Antioxidant and Anti-Inflammatory Activities of Safflower (Carthamus tinctorius L.) Honey Extract



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Safflower Honey Samples and Physical Deteriminations
2.3. Extraction on the Safflower Honey
2.4. Preliminary Analysis of ECH Phenolic Flavonoids by HPLC-QTOF-MS
2.5. In Vitro Free Radical Scavenging Ability Determination Experiment
2.5.1. DPPH Free Radical Scavenging Experiment
2.5.2. ABTS+ Free Radical Scavenging Experiment
2.6. Cell Experiment
2.6.1. ECH Measurement on Cell Relative Survival Rate
2.6.2. Determination of the Nitric Oxide (NO) Concentration in LPS-Induced Cells Treatment with ECH
2.6.3. ECH Detection of the mRNA Expression Related to Inflammation and Oxidation in LPS-Induced Cells
2.6.4. ECH Detection of Related Protein Expression in LPS-Induced Cells
2.6.5. Effect of ECH on LPS-Induced Nuclear Localization of p65 (NF-κB)
2.7. Statistics Analysis
3. Results
3.1. Physical and Chemical Analysis on the Safflower Honey
3.2. Preliminary Analysis of Phenolic Flavonoids in ECH by HPLC-QTOF-MS
3.3. In Vitro Antioxidant Free Radical Scavenging Capacity of ECH
3.4. In Vitro Antioxidant, Anti-Inflammatory Activies by ECH
3.4.1. Effects of ECH on Raw 264.7 Cell Survival
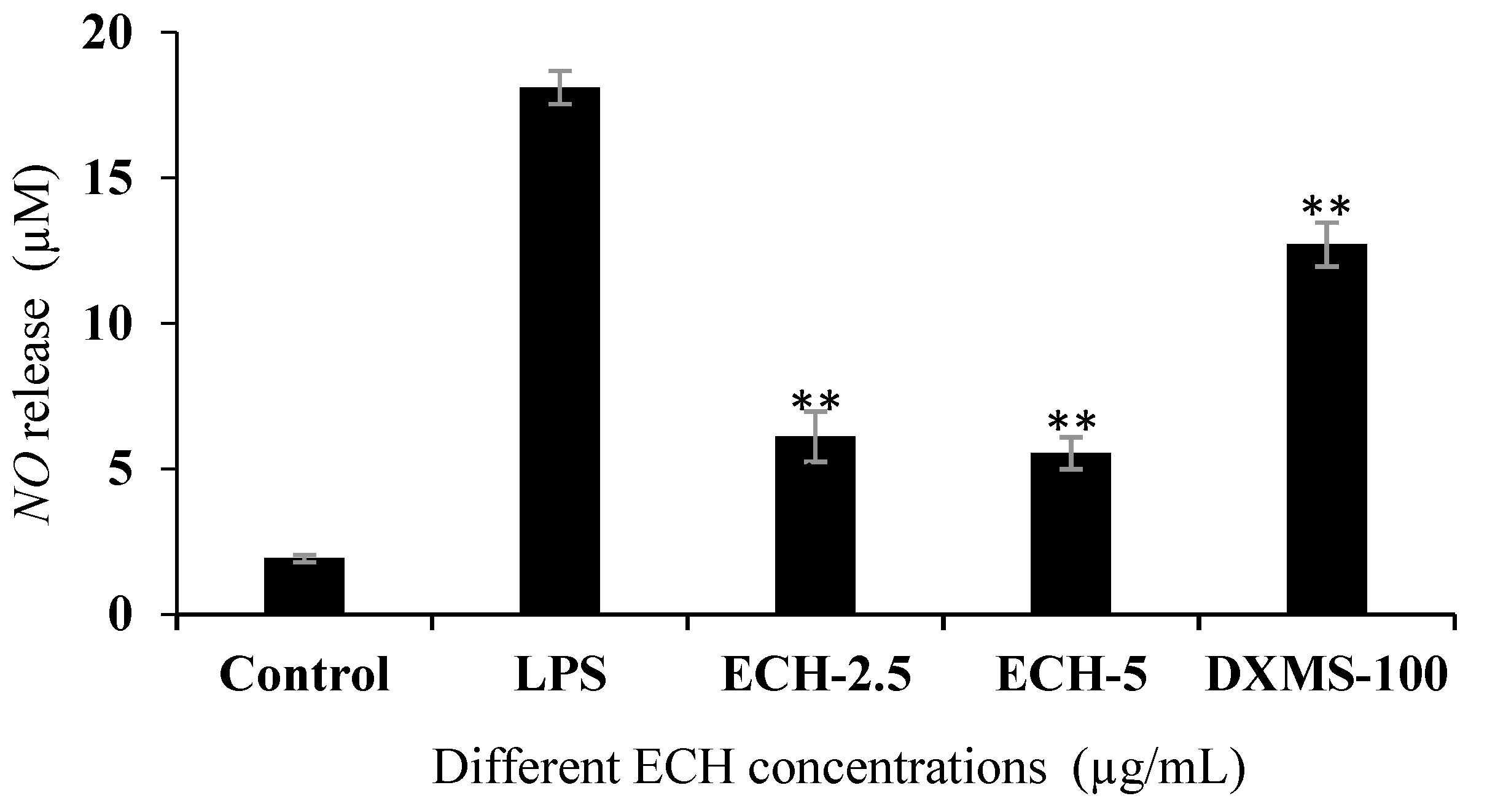
3.4.2. Effect of ECH on LPS-Induced Nitric Oxidase (NO) Release in RAW 264.7 Cells
3.4.3. Effect of ECH on LPS-Induced Inflammation and Oxidation-Related Gene Expression in RAW 264.7 Cells
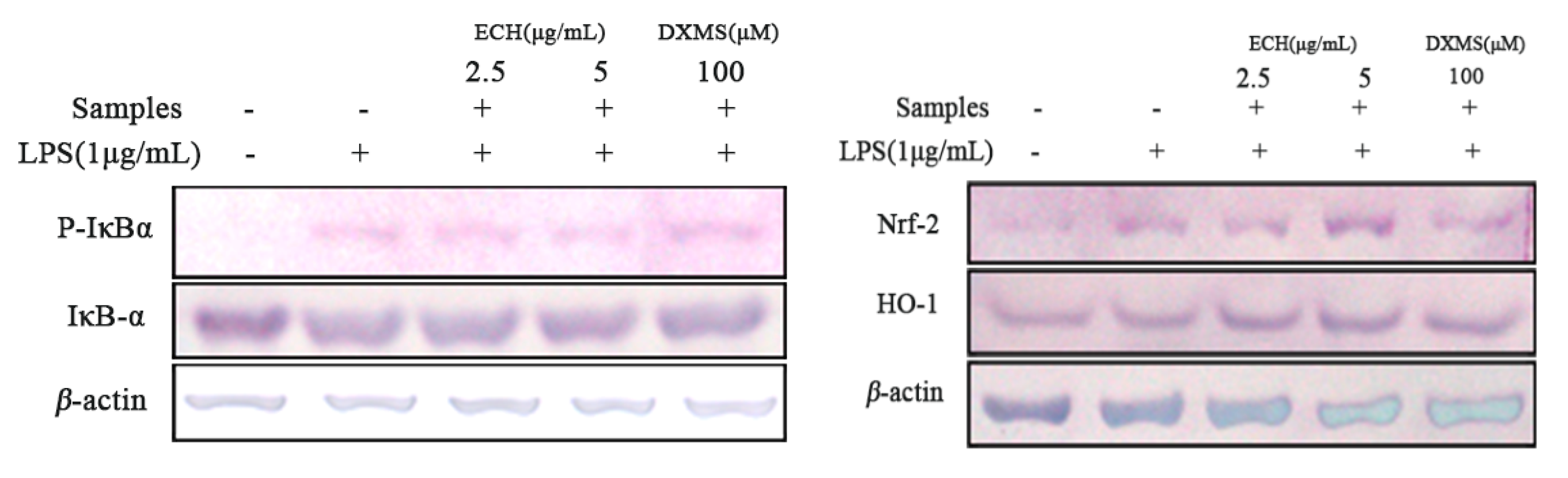
3.4.4. Effect of ECH on the Expressions of Inflammation and Anti-Oxidant Signaling Related Proteins in LPS-Activated RAW 264.7 Cells
3.4.5. Effect of ECH on the Nuclear Localization of NF-κB-p65 Induced by LPS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, T.; Wang, Q.; Cheng, N.; Cao, W. Effects of thermal processing on honey quality. Food Ferment. Ind. 2019, 45, 245–249. [Google Scholar] [CrossRef]
- Mandal, M.D.; Mandal, S. Honey: Its medicinal property and antibacterial activity. Asian Pac. J. Trop. Biomed. 2011, 1, 154–160. [Google Scholar] [CrossRef]
- Israili, Z.H. Antimicrobial Properties of Honey. Am. J. Ther. 2014, 21, 304–323. [Google Scholar] [CrossRef] [PubMed]
- Almasaudi, S.B.; El-Shitany, N.A.; Abbas, A.T.; Abdel-dayem, U.A.; Ali, S.S.; Al Jaouni, S.S.; Steve, H. Antioxidant, Anti-inflammatory, and Antiulcer Potential of Manuka Honey against Gastric Ulcer in Rats. Oxidative Med. Cell. Longev. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kassim, M.; Achoui, M.; Mustafa, M.R.; Mohd, M.A.; Yusoff, K.M. Ellagic acid, phenolic acids, and flavonoids in Malaysian honey extracts demonstrate in vitro anti-inflammatory activity. Nutr. Res. 2010, 30, 650–659. [Google Scholar] [CrossRef]
- Zhao, H.; Cheng, N.; He, L.; Peng, G.; Cao, W. Hepatoprotective Effects of the Honey of Apis cerana Fabricius on Bromobenzene-Induced Liver Damage in Mice. J. Food Sci. 2018, 83, 509–516. [Google Scholar] [CrossRef]
- Nooh, H.Z.; Nour-Eldien, N.M. The dual anti-inflammatory and antioxidant activities of natural honey promote cell proliferation and neural regeneration in a rat model of colitis. Acta Histochem. 2016, 118, 588–595. [Google Scholar] [CrossRef]
- Almasaudi, S.B.; Abbas, A.T.; Al-Handi, R.R.; El-Shitany, N.A.; Abdel-dayem, U.A.; Ali, S.S.; Saleh, R.M.; Al Jaouni, S.K.; Kamal, M.A.; Harakeh, S.M. Manuka Honey Exerts Antioxidant and Anti-Inflammatory Activities That Promote Healing of Acetic Acid-Induced Gastric Ulcer in Rats. Evid. Based Complement. Altern. Med. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Shamshuddin, N.S.S.; Zohdi, R.M. Gelam honey attenuates ovalbumin-induced airway inflammation in a mice model of allergic asthma. J. Tradit. Complement. Med. 2016, 8, 39–45. [Google Scholar] [CrossRef]
- Gül, A.; Pehlivan, T. Antioxidant Activities of some Monofloral Honey Types Produced Across Turkey. Saudi J. Biol. Sci. 2018, 25, 1056–1065. [Google Scholar] [CrossRef]
- Guo-Tai, W.U.; Yu-Peng, W.U.; Xiao-Fei, H.E.; Wang, X.F.; Niu, T.H.; Ren, Y. Progresses on Chemistry, Pharmacology and Application of Medihoney. J. Bee 2017, 1, 3–6. [Google Scholar]
- Dżugan, M.; Sowa, P.; Kwaśniewska, M.; Wesołowska, M.; Czernicka, M. Physicochemical Parameters and Antioxidant Activity of Bee Honey Enriched with Herbs. Plant Foods Hum. Nutr. 2017, 72, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.U.; Naz, S.; Abudabos, A.M. Towards a better understanding of the therapeutic applications and corresponding mechanisms of action of honey. Environ. Sci. Pollut. Res. 2017, 24, 27755–27766. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Zuo-Lin, Y.I.; Jin, X.L.; Chao, J.I.; Zhang, Z.Y. Chemical analysis on the safflower(Carthamus tinctorius)honey collected from Xinjiang. Sci. Technol. Food Ind. 2017, 38, 281–285. [Google Scholar]
- Yuan, W.; Ge, J.; Meng, W.; Ni, C.; Wei, C.; Jing, Z. A review of antioxidant activity in honey. Food Ferment. Ind. 2014, 40, 111–114. [Google Scholar]
- Erejuwa, O.O.; Sulaiman, S.A.; Ab Wahab, M.S. Honey: A novel antioxidant. Molecules 2012, 17, 4400–4423. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. Honey and Health: A Review of Recent Clinical Research. Pharmacogn. Res. 2017, 9, 121–127. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Yung An, C.; Rao, P.V.; Hawlader, M.N.I.; Azlan, S.A.B.M.; Sulaiman, S.A.; Gan, S.H. Identification of phenolic acids and flavonoids in monofloral honey from Bangladesh by high performance liquid chromatography: Determination of antioxidant capacity. Biomed. Res. Int. 2014, 2014, 737490. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, J.C.; Matus-Basto, A.J.; Acereto-Escoffié, P.; Segura-Campos, M.R. Antioxidant and anti-inflammatory activities of phenolic compounds isolated from Melipona beecheii honey. Food Agric. Immunol. 2017, 28, 1424–1437. [Google Scholar] [CrossRef]
- Anand, S.; Pang, E.; Livanos, G.; Mantri, N. Characterization of Physico-Chemical Properties and Antioxidant Capacities of Bioactive Honey Produced from Australian Grown Agastache rugosa and its Correlation with Colour and Poly-Phenol Content. Molecules 2018, 37, 108. [Google Scholar] [CrossRef]
- Ying, X.; Yu, K.; Chen, X.; Chen, H.; Hong, J.; Cheng, S.; Peng, L. Piperine inhibits LPS induced expression of inflammatory mediators in RAW 264.7 cells. Cellular Immunol. 2013, 285, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-S.; Shin, P.-G.; Lee, J.-H.; Kim, G.-D. The regulatory effect of veratric acid on NO production in LPS-stimulated RAW264.7 macrophage cells. Cellular Immunol. 2012, 280, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.-T.; Zhang, Z.-Y.; Jiang, C.-H.; Chen, J.-Q.; Ye, J.-Q.; Jia, X.-B.; Yang, Y.; Ni, Q.; Wang, S.-X.; Song, J.; et al. Nauclea officinalis inhibits inflammation in LPS-mediated RAW 264.7 macrophages by suppressing the NF-κB signaling pathway. J. Ethnopharmacol. 2016, 183, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Assis, P.O.A.D.; Guerra, G.C.B.; Araújo, D.F.D.S.; Araújo, R.F.D.; Queiroga, R.d.C.R.d.E. Intestinal anti-inflammatory activity of goat milk and goat yoghurt in the acetic acid model of rat colitis. Int. Dairy J. 2016, 56, 45–54. [Google Scholar] [CrossRef]
- Wang, K.; Ping, S.; Huang, S.; Hu, L. Molecular Mechanisms Underlying the In Vitro Anti-Inflammatory Effects of a Flavonoid-Rich Ethanol Extract from Chinese Propolis (Poplar Type). Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Pamukcu, B.; Lip, G.Y.H.; Shantsila, E. The nuclear factor–kappa B pathway in atherosclerosis: A potential therapeutic target for atherothrombotic vascular disease. Thromb. Res. 2011, 128, 117–123. [Google Scholar] [CrossRef]
- Roy, A.; Park, H.-J.; Jung, H.A.; Choi, J.S. Estragole Exhibits Anti-inflammatory Activity with the Regulation of NF-κB and Nrf-2 Signaling Pathways in LPS-induced RAW 264.7 cells. Nat. Prod. Sci. 2018, 24, 13–20. [Google Scholar] [CrossRef]
- Liu, J.; Tang, J.; Zuo, Y.; Yu, Y.; Luo, P.; Yao, X.; Dong, Y.; Wang, P.; Liu, L.; Zhou, H. Stauntoside B inhibits macrophage activation by inhibiting NF-κB and ERK MAPK signalling. Pharmacol. Res. 2016, 111, 303–315. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef]
- Song, D.; Lu, Z.; Wang, F.; Wang, Y. 041 Biogenic nano-selenium particles effectively attenuate oxidative stress–induced intestinal epithelial barrier injury by activating the Nrf2 antioxidant pathway. J. Anim. Sci. 2017, 95, 20–21. [Google Scholar] [CrossRef][Green Version]
- Hou, X.; Xua, X.; Li, H.; He, S.; Wana, C.; Yina, P.; Liu, M.; Liu, F.; Xu, J. Punicalagin Induces Nrf2/HO-1 expression via upregulation of PI3K/AKT pathway and inhibits LPS-induced oxidative stress in RAW264.7 Macrophages. Mediat. Inflamm. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kadri, S.M.; Zaluski, R.; Pereira Lima, G.P.; Mazzafera, P.; de Oliveira Orsi, R. Characterization of Coffea arabica monofloral honey from Espírito Santo, Brazil. Food Chem. 2016, 203, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Xue-Feng, M.U.; Sun, L.P.; Xiang, X.U.; Pang, J.; Wei, H.E.; Huang, L.; Shen, X.F. Optimization of Purification Process for DPPH Free Radical Scavenging Components from Acidic Aqueous Extract from Chinese Date Honey by Macroporous Resin Adsorption. Food Sci. 2011, 32, 98–102. [Google Scholar]
- Guo, N.; Zhao, L.; Zhao, Y.; Li, Q.; Xue, X.; Wu, L.; Gomez Escalada, M.; Wang, K.; Peng, W. Comparison of the Chemical Composition and Biological Activity of Mature and Immature Honey: An HPLC/QTOF/MS-Based Metabolomic Approach. J. Agric. Food Chem. 2020, 68, 4062–4071. [Google Scholar] [CrossRef]
- Wu, H.C.; Chen, H.-M.; Shiau, C.-Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Yang, H.; Dong, Y.; Du, H.; Shi, H.; Li, X. Antioxidant Compounds from Propolis Collected in Anhui, China. Molecules 2011, 16, 3444–3455. [Google Scholar] [CrossRef]
- Gutierrez, R.M.P.; Hoyo-Vadillo, C. Anti-inflammatory Potential of Petiveria alliacea on Activated RAW264.7 Murine Macrophages. Pharmacogn. Mag. 2017, 13, 174. [Google Scholar] [CrossRef]
- Li, Y.; Meng, T.; Hao, N.; Tao, H.; Zou, S.; Li, M.; Ming, P.; Ding, H.; Dong, J.; Feng, S. Immune regulation mechanism of Astragaloside IV on RAW264.7 cells through activating the NF-κB/MAPK signaling pathway. Int. Immunopharmacol. 2017, 49, 38–49. [Google Scholar] [CrossRef]
- Cui, C.; Lu, H.; Hui, Q.; Lu, S.; Liu, Y.; Ahmad, W.; Wang, Y.; Hu, P.; Liu, X.; Cai, Y.; et al. A preliminary investigation of the toxic effects of Benzylpenicilloic acid. Food Chem. Toxicol. 2018, 111, 567–577. [Google Scholar] [CrossRef]
- Ah, L.H.; Ram, S.B.; Ryeong, K.H.; Eun, K.J.; Bin, Y.W.; Ju, P.J.; Lim, L.M.; Young, C.J.; Seob, L.H.; Youn, H.D. Butanol extracts of Asparagus cochinchinensis fermented with Weissella cibaria inhibit iNOS-mediated COX-2 induction pathway and inflammatory cytokines in LPS-stimulated RAW264.7 macrophage cells. Exp. Ther. Med. 2017, 14, 4986–4994. [Google Scholar]
- Wang, K.; Hu, L.; Jin, X.-L.; Ma, Q.-X.; Marcucci, M.C.; Netto, A.A.L.; Sawaya, A.C.H.F.; Huang, S.; Ren, W.-K.; Conlon, M.A.; et al. Polyphenol-rich propolis extracts from China and Brazil exert anti-inflammatory effects by modulating ubiquitination of TRAF6 during the activation of NF-κB. J. Funct. Foods 2015, 19, 464–478. [Google Scholar] [CrossRef]
- Wang, B.; Chang, H.; Su, S.; Sun, L.; Wang, K. Antioxidative and Anti-inflammatory Activities of Ethanol Extract of Geopropolis from Stingless Bees. Sci. Agric. Sin. 2019, 52, 939–948. [Google Scholar]
- Özkök, D.; Silici, S. Effects of honey HMF on enzyme activities and serum biochemical parameters of Wistar rats. Environ. Sci. Pollut. Res. 2016, 23, 20186–20193. [Google Scholar] [CrossRef] [PubMed]
- Shapla, U.M.; Solayman, M.; Alam, N.; Khalil, M.I.; Gan, S.H. 5-Hydroxymethylfurfural (HMF) levels in honey and other food products: Effects on bees and human health. Chem. Cent. J. 2018, 12, 35. [Google Scholar] [CrossRef]
- El-Haskoury, R.; Kriaa, W.; Lyoussi, B.; Makni, M. Ceratonia siliqua honeys from Morocco: Physicochemical properties, mineral contents, and antioxidant activities. J. Food Drug Anal. 2018, 26, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Michalkiewicz, A.; Biesaga, M. Solid-phase extraction procedure for determination of phenolic acids and some flavonols in honey. J. Chromatogr. A 2008, 1187, 18–24. [Google Scholar] [CrossRef]
- Yung An, C.; Hossain, M.M.; Alam, F.; Islam, M.A.; Khalil, M.I.; Alam, N.; Gan, S.H. Efficiency of Polyphenol Extraction from Artificial Honey Using C18 Cartridges and Amberlite® XAD-2 Resin: A Comparative Study. J. Chem. 2016, 2016, 8356739. [Google Scholar] [CrossRef]
- Afroz, R.; Tanvir, E.M.; Paul, S.; Bhoumik, N.C.; Gan, S.H.; Khalil, M.I. DNA Damage Inhibition Properties of Sundarban Honey and its Phenolic Composition. J. Food Biochem. 2016, 40, 436–445. [Google Scholar] [CrossRef]
- Flanjak, I.; Kenjerić, D.; Bubalo, D.; Primorac, L. Characterisation of selected Croatian honey types based on the combination of antioxidant capacity, quality parameters, and chemometrics. Eur. Food Res. Technol. 2016, 242, 467–475. [Google Scholar] [CrossRef]
- Kaygusuz, H.; Tezcan, F.; Bedia Erim, F.; Yildiz, O.; Sahin, H.; Can, Z.; Kolayli, S. Characterization of Anatolian honeys based on minerals, bioactive components and principal component analysis. LWT Food Sci. Technol. 2016, 68, 273–279. [Google Scholar] [CrossRef]
- Calixto-Campos, C.; Carvalho, T.T.; Hohmann, M.S.N.; Pinho-Ribeiro, F.A.; Fattori, V.; Manchope, M.F.; Zarpelon, A.C.; Baracat, M.M.; Georgetti, S.R.; Casagrande, R.; et al. Vanillic Acid Inhibits Inflammatory Pain by Inhibiting Neutrophil Recruitment, Oxidative Stress, Cytokine Production, and NFκB Activation in Mice. J. Nat. Prod. 2015, 78, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Spilioti, E.; Jaakkola, M.; Tolonen, T.; Lipponen, M.; Virtanen, V.; Chinou, I.; Kassi, E.; Karabournioti, S.; Moutsatsou, P. Phenolic acid composition, antiatherogenic and anticancer potential of honeys derived from various regions in Greece. PLoS ONE 2014, 9, e94860. [Google Scholar] [CrossRef] [PubMed]
- Rajan, V.K.; Muraleedharan, K. A computational investigation on the structure, global parameters and antioxidant capacity of a polyphenol, Gallic acid. Food Chem. 2017, 220, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hou, L.; Xue, H.; Wang, C. Gallic acid inhibits inflammatory response of RAW264.7 macrophages by blocking the activation of TLR4/NF-κB induced by LPS. Chin. J. Cell. Mol. Immunol 2016, 32, 1610–1614. [Google Scholar]
- Basu Mallik, S.; Mudgal, J.; Nampoothiri, M.; Hall, S.; Dukie, S.A.; Grant, G.; Rao, C.M.; Arora, D. Caffeic acid attenuates lipopolysaccharide-induced sickness behaviour and neuroinflammation in mice. Neurosci. Lett. 2016, 632, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Sadar, S.S.; Vyawahare, N.S.; Bodhankar, S.L. Ferulic acid ameliorates TNBS-induced ulcerative colitis through modulation of cytokines, oxidative stress, iNOs, COX-2, and apoptosis in laboratory rats. EXCLI J. 2016, 15, 482–499. [Google Scholar]
- Batiha, G.E.-S.; Besbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef]
- Hou, W.; Hu, S.; Su, Z.; Wang, Q.; Meng, G.; Guo, T.; Zhang, J.; Gao, P. Myricetin attenuates LPS-induced inflammation in RAW 264.7 macrophages and mouse models. Future Med. Chem. 2018, 10, 2253–2264. [Google Scholar] [CrossRef]
- Tian, R.; Yang, W.; Xue, Q.; Gao, L.; Huo, J.; Ren, D.; Chen, X. Rutin ameliorates diabetic neuropathy by lowering plasma glucose and decreasing oxidative stress via Nrf2 signaling pathway in rats. Eur. J. Pharmacol. 2016, 771, 84–92. [Google Scholar] [CrossRef]
- De Oliveira, M.R.; Andrade, C.M.B.; Fürstenau, C.R. Naringenin Exerts Anti-inflammatory Effects in Paraquat-Treated SH-SY5Y Cells Through a Mechanism Associated with the Nrf2/HO-1 Axis. Neurochem. Res. 2018, 43, 894–903. [Google Scholar] [CrossRef]
- Chao, C.-S.; Tsai, C.-S.; Chang, Y.-P.; Chen, J.-M.; Chin, H.-K.; Yang, S.-C. Hyperin inhibits nuclear factor kappa B and activates nuclear factor E2-related factor-2 signaling pathways in cisplatin-induced acute kidney injury in mice. Int. Immunopharmacol. 2016, 40, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Ye, G.; Huang, B. Kaempferol Alleviates the Interleukin-1β-Induced Inflammation in Rat Osteoarthritis Chondrocytes via Suppression of NF-κB. Med. Sci. Monit. 2017, 23, 3925–3931. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Phase A% (0.1% Formic Acid) | Phase B% (100% Acetonitrile) |
|---|---|---|
| 0 | 90 | 10 |
| 0–15 | 75 | 25 |
| 15–20 | 70 | 30 |
| 20–30 | 65 | 35 |
| 30–35 | 30 | 70 |
| 35–40 | 30 | 70 |
| 40–42 | 90 | 10 |
| 42–50 | 90 | 10 |
| Gene | Upstream Primer Sequence | Downstream Primer Sequence |
|---|---|---|
| iNOS | 5′-TTTCCAGAAGCAGAATGTGACC-3′ | 5′-AACACCACTTTCACCAAGACTC-3′ |
| IL-1β | 5′-CCAACAAGTGATATTCTCCATGAG-3′ | 5′-ACTCTGCAGACTCAAACTCCA-3′ |
| TNF-α | 5′-CCACGCTCTTCTGTCTACTG-3′ | 5′-ACTTGGTGGTTTGCTACGAC-3′ |
| MCP-1 | 5′-AAGAAGCTGTAGTTTTTGTCACCA-3′ | 5′-TGAAGACCTTAGGGCAGATGC-3′ |
| HO-1 | 5′-ACATTGAGCTGTTTGAGGAG-3′ | 5′-TACATGGCATAAATTCCCACTG-3′ |
| TXNRD | 5′-AGGATTTCTGGCTGGTATCG-3′ | 5′-CTCGCTGTTTGTGGATTGAG-3′ |
| NQO | 5′-TTCAACCCCATCATTTCC-3′ | 5′-TCAGGCGTCCTTCCTTATA-3′ |
| Physical and Chemical Indicators | Result | Standard Limited | Standard Method |
|---|---|---|---|
| Moisture (%) | 18.2 ± 0.25 | ≤20 | 2001/110/EC |
| Acidity (mL/kg) | 25.0 ± 0.43 | ≤50 | 2001/110/EC |
| Amylase value (mL/(g·h)) | 21.1 ± 0.36 | ≥8 | 2001/110/EC |
| Hydroxymethylfurfural (mg/kg) | ND | ≤40 | 2001/110/EC |
| Fructose (%) | 36.9 ± 0.26 | ≥60 | 2001/110/EC |
| Glucose (%) | 25.2 ± 0.14 | ||
| Sucrose (%) | ND | ≤5 | 2001/110/EC |
| Ash (%) | ND | ≤0.1 | 2001/110/EC |
| Compound Name | Standard Curve Equation | R2 | Concentration Ranges (ng/mL) | RT (min) | [M − H]− | Content (mg/kg) |
|---|---|---|---|---|---|---|
| gallic acid | y = 573.42x − 32178.21 | 0.9983 | 50–2000 | 1.765 | 168.7 | 0.636 |
| Protocatechuic acid | y = 925.79x + 37631.28 | 0.9938 | 50–2000 | 2.501 | 153 | 0.115 |
| protocatechualdehyde | y = 338.47x + 18654.55 | 0.9991 | 50–1000 | 3.589 | 136.9 | ND |
| p-hydroxybenzoic acid | y = 437.73x + 2112.43 | 0.9975 | 50–2000 | 3.963 | 137 | 1.524 |
| vanillic acid | y = 17.86x − 166.86 | 0.9973 | 50–2000 | 4.865 | 167 | 3.196 |
| caffeic acid | y = 1377.37x + 6941.68 | 0.9984 | 50–2000 | 5.046 | 179 | 0.041 |
| ferulic acid | y = 108.21x − 1692.97 | 0.9989 | 50–2000 | 10.924 | 193.1 | 0.248 |
| cinnamic acid | y = 12.75x − 100.06 | 0.9983 | 50–2000 | 18.060 | 146.9 | 0.559 |
| rutin | y = 1017.39x − 35010.60 | 0.9993 | 50–2000 | 11.689 | 609.1 | 0.388 |
| quercetin-3-O-glucoside | y = 2782.17x − 74249.70 | 0.9993 | 50–2000 | 11.906 | 462.8 | 0.300 |
| kaempferol-3-O-glucoside | y = 2350.39x + 111703.32 | 0.9952 | 50–2000 | 13.776 | 446.8 | ND |
| myricetin | y = 0.91x − 77.39 | 0.9953 | 50–2000 | 15.033 | 317 | 1.021 |
| morin | y = 611.76x + 3378.56 | 0.9962 | 50–2000 | 16.578 | 301 | ND |
| luteolin | y = 1103.83x + 98837.97 | 0.9961 | 50–2000 | 18.06 | 285 | ND |
| quercetin | y = 3032.78x − 1438777.72 | 0.998 | 100–2000 | 18.153 | 301 | 5.342 |
| kaempferol | y = 10.83x − 72.24 | 0.9987 | 50–2000 | 20.439 | 271.1 | 0.126 |
| naringenin | y = 14.58x + 410.22 | 0.9969 | 50–2000 | 20.439 | 271.1 | 0.072 |
| pinobanksin | y = 71.63x + 450.56 | 0.995 | 50–2000 | 20.709 | 271.1 | 0.384 |
| diosmetin | y = 4225.03x + 392274.94 | 0.9916 | 50–2000 | 21.102 | 298.9 | ND |
| chrysin | y = 498.99x + 27824.49 | 0.9989 | 50–2000 | 25.205 | 253.1 | ND |
| pinocembrin | y = 340.00x + 29581.47 | 0.9902 | 50–2000 | 25.739 | 255.1 | ND |
| galanin | y = 120.24x + 9565.86 | 0.9952 | 50–2000 | 25.907 | 269 | ND |
| caffeic acid phenethyl ester | y = 15593.82x + 3956352.11 | 0.9976 | 100–2000 | 26.155 | 283.1 | ND |
| IC50 (µg/mL) | |
|---|---|
| DPPH | 68.23 ± 0.40 |
| ABTS+ | 81.88 ± 0.54 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.-P.; Shi, F.-F.; Zhang, W.-W.; Zhang, Z.-H.; Wang, K. Antioxidant and Anti-Inflammatory Activities of Safflower (Carthamus tinctorius L.) Honey Extract. Foods 2020, 9, 1039. https://doi.org/10.3390/foods9081039
Sun L-P, Shi F-F, Zhang W-W, Zhang Z-H, Wang K. Antioxidant and Anti-Inflammatory Activities of Safflower (Carthamus tinctorius L.) Honey Extract. Foods. 2020; 9(8):1039. https://doi.org/10.3390/foods9081039
Chicago/Turabian StyleSun, Li-Ping, Feng-Feng Shi, Wen-Wen Zhang, Zhi-Hao Zhang, and Kai Wang. 2020. "Antioxidant and Anti-Inflammatory Activities of Safflower (Carthamus tinctorius L.) Honey Extract" Foods 9, no. 8: 1039. https://doi.org/10.3390/foods9081039
APA StyleSun, L.-P., Shi, F.-F., Zhang, W.-W., Zhang, Z.-H., & Wang, K. (2020). Antioxidant and Anti-Inflammatory Activities of Safflower (Carthamus tinctorius L.) Honey Extract. Foods, 9(8), 1039. https://doi.org/10.3390/foods9081039