CAX1 suppresses Cd-induced gen- eration of reactive oxygen species in Arabidopsis halleri. Plant. Cell. Environ. 2018, 41, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K. Lead exposure and its impact on immune system: A review. Toxicol. In Vitro 2009, 23, 969–972. [Google Scholar] [CrossRef]
- Murtaza, B.; Naeem, F.; Shahid, M.; Abbas, G.; Shah, N.S.; Amjad, M.; Bakhat, H.F.; Imran, M.; Niazi, N.K.; Murtaza, G. A multivariate analysis of physiological and antioxidant responses and health hazards of wheat under cadmium and lead stress. Environ. Sci. Pollut. Res. 2018, 26, 362–370. [Google Scholar] [CrossRef]
- Iqbal, S.; Basra, S.M.A.; Afzal, I.; Wahid, A.; Saddiq, M.S.; Hafeez, M.B.; Jacobsen, S.-E. Yield potential and salt tolerance of quinoa on salt-degraded soils of Pakistan. J. Agron. Crop Sci. 2019, 205, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Abbas, G.; Amjad, M.; Saqib, M.; Murtaza, B.; Asif Naeem, M.; Shabbir, A.; Murtaza, G. Soil sodicity is more detrimental than salinity for quinoa (Chenopodium quinoa Willd.): A multivariate comparison of physiological, biochemical and nutritional quality attributes. J. Agron. Crop Sci. 2021, 207, 59–73. [Google Scholar] [CrossRef]
- Shabala, S.; Hariadi, Y.; Jacobsen, S.-E. Genotypic difference in salinity tolerance in quinoa is determined by differential control of xylem Na+ loading and stomatal density. J. Plant Physiol. 2013, 170, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Islam, E.; Liu, D.; Li, T.; Yang, X.; Jin, X.; Mahmood, Q.; Tian, S.; Li, J. Effect of Pb toxicity on leaf growth, physiology and ultrastructure in the two ecotypes of Elsholtzia argyi. J. Hazard. Mater. 2008, 154, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxida- tive stress, anti-oxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Hemeda, H.M.; Klein, B.P. Effects of Naturally Occurring Antioxidants on Peroxidase Activity of Vegetable Extracts. J. Food Sci. 1990, 55, 184–185. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Steel, R.; Torrie, J.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Panda, A.; Rangani, J.; Kumari, A.; Parida, A. Efficient regulation of arsenic translocation to shoot tissue and modulation of phytochelatin levels and antioxidative defense system confers salinity and arsenic tolerance in the Halophyte Suaeda maritima. Environ. Exp. Bot. 2017, 143, 149–171. [Google Scholar] [CrossRef]
- Shabir, R.; Abbas, G.; Saqib, M.; Shahid, M.; Shah, G.M.; Akram, M.; Niazi, N.K.; Naeem, M.A.; Hussain, M.; Ashraf, F. Cadmium tolerance and phytoremediation potential of acacia (Acacia nilotica L.) under salinity stress. Int. J. Phytoremediation 2018, 20, 739–746. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amjad, M.; Akhtar, S.S.; Yang, A.; Akhtar, J.; Jacobsen, S.E. Antioxidative Response of Quinoa Exposed to Iso-Osmotic, Ionic and Non-Ionic Salt Stress. J. Agron. Crop Sci. 2015, 201, 452–460. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Wang, X.; Yang, X.; Cui, Z. Bioaugmentation-assisted phytoremediation of lead and salinity co-contaminated soil by Suaeda salsa and Trichoderma asperellum. Chemosphere 2019, 224, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar] [PubMed] [Green Version]
- Souri, Z.; Karimi, N.; De Oliveira, L.M. Antioxidant enzymes responses in shoots of arsenic hyperaccumulator, Isatis cappadocica Desv., under interaction of arsenate and phosphate. Environ. Technol. 2017, 39, 1316–1327. [Google Scholar] [CrossRef]
- Abbaspour, A.; Kalbasi, M.; Hajrasuliha, S.; Fotovat, A. Effect of Organic Matter and Salinity on Ethylenediaminetetraacetic Acid–Extractable and Solution Species of Cadmium and Lead in Three Agricultural Soils. Commun. Soil Sci. Plant Anal. 2008, 39, 983–1005. [Google Scholar] [CrossRef]
- Nawrot, N.; Wojciechowska, E.; Pazdro, K.; Szmagliński, J.; Pempkowiak, J. Uptake, accumulation, and translocation of Zn, Cu, Pb, Cd, Ni, and Cr by P. australis seedlings in an urban dredged sediment mesocosm: Impact of seedling origin and initial trace metal content. Sci. Total Environ. 2021, 768, 144983. [Google Scholar] [CrossRef]
- Xu, J.; Hu, C.; Wang, M.; Zhao, Z.; Zhao, X.; Cao, L.; Lu, Y.; Cai, X. Changeable effects of coexisting heavy metals on transfer of cadmium from soils to wheat grains. J. Hazard. Mater. 2021, 423, 127182. [Google Scholar] [CrossRef]
- Yan, X.; An, J.; Yin, Y.; Gao, C.; Wang, B.; Wei, S. Heavy metals uptake and translocation of typical wetland plants and their ecological effects on the coastal soil of a contaminated bay in Northeast China. Sci. Total Environ. 2021, 803, 149871. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments (µM) | Shoot Length (cm) | Root Length (cm) | Shoot Dry Weight (g plant−1) | Root Dry Weight (g plant−1) |
|---|---|---|---|---|
| Control | 18.0 ± 0.60 a | 23 ± 0.5 a | 0.34 ± 0.01 a | 0.32 ± 0.02 a |
| Cd 50 | 14.0 ± 0.23 c | 18 ± 0.8 c | 0.26 ± 0.008 b | 0.26 ± 0.008 b |
| Cd 100 | 12.0 ± 0.50 d | 15 ± 0.7 d | 0.21 ± 0.01 c | 0.22 ± 0.01 c |
| Pb 250 | 16.0 ± 0.40 b | 20 ± 0.5 b | 0.30 ± 0.013 ab | 0.30 ± 0.012 ab |
| Pb 500 | 13.0 ± 0.30 cd | 17 ± 0.4 c | 0.26 ± 0.005 b | 0.25 ± 0.008 b |
| Cd 50 + Pb 250 | 12.0 ± 0.50 d | 17 ± 0.8 c | 0.24 ± 0.01 bc | 0.24 ± 0.015 bc |
| Cd 100 + Pb 250 | 10.5 ± 0.20 e | 13 ± 0.5 de | 0.18 ± 0.008 cd | 0.18 ± 0.012 d |
| Cd 50 + Pb 500 | 9.0 ± 0.30 f | 12 ± 0.4 e | 0.16 ± 0.01 d | 0.19 ± 0.008 d |
| Cd 100 + Pb 500 | 7.0 ± 0.40 g | 9 ± 0.5 f | 0.13 ± 0.009 e | 0.14 ± 0.01 e |
| Treatments (µM) | Chl-a (µg g−1 FW) | Chl-b (µg g−1 FW) | Total Chl (µg g−1 FW) | Stomatal Conductance (mmol m−2s−1) |
|---|---|---|---|---|
| Control | 245 ± 8.0 a | 140 ± 5.0 a | 385 ± 10.0 a | 500 ± 10.0 a |
| Cd 50 | 205 ± 7.0 bc | 110 ± 3.0 bc | 315 ± 13.0 bc | 450 ± 8.0 b |
| Cd 100 | 170 ± 3.0 cd | 95 ± 4.0 cd | 265 ± 4.0 d | 400 ± 15.0 c |
| Pb 250 | 210 ± 8.0 b | 115 ± 2.0 bc | 325 ± 10.0 cd | 470 ± 8.0 ab |
| Pb 500 | 190 ± 5.0 bc | 100 ± 2.0 cd | 290 ± 5.0 c | 430 ± 16.0 b |
| Cd 50 + Pb 250 | 180 ± 4.0 d | 90 ± 3.0 d | 270 ± 10.0 cd | 410 ± 18.0 bc |
| Cd 100 + Pb 250 | 160 ± 8.0 e | 80 ± 4.0 e | 240 ± 5.0 e | 350 ± 10.0 d |
| Cd 50 + Pb 500 | 140 ± 4.0 f | 65 ± 3.0 f | 205 ± 10.0 f | 310 ± 8.0 e |
| Cd 100 + Pb 500 | 110 ± 5.0 g | 55 ± 2.0 g | 165 ± 5.0 g | 265 ± 12.0 f |
| Cd | Pb | ||||||
|---|---|---|---|---|---|---|---|
| Treatments (µM) | BCF Shoot | BCF Root | TF | BCF Shoot | BCF Root | TF | TI |
| Cd 50 | 8.83 ± 0.5 b | 9.8 ± 0.4 c | 0.9 ± 0.05 a | – | – | 0.18 ± 0.005 b | 76.5 ± 3 b |
| Cd 100 | 5.78 ± 0.2 c | 6.4 ± 0.8 d | 0.90 ± 0.04 a | – | – | 0.21 ± 0.008 a | 61.8 ± 2 c |
| Pb 250 | – | – | 0.5 ± 0.03 c | 4.30 ± 0.05 a | 24.4 ± 1.8 ab | 0.16 ± 0.01 b | 88.2 ± 3 a |
| Pb 500 | – | – | 0.8 ± 0.02 b | 2.90 ± 0.04 c | 17.6 ± 1.5 c | 0.16 ± 0.005 c | 76.5 ± 3.1 b |
| Cd 50 + Pb 250 | 9.79 ± 0.5 b | 12.5 ± 0.7 b | 0.79 ± 0.05 b | 3.67 ± 0.2 b | 22.8 ± 2.4 b | 0.16 ± 0.003 c | 70.6 ± 3 bc |
| Cd 100 + Pb 250 | 6.67 ± 0.4 c | 7.6 ± 0.5 d | 0.88 ± 0.02 a | 1.59 ± 0.1 d | 10.2 ± 1.2 d | 0.16 ± 0.005 c | 52.9 ± 1.5 d |
| Cd 50 + Pb 500 | 13.3 ± 0.3 a | 16.9 ± 0.6 a | 0.79 ± 0.03 b | 4.83 ± 0.2 a | 27.0 ± 2.5 a | 0.18 ± 0.006 b | 47.1 ± 2.5 e |
| Cd 100 + Pb 500 | 8.77 ± 0.2 b | 10.6 ± 1.2 bc | 0.82 ± 0.04 b | 1.93 ± 0.2 d | 11.6 ± 1.3 d | 0.17 ± 0.003 c | 38.2 ± 2 f |
| Variables | RDW | SDW | RL | SL | Chla | Chlb | TChl | H2O2 | MDA | MSI | SOD | CAT | POD | Cond | SK | RK | SCd | RCd | SPb | RPb |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SDW | 0.9879 | 1 | ||||||||||||||||||
| RL | 0.9895 | 0.9938 | 1 | |||||||||||||||||
| SL | 0.9808 | 0.9865 | 0.9885 | 1 | ||||||||||||||||
| Chla | 0.9704 | 0.9820 | 0.9897 | 0.9887 | 1 | |||||||||||||||
| Chlb | 0.9590 | 0.9755 | 0.9778 | 0.9909 | 0.9881 | 1 | ||||||||||||||
| TChl | 0.9687 | 0.9822 | 0.9878 | 0.9924 | 0.9982 | 0.9956 | 1 | |||||||||||||
| H2O2 | −0.9717 | −0.9783 | −0.9728 | −0.9713 | −0.9704 | −0.9665 | −0.9716 | 1 | ||||||||||||
| MDA | −0.9382 | −0.9381 | −0.9353 | −0.9450 | −0.9434 | −0.9489 | −0.9483 | 0.9837 | 1 | |||||||||||
| MSI | 0.9607 | 0.9684 | 0.9618 | 0.9809 | 0.9747 | 0.9793 | 0.9793 | −0.9868 | −0.9826 | 1 | ||||||||||
| SOD | −0.9311 | −0.9483 | −0.9509 | −0.9508 | −0.9558 | −0.9536 | −0.9576 | 0.9833 | 0.9753 | −0.9676 | 1 | |||||||||
| CAT | −0.9651 | −0.9659 | −0.9626 | −0.9682 | −0.9649 | −0.9657 | −0.9680 | 0.9968 | 0.9935 | −0.9910 | 0.9810 | 1 | ||||||||
| POD | −0.9168 | −0.9249 | −0.9274 | −0.9363 | −0.9410 | −0.9441 | −0.9449 | 0.9765 | 0.9931 | −0.9729 | 0.9886 | 0.9849 | 1 | |||||||
| Cond | 0.9708 | 0.9781 | 0.9809 | 0.9776 | 0.9824 | 0.9746 | 0.9821 | −0.9957 | −0.9810 | 0.9849 | −0.9893 | −0.9929 | −0.9807 | 1 | ||||||
| SK | 0.9655 | 0.9718 | 0.9742 | 0.9839 | 0.9820 | 0.9792 | 0.9837 | −0.9902 | −0.9727 | 0.9899 | −0.9824 | −0.9895 | −0.9725 | 0.9930 | 1 | |||||
| RK | 0.9656 | 0.9723 | 0.9724 | 0.9812 | 0.9830 | 0.9716 | 0.9813 | −0.9863 | −0.9749 | 0.9920 | −0.9760 | −0.9862 | −0.9728 | 0.9914 | 0.9914 | 1 | ||||
| SCd | −0.9070 | −0.9170 | −0.8818 | −0.8823 | −0.8640 | −0.8499 | −0.8609 | 0.8924 | 0.8665 | −0.8953 | 0.8324 | 0.8813 | 0.8314 | −0.8749 | −0.8567 | −0.8974 | 1 | |||
| RCd | −0.9099 | −0.9245 | −0.8901 | −0.8990 | −0.8817 | −0.8735 | −0.8810 | 0.9113 | 0.8946 | −0.9209 | 0.8612 | 0.9041 | 0.8651 | −0.8955 | −0.8812 | −0.9197 | 0.9959 | 1 | ||
| SPb | −0.3167 | −0.3263 | −0.3771 | −0.4155 | −0.4289 | −0.5004 | −0.4584 | 0.3698 | 0.4131 | −0.4019 | 0.4296 | 0.4003 | 0.4452 | −0.4025 | −0.4364 | −0.3578 | −0.0098 | 0.0476 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamagoos, A.A.; Alharby, H.F.; Abbas, G. Differential Uptake and Translocation of Cadmium and Lead by Quinoa: A Multivariate Comparison of Physiological and Oxidative Stress Responses. Toxics 2022, 10, 68. https://doi.org/10.3390/toxics10020068
Bamagoos AA, Alharby HF, Abbas G. Differential Uptake and Translocation of Cadmium and Lead by Quinoa: A Multivariate Comparison of Physiological and Oxidative Stress Responses. Toxics. 2022; 10(2):68. https://doi.org/10.3390/toxics10020068
Chicago/Turabian StyleBamagoos, Atif A., Hesham F. Alharby, and Ghulam Abbas. 2022. "Differential Uptake and Translocation of Cadmium and Lead by Quinoa: A Multivariate Comparison of Physiological and Oxidative Stress Responses" Toxics 10, no. 2: 68. https://doi.org/10.3390/toxics10020068
APA StyleBamagoos, A. A., Alharby, H. F., & Abbas, G. (2022). Differential Uptake and Translocation of Cadmium and Lead by Quinoa: A Multivariate Comparison of Physiological and Oxidative Stress Responses. Toxics, 10(2), 68. https://doi.org/10.3390/toxics10020068