Clusters of Lactobacillus Strains from Vegetal Origins Are Associated with Beneficial Functions: Experimental Data and Statistical Interpretations

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Antibacterial Activity
2.3. Quantification of the Lactic Acid
2.4. Biofilm Formation Assessment of Lactobacillus Strains on Polystyrene Tissue Culture Plates (TCP)
2.5. Exopolysaccharide Production
2.6. Inhibition of Biofilm Formation by Lactobacillus CFS
2.7. Ultra-Structure Alterations as Visualized by Transmission Electron Microscopy (TEM)
2.8. Co-Aggregation Test
2.9. Inhibition of Pathogenic Strains Adhesion to Caco-2 Cells by Lactobacillus Strains
2.10. CFS, Intact Cells and Intracellular Cell-Free Extract Scavenging Activity on Free Radical DPPH
2.11. Statistical Analysis
3. Results
3.1. Lactobacillus Strains Displayed Antibacterial Properties through Different Mechanisms
3.2. Lactic Acid Quantification
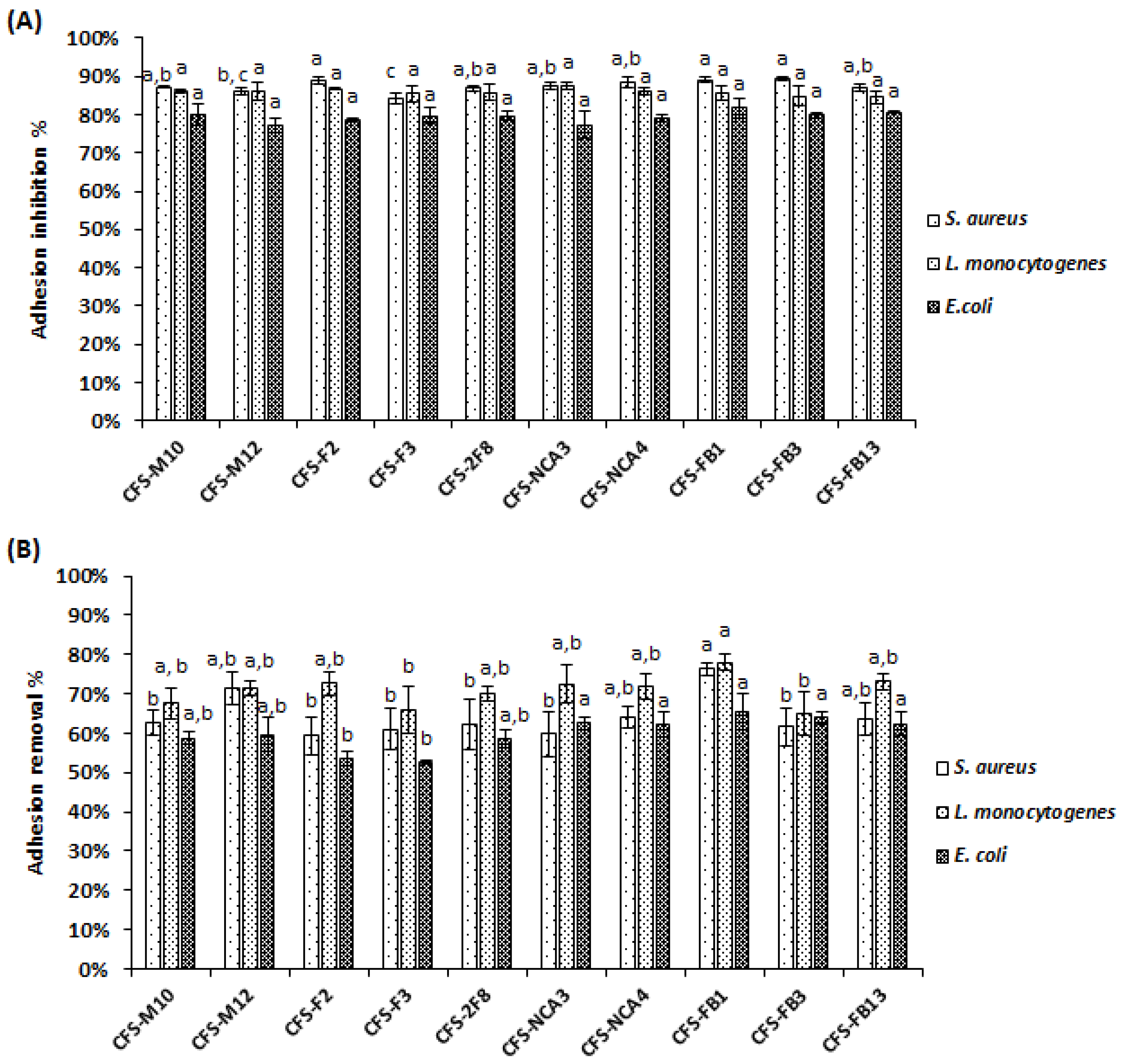
3.3. Adhesive Capacity of Lactobacillus and Pathogenic Strains on Polystyrene Microplates
3.4. The CFS from Lactobacillus Strains Were Able to Prevent the Pathogens Biofilm Formation
3.5. Exopolysaccharide Production
3.6. Cellular Ultra-Structural Alterations of Pathogens Treated with CFS Examined by Transmission Electron Microscopy (TEM)
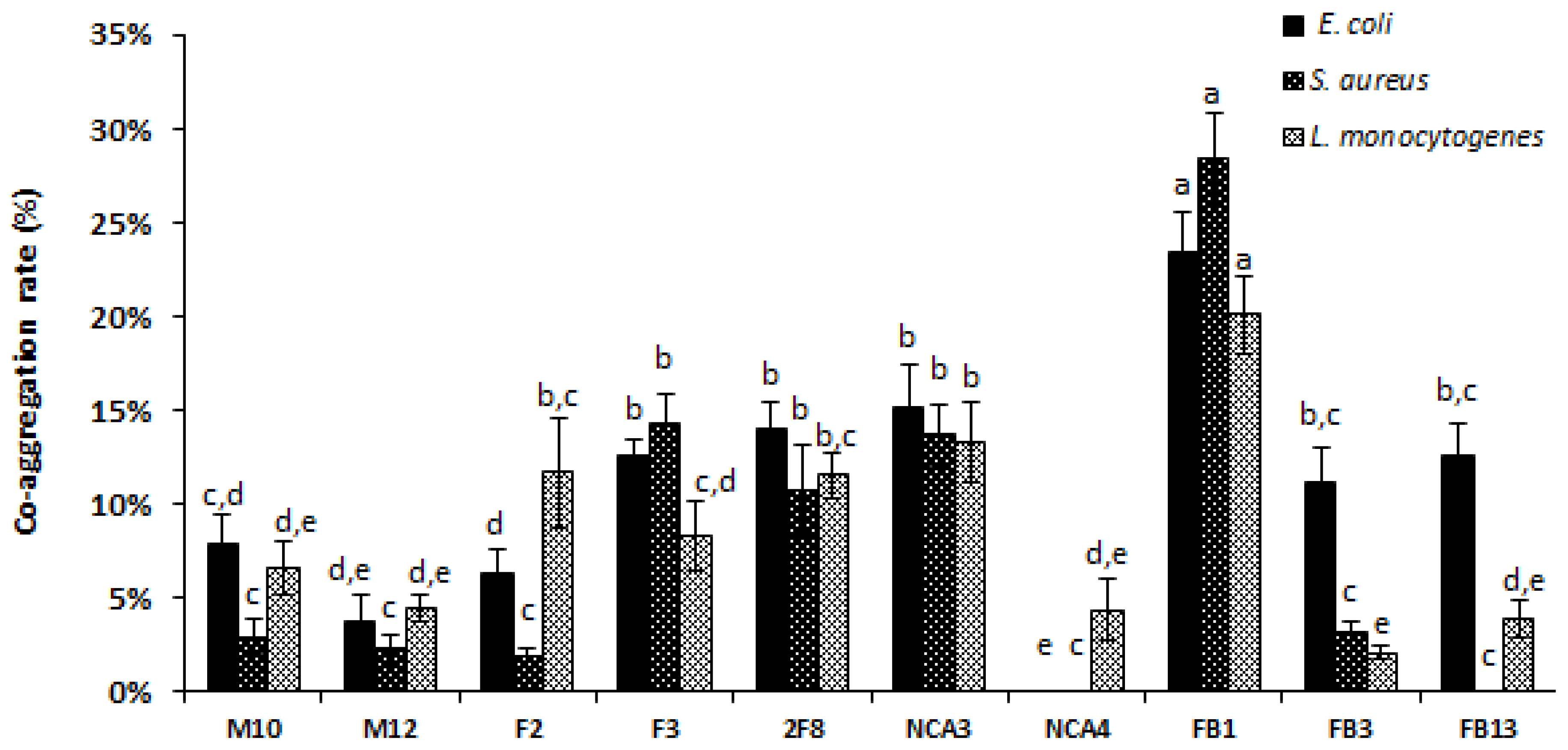
3.7. The Co-Aggregation Ability of Lactobacilli with Pathogens
3.8. Anti-Adhesive Activity of Lactobacillus Strains towards Pathogens on Caco-2 Cells
3.9. DPPH Free Radical Scavenging Activity
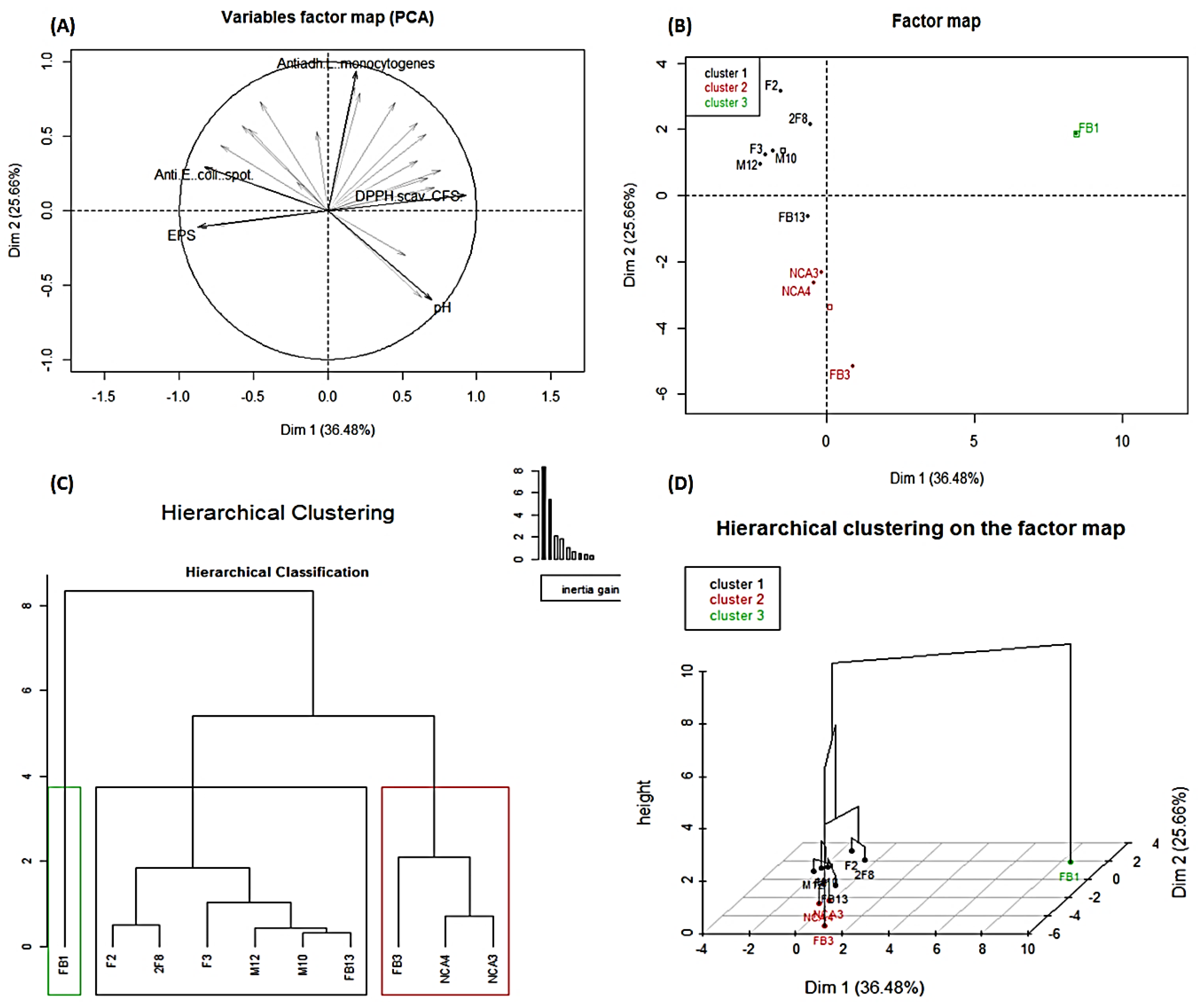
3.10. Principal Component Analysis and Summary Hierarchical Classification of the Ten Studied Lactobacillus Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jacob, C.; Mathiasen, L.; Powell, D. Designing effective messages for microbial food safety hazards. Food Control 2010, 21, 1–6. [Google Scholar] [CrossRef]
- Mousavi Khaneghah, A.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in hosts and foods: A review. Trends Food Sci. Technol. 2020, 95, 205–218. [Google Scholar] [CrossRef]
- Rawat, S. Food Spoilage: Microorganisms and their prevention. Asian J. Plant Sci. Res. 2015, 5. [Google Scholar]
- Nyachuba, D.G. Foodborne illness: Is it on the rise? Nutr. Rev. 2010, 68, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Painter, J.A.; Hoekstra, R.M.; Ayers, T.; Tauxe, R.V.; Braden, C.R.; Angulo, F.J.; Griffin, P.M. Attribution of Foodborne Illnesses, Hospitalizations, and Deaths to Food Commodities by using Outbreak Data, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Zhang, W. Understanding China’s food safety problem: An analysis of 2387 incidents of acute foodborne illness. Food Control 2013, 30, 311–317. [Google Scholar] [CrossRef]
- Fetsch, A.; Johler, S. Staphylococcus aureus as a Foodborne Pathogen. Curr. Clin. Microbiol. Rep. 2018, 5, 88–96. [Google Scholar] [CrossRef]
- Kavitha, S.; Harikrishnan, A.; Jeevaratnam, K. Characterization and evaluation of antibacterial efficacy of a novel antibiotic-type compound from a probiotic strain Lactobacillus plantarum KJB23 against food-borne pathogens. LWT 2020, 118, 108759. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef]
- Chammem, N.; Issaoui, M.; De Almeida, A.I.D.; Delgado, A.M. Food Crises and Food Safety Incidents in European Union, United States, and Maghreb Area: Current Risk Communication Strategies and New Approaches. J. AOAC Int. 2018, 101, 923–938. [Google Scholar] [CrossRef]
- Ouarabi, L.; Chait, Y.A.; Seddik, H.A.; Drider, D.; Bendali, F. Newly Isolated Lactobacilli strains from Algerian Human Vaginal Microbiota: Lactobacillus fermentum Strains Relevant Probiotic’s Candidates. Probiotics Antimicrob. Proteins 2019, 11, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Del Nobile, M.A.; Lucera, A.; Costa, C.; Conte, A. Food applications of natural antimicrobial compounds. Front. Microbiol. 2012, 3, 287. [Google Scholar] [CrossRef]
- Corsetti, A.; Perpetuini, G.; Tofalo, R. 13—Biopreservation effects in fermented foods. In Advances in Fermented Foods and Beverages; Holzapfel, W., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2015; pp. 311–332. ISBN 978-1-78242-015-6. [Google Scholar]
- Thakkar, P.N.; Patel, A.; Modi, H.A.; Prajapati, J.B. Hypocholesterolemic Effect of Potential Probiotic Lactobacillus fermentum Strains Isolated from Traditional Fermented Foods in Wistar Rats. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef]
- Lin, X.; Xia, Y.; Yang, Y.; Wang, G.; Zhou, W.; Ai, L. Probiotic characteristics of Lactobacillus plantarum AR113 and its molecular mechanism of antioxidant. LWT 2020, 126, 109278. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. Effects of single- and multi-strain probiotics on biofilm formation and in vitro adhesion to bladder cells by urinary tract pathogens. Anaerobe 2014, 27, 71–76. [Google Scholar] [CrossRef]
- Cui, T.; Bai, F.; Sun, M.; Lv, X.; Li, X.; Zhang, D.; Du, H. Lactobacillus crustorum ZHG 2-1 as novel quorum-quenching bacteria reducing virulence factors and biofilms formation of Pseudomonas aeruginosa. LWT 2020, 117, 108696. [Google Scholar] [CrossRef]
- Tarrah, A.; Da Silva Duarte, V.; De Castilhos, J.; Pakroo, S.; Lemos Junior, W.J.F.; Luchese, R.H.; Fioravante Guerra, A.; Rossi, R.C.; Righetto Ziegler, D.; Corich, V.; et al. Probiotic potential and biofilm inhibitory activity of Lactobacillus casei group strains isolated from infant feces. J. Funct. Foods 2019, 54, 489–497. [Google Scholar] [CrossRef]
- Jang, H.J.; Lee, N.-K.; Paik, H.-D. Probiotic characterization of Lactobacillus brevis KU15153 showing antimicrobial and antioxidant effect isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 1521–1528. [Google Scholar] [CrossRef]
- Coenye, T.; Kjellerup, B.; Stoodley, P.; Bjarnsholt, T. The future of biofilm research—Report on the ‘2019 Biofilm Bash’. Biofilm 2020, 2, 100012. [Google Scholar] [CrossRef]
- Tan, Y.; Leonhard, M.; Moser, D.; Ma, S.; Schneider-Stickler, B. Inhibitory effect of probiotic lactobacilli supernatants on single and mixed non-albicans Candida species biofilm. Arch. Oral Biol. 2018, 85, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Campana, R.; Van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Davoren, M.J.; Liu, J.; Castellanos, J.; Rodríguez-Malavé, N.I.; Schiestl, R.H. A novel probiotic, Lactobacillus johnsonii 456, resists acid and can persist in the human gut beyond the initial ingestion period. Gut Microbes 2019, 10, 458–480. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, S.; Kim, S.H. Released exopolysaccharide (r-EPS) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157:H7. Biochem. Biophys. Res. Commun. 2009, 379, 324–329. [Google Scholar] [CrossRef]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Vitali, B.; Minervini, G.; Rizzello, C.G.; Spisni, E.; Maccaferri, S.; Brigidi, P.; Gobbetti, M.; Di Cagno, R. Novel probiotic candidates for humans isolated from raw fruits and vegetables. Food Microbiol. 2012, 31, 116–125. [Google Scholar] [CrossRef]
- De Albuquerque, T.M.R.; Garcia, E.F.; De Oliveira Araújo, A.; Magnani, M.; Saarela, M.; De Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Proteins 2018, 10, 704–716. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Kubow, S.; Sadiq, F.A. Isolation and in-vitro probiotic characterization of fructophilic lactic acid bacteria from Chinese fruits and flowers. LWT 2019, 104, 70–75. [Google Scholar] [CrossRef]
- Barache, N.; Ladjouzi, R.; Belguesmia, Y.; Bendali, F.; Drider, D. Abundance of Lactobacillus plantarum Strains with Beneficial Attributes in Blackberries (Rubus sp.), Fresh Figs (Ficus carica), and Prickly Pears (Opuntia ficus-indica) Grown and Harvested in Algeria. Probiotics Antimicrob. Proteins 2020. [Google Scholar] [CrossRef]
- Al-Seraih, A.; Belguesmia, Y.; Baah, J.; Szunerits, S.; Boukherroub, R.; Drider, D. Enterocin B3A-B3B produced by LAB collected from infant faeces: Potential utilization in the food industry for Listeria monocytogenes biofilm management. Antonie Leeuwenhoek 2017, 110, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Bendali, F.; Durand, A.; Hébraud, M.; Sadoun, D. Lactobacillus paracasei subsp. paracasei: An Algerian isolate with antibacterial activity against enteric pathogens and probiotic fitness. J. Food Nutr. Res. Slovak Repub. 2011, 50, 139–149. [Google Scholar]
- Ait Seddik, H.; Bendali, F.; Cudennec, B.; Drider, D. Anti-pathogenic and probiotic attributes of Lactobacillus salivarius and Lactobacillus plantarum strains isolated from feces of Algerian infants and adults. Res. Microbiol. 2017, 168, 244–254. [Google Scholar] [CrossRef]
- Ait Ouali, F.; Al Kassaa, I.; Cudennec, B.; Abdallah, M.; Bendali, F.; Sadoun, D.; Chihib, N.-E.; Drider, D. Identification of lactobacilli with inhibitory effect on biofilm formation by pathogenic bacteria on stainless steel surfaces. Int. J. Food Microbiol. 2014, 191, 116–124. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Van Geel-Schutten, G.H.; Flesch, F.; Ten Brink, B.; Smith, M.R.; Dijkhuizen, L. Screening and characterization of Lactobacillus strains producing large amounts of exopolysaccharides. Appl. Microbiol. Biotechnol. 1998, 50, 697–703. [Google Scholar] [CrossRef]
- Shokri, D.; Khorasgani, M.R.; Mohkam, M.; Fatemi, S.M.; Ghasemi, Y.; Taheri-Kafrani, A. The Inhibition Effect of Lactobacilli Against Growth and Biofilm Formation of Pseudomonas aeruginosa. Probiotics Antimicrob. Proteins 2018, 10, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kos, B.; Susković, J.; Vuković, S.; Simpraga, M.; Frece, J.; Matosić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef]
- Pinto, M.; Robineleon, S.; Appay, M.D.; Kedinger, M.; Triadou, N.; Dussaulx, E.; Lacroix, B.; Simonassmann, P.; Haffen, K.; Fogh, J.; et al. Enterocyte-like differentiation and polarization of the human-colon carcinoma cell-line Caco-2 in culture. Biol. Cell 1983, 47, 323–330. [Google Scholar]
- Bendali, F.; Hébraud, M.; Sadoun, D. Anti-bacterial and anti-adherence activities of a probiotic strain of Lactobacillus paracasei subsp. paracasei against Listeria monocytogenes. Int. J. Appl. Microbiol. Biotechnol. Res. 2014, 2, 52–63. [Google Scholar]
- Jayashree, S.; Karthikeyan, R.; Nithyalakshmi, S.; Ranjani, J.; Gunasekaran, P.; Rajendhran, J. Anti-adhesion Property of the Potential Probiotic Strain Lactobacillus fermentum 8711 Against Methicillin-Resistant Staphylococcus aureus (MRSA). Front. Microbiol. 2018, 9, 411. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Mahajan, R.; Attri, S.; Goel, G. Selection of indigenous Lactobacillus paracasei CD4 and Lactobacillus gastricus BTM 7 as probiotic: Assessment of traits combined with principal component analysis. J. Appl. Microbiol. 2017, 122, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Ianniello, R.G.; Ricciardi, A.; Parente, E.; Tramutola, A.; Reale, A.; Zotta, T. Aeration and supplementation with heme and menaquinone affect survival to stresses and antioxidant capability of Lactobacillus casei strains. LWT 2015, 60, 817–824. [Google Scholar] [CrossRef]
- Hnasko, R.M.; Lin, A.V.; Stanker, L.H.; Bala, K.; McGarvey, J.A. Prion Extraction Methods: Comparison of Bead Beating, Ultrasonic Disruption, and Repeated Freeze-Thaw Methodologies for the Recovery of Functional Renilla-Prion Fusion Protein from Bacteria. In Sample Preparation Techniques for Soil, Plant, and Animal Samples; Micic, M., Ed.; Springer Protocols Handbooks; Springer: New York, NY, USA, 2016; pp. 389–399. ISBN 978-1-4939-3185-9. [Google Scholar]
- Mallappa, R.H.; Singh, D.K.; Rokana, N.; Pradhan, D.; Batish, V.K.; Grover, S. Screening and selection of probiotic Lactobacillus strains of Indian gut origin based on assessment of desired probiotic attributes combined with principal component and heatmap analysis. LWT 2019, 105, 272–281. [Google Scholar] [CrossRef]
- Tang, H.; Qian, B.; Xia, B.; Zhuan, Y.; Yao, Y.; Gan, R.; Zhang, J. Screening of lactic acid bacteria isolated from fermented Cornus officinalis fruits for probiotic potential. J. Food Saf. 2018, 38, e12565. [Google Scholar] [CrossRef]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Effect of organic acids on biofilm formation and quorum signaling of pathogens from fresh fruits and vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef]
- Ait Seddik, H.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Fu, W.; Mathews, A.P. Lactic acid production from lactose by Lactobacillus plantarum: Kinetic model and effects of pH, substrate, and oxygen. Biochem. Eng. J. 1999, 3, 163–170. [Google Scholar] [CrossRef]
- Passos, F.V.; Fleming, H.P.; Ollis, D.F.; Felder, R.M.; McFeeters, R.F. Kinetics and Modeling of Lactic Acid Production by Lactobacillus plantarum. Appl. Environ. Microbiol. 1994, 60, 2627–2636. [Google Scholar] [CrossRef]
- Mahdhi, A.; Leban, N.; Chakroun, I.; Chaouch, M.A.; Hafsa, J.; Fdhila, K.; Mahdouani, K.; Majdoub, H. Extracellular polysaccharide derived from potential probiotic strain with antioxidant and antibacterial activities as a prebiotic agent to control pathogenic bacterial biofilm formation. Microb. Pathog. 2017, 109, 214–220. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Llamas, M.G.; Iraporda, C.; Dueñas, M.T.; Abraham, A.G.; Garrote, G.L. Impact of growth temperature on exopolysaccharide production and probiotic properties of Lactobacillus paracasei strains isolated from kefir grains. Food Microbiol. 2018, 69, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Sun, Y.; Wu, X.; Yuan, Y.; Dai, Y.; Jia, S. Emulsifying, Flocculating, and Physicochemical Properties of Exopolysaccharide Produced by Cyanobacterium Nostoc flagelliforme. Appl. Biochem. Biotechnol. 2014, 172, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Vasanthakumari, D.S.; Harikumar, S.; Beena, D.J.; Pandey, A.; Nampoothiri, K.M. Physicochemical Characterization of an Exopolysaccharide Produced by a Newly Isolated Weissella cibaria. Appl. Biochem. Biotechnol. 2015, 176, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.I.; Mizan, M.F.R.; Ashrafudoulla, M.; Nahar, S.; Joo, H.-J.; Jahid, I.K.; Park, S.H.; Kim, K.-S.; Ha, S.-D. Inhibitory effects of probiotic potential lactic acid bacteria isolated from kimchi against Listeria monocytogenes biofilm on lettuce, stainless-steel surfaces, and MBEC™ biofilm device. LWT 2020, 118, 108864. [Google Scholar] [CrossRef]
- Cotar, I.A.; Saviuc, C.; Andreea Nita, R.; Bezirtzoglou, E.; Lazar, V.; Carmen Chifiriuc, M. Anti-pathogenic Strategies for Fighting Pseudomonas aeruginosa Infections- probiotic Soluble Compounds as Inhibitors of Quorum Sensing Genes Expression. Curr. Org. Chem. 2013, 17, 155–161. [Google Scholar] [CrossRef]
- Bendali, F.; Gaillard-Martinie, B.; Hebraud, M.; Sadoun, D. Kinetic of production and mode of action of the Lactobacillus paracasei subsp. paracasei anti-listerial bacteriocin, an Algerian isolate. LWT 2008, 41, 1784–1792. [Google Scholar] [CrossRef]
- Rybalchenko, O.V.; Bondarenko, V.M.; Orlova, O.G.; Markov, A.G.; Amasheh, S. Inhibitory effects of Lactobacillus fermentum on microbial growth and biofilm formation. Arch. Microbiol. 2015, 197, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Sambanthamoorthy, K.; Feng, X.; Patel, R.; Patel, S.; Paranavitana, C. Antimicrobial and antibiofilm potential of biosurfactants isolated from lactobacilli against multi-drug-resistant pathogens. BMC Microbiol. 2014, 14, 197. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Wang, Z.; Yang, Q. Ability of Lactobacillus to inhibit enteric pathogenic bacteria adhesion on Caco-2 cells. World J. Microbiol. Biotechnol. 2011, 27, 881–886. [Google Scholar] [CrossRef]
- Lemichez, E.; Lecuit, M.; Nassif, X.; Bourdoulous, S. Breaking the wall: Targeting of the endothelium by pathogenic bacteria. Nat. Rev. Microbiol. 2010, 8, 93–104. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Merino, L.; Trejo, F.M.; De Antoni, G.; Golowczyc, M.A. Lactobacillus strains inhibit biofilm formation of Salmonella sp. isolates from poultry. Food Res. Int. 2019, 123, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Beganović, J.; Frece, J.; Kos, B.; Leboš Pavunc, A.; Habjanič, K.; Šušković, J. Functionality of the S-layer protein from the probiotic strain Lactobacillus helveticus M92. Antonie Leeuwenhoek 2011, 100, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, V.; Quiberoni, A.; Reinheimer, J.; Suárez, V. Functional properties of Lactobacillus plantarum strains: A study in vitro of heat stress influence. Food Microbiol. 2016, 54, 154–161. [Google Scholar] [CrossRef]
- Kaewnopparat, S.; Dangmanee, N.; Kaewnopparat, N.; Srichana, T.; Chulasiri, M.; Settharaksa, S. In vitro probiotic properties of Lactobacillus fermentum SK5 isolated from vagina of a healthy woman. Anaerobe 2013, 22, 6–13. [Google Scholar] [CrossRef]
- Mu, G.; Li, H.; Tuo, Y.; Gao, Y.; Zhang, Y. Antioxidative effect of Lactobacillus plantarum Y44 on 2,2′-azobis(2-methylpropionamidine) dihydrochloride (ABAP)-damaged Caco-2 cells. J. Dairy Sci. 2019, 102, 6863–6875. [Google Scholar] [CrossRef]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as Potential Antioxidants: A Systematic Review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef]
- Lin, M.-Y.; Chang, F.-J. Antioxidative Effect of Intestinal Bacteria Bifidobacterium longum ATCC 15708 and Lactobacillus acidophilus ATCC 4356. Dig. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Screening for potential new probiotic based on probiotic properties and α-glucosidase inhibitory activity. Food Control 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Amaretti, A.; Di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-J.; Lee, J.-E.; Lim, S.-M.; Kim, Y.-J.; Lee, N.-K.; Paik, H.-D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Diameter of Inhibition Zones Given in Millimeters (mm) | Supernatants pH | |||||
|---|---|---|---|---|---|---|---|
| Spot-on-lawn Method | Wells Diffusion Method | ||||||
| E. coli ATCC 8739 | S. aureus 2S6 | L. monocytogenes 162 | E. coli ATCC 8739 | S. aureus 2S6 | L. monocytogenes 162 | ||
| M10 | 43± 1.00 a | 31 ± 1.04 a,b | 21 ± 1.04 d | 14 ± 0.29 a | 13 ± 1.00 a | 13 ± 0.35 a | 3.81 ± 0.00 c,d |
| M12 | 41 ± 1.52 a | 31 ± 1.00 a,b | 23 ± 1.04 b,c,d | 13 ± 0.35 a,b | 13 ± 0.50 a | 13 ± 0.16 a | 3.79 ± 0.02 c,d |
| F2 | 41 ± 1.51 a,b | 30 ± 1.23 a,b | 25 ± 0.50 a,b | 14 ± 0.50 a | 13 ± 0.46 a | 12 ± 0.17 a,b | 3.83 ± 0.01 c |
| F3 | 34 ± 0.51 c,d | 28 ± 1.14 b | 24 ± 1.04 a,b,c | 14 ± 0.60 a | 13 ± 0.29 a | 12 ± 0.29 a,b | 3.77 ± 0.02 d |
| 2F8 | 42 ± 1.72 a | 31 ± 1.30 a,b | 27 ± 0.50 a | 14 ± 0.80 a | 13 ± 0.23 a | 12 ± 0.35a,b | 3.79 ± 0.02 c,d |
| NCA3 | 37 ± 1.50 b,c | 30 ± 1.00 a,b | 23 ± 1.52 b,c,d | 12 ± 0.60 b | 10 ± 0.57 c | 11 ± 0.29 b | 3.94 ± 0.01 b |
| NCA4 | 36 ± 1.00 c,d | 29 ± 1.23 a,b | 22 ± 1.15 b,c,d | 12 ± 1.00 b | 10 ± 0.57 c | 11 ± 0.29 b | 3.98 ± 0.02 b |
| FB1 | 25 ± 2.00 e | 23 ± 0.20 c | 23 ± 0.50 b,c,d | 12 ± 0.30 b | 11 ± 0.46 b,c | 11 ± 0.17 b | 4.07 ± 0.01 a |
| FB3 | 32 ± 1.60 d | 31 ± 0.91 a | 22 ± 0.50 c,d | 12 ± 1.00 b | 11 ± 0.29 b,c | 11 ± 0.50 b | 4.10 ± 0.00 a |
| FB13 | 37 ± 1.80 b,c | 31 ± 1.00 a,b | 22 ± 1.50 c,d | 13 ± 0.65 a,b | 12 ± 0.35 a,b | 12 ± 1.00 a,b | 3.78 ± 0.01 d |
| Strain | Lactic Acid Quantification (g/L) | EPS Production (mg/L) | ||
|---|---|---|---|---|
| 8 h | 18 h | 24 h | ||
| M10 | 4.18 ± 0.06 b | 12.4 ± 0.64 a,b | 15.28 ± 0.13 b | 424.48 ± 23.59 b,c |
| M12 | 3.76 ± 0.06 c | 13.20 ± 0.28 a | 16.15 ± 0.18 a,b | 436.47 ± 14.82 a,b,c |
| F2 | 4.27 ± 0.06 b | 11.05 ± 1.03 b,c | 15.88 ± 0.22 a,b | 454.12 ± 25.53 a,b |
| F3 | 4.86 ± 0.04 a | 12.55 ± 0.43 a | 16.74 ± 0.36 a | 453.32 ± 17.55 a,b |
| 2F8 | 4.58 ± 0.07 a | 12.84 ± 0.14 a | 16.74 ± 0.30 a | 425.13 ± 11.81 b,c |
| NCA3 | 4.17 ± 0.24 b | 12.85 ± 0.26 a | 15.19 ± 0.24 a,b | 433.18 ± 14.57 b,c |
| NCA4 | 4.77 ± 0.02 a | 13.65 ± 0.13 a | 16.21 ± 0.62 a,b | 483.22 ± 15.39 a |
| FB1 | 3.01 ± 0.09 e | 9.88 ± 0.36 c | 10.73 ± 0.37 c | 315.55 ± 13.87 d |
| FB3 | 2.59 ± 0.13 f | 7.18 ± 0.36 d | 9.64 ± 0.33 d | 402.22 ± 18.76 c |
| FB13 | 3.40 ± 0.07 d | 12.57 ± 0.25 a | 15.67 ± 0.35 b | 412.69 ± 10.69 b,c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barache, N.; Belguesmia, Y.; Ladjouzi, R.; Bendali, F.; Drider, D. Clusters of Lactobacillus Strains from Vegetal Origins Are Associated with Beneficial Functions: Experimental Data and Statistical Interpretations. Foods 2020, 9, 985. https://doi.org/10.3390/foods9080985
Barache N, Belguesmia Y, Ladjouzi R, Bendali F, Drider D. Clusters of Lactobacillus Strains from Vegetal Origins Are Associated with Beneficial Functions: Experimental Data and Statistical Interpretations. Foods. 2020; 9(8):985. https://doi.org/10.3390/foods9080985
Chicago/Turabian StyleBarache, Nacim, Yanath Belguesmia, Rabia Ladjouzi, Farida Bendali, and Djamel Drider. 2020. "Clusters of Lactobacillus Strains from Vegetal Origins Are Associated with Beneficial Functions: Experimental Data and Statistical Interpretations" Foods 9, no. 8: 985. https://doi.org/10.3390/foods9080985
APA StyleBarache, N., Belguesmia, Y., Ladjouzi, R., Bendali, F., & Drider, D. (2020). Clusters of Lactobacillus Strains from Vegetal Origins Are Associated with Beneficial Functions: Experimental Data and Statistical Interpretations. Foods, 9(8), 985. https://doi.org/10.3390/foods9080985