Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides


Abstract
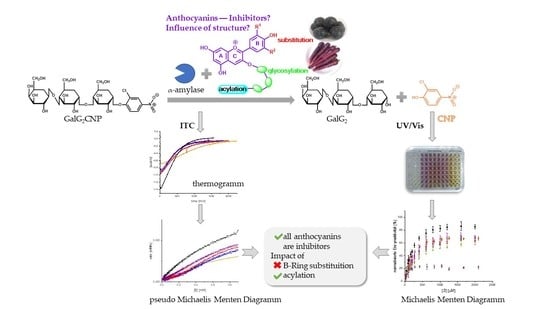
1. Introduction
2. Materials and Methods
2.1. Materials, Solvents, and Reagents
2.2. Preparation of the Black Carrot Anthocyanin Extract (BC-ACY) by Solid Phase Extraction
2.3. Isolation of Black Carrot Anthocyanins by Preparative HPLC
2.4. Structure Characterization of the Black Carrot Anthocyanins by Mass Spectrometry
2.5. Determination of the Absorption Coefficient of Black Carrot Anthocyanins Based on 1H-NMR
- IQRef = absolute integral of the considered signal from reference compounds
- SW = spectral width
- MW = molar weight (lactic acid: 90.080 g/mol; citric acid: 192.124 g/mol)
- SI = size of the real spectrum after Fourier transformation
- cQRef = concentration of lactic/citric acid in the QuantRef standard (mmol/L).
- NHQRef = number of protons of lactic/citric acid causing the signal under consideration
- DF = dilution factor
- ßx = mass concentration of the anthocyanin (mg/L).
- I = absolute integral of the anthocyanin signal under consideration
- SW = spectral width
- MWx = molar weight of the anthocyanin (g/mol).
- Px/PQRef = duration of the excitation pulse (µs).
- SI = size of the real spectrum after Fourier transformation
- NHx = number of anthocyanin protons causing the signal under consideration
- DF = dilution factor
- Abs = absorption at λmax
- MWx = molar weight of the anthocyanin (g/mol)
- ßx = average mass concentration of the anthocyanin determined by qNMR (mg/L)
- l = path length (1 cm)
- DF = dilution factor
- 1000 = conversion to L
2.6. Enzyme Activity Assay
- KM: Michaelis Menten constant (µM)
- vmax: Maximal conversion rate of the uninhibited reaction (µM/min)
- v0: Observed conversion rate at given substrate and inhibitor concentration (µM/min)
- [S]: Substrate concentration
- [I]: Inhibitor concentration
- Kic: Inhibitor constant, reflecting the competitive inhibition in the equation (µM)
- Kiu: Inhibitor constant, reflecting the uncompetitive inhibition in the equation (µM)
- vmaxapp: Calculated maximal conversion rate, when the Michaelis Menten equation is used on an inhibited curve (µM/min)
- Kmapp: Calculated Michaelis Menten constant, when the Michaelis Menten equation is used on an inhibited curve (µM)
- IC50: Half maximal inhibitory concentration (inhibitor concentration needed to achieve 50% inhibition; it is dependent on the substrate concentration) (µM)
2.7. Isothermal Titration Calorimetry (ITC)
3. Results
3.1. Determination of α-Amylase Inhibition by Anthocyanin-3-Glucosides Using UV/Vis-Spectroscopy
3.2. Determination of α-Amylase Inhibition by Anthocyanin-3-Glucosides Using Isothermal Titration Calorimetry
3.3. Determination of α-Amylase Inhibition by Black Carrot Anthocyanins by UV/Vis-Spectroscopy
3.4. Determination of α-Amylase Inhibition by Anthocyanin-3-Glucosides Using Isothermal Titration Calorimetry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | c (µM) | λmax (nm) | ε for Each Concentration at λmax Mean ± SD (L/(cm · mol)) | ε Mean ± SD (L/(cm · mol)) | Value for ε Used (L/(cm · mol)) | Reference Value |
|---|---|---|---|---|---|---|
| Plg-3-glc | 9.79 15.66 19.58 39.16 | 497 | 23.017 ± 259 24.733 ± 227 24.209 ± 1302 25.709 ± 129 | 24.417 ± 1121 | 24.500 | 27.300 (L/cm · mol) in 1% aqueous HCl [36], 15.600 (L/cm · mol) in 0.2 M KCl [37,38] |
| Cyd-3-glc | 12.75 20.39 25.49 50.99 | 510 | 23.111 ± 1825 24.598 ± 1457 26.465 ± 1180 27.279 ± 453 | 25.363 ± 1974 | 25.500 | 26.900 L/(cm · mol) in 0.2 M KCl [37] |
| Dpd-3-glc | 11.58 18.53 23.16 46.32 | 516 | 25.999 ± 523 24.862 ± 1555 25.718 ± 296 25.495 ± 212 | 25.519 ± 484 | 25.500 | 29.000 L/(cm · mol) in pH 1 buffer (not stated what buffer was used (0.2 KCl) [39] |
| Peo-3-glc | 8.40 13.44 16.80 33.60 | 511 | 26.829 ± 802 25.939 ± 550 26.511 ± 624 25.420 ± 573 | 26.175 ± 623 | 26.000 | 11.300 L/(cm · cm) in 0.1% metanolic HCl [40] |
| Mlv-3-glc | 8.19 13.10 16.37 32.75 | 519 | 23.025 ± 2349 23.653 ± 1670 24.078 ± 743 25.496 ± 349 | 24.063 ± 1049 | 24.000 | 28.000 L/(cm · mol) in 0.1 mol/L HCl [38] |

| No. | m/z [MH]+ and Respective Fragments * | Structure | Assigned Anthocyanin Structure According to [7] | Proportion of Total Peak Area at 520 nm (%) | Absorption Coefficient ε at 520 nm, pH1 ** (M · cm) * |
|---|---|---|---|---|---|
| BC 1 | 743 581 450 287 | Cyd-trihex Cyd-dihex Cyd-hex Cyd | Cyd-gal-xyl-glc | 4.75 | 32.500 |
| BC 2 | 581 449 287 | Cyd-dihex Cyd-hex Cyd | Cyd-gal-xyl | 27.06 | 32.000 |
| BC 3 | 949 743 287 | Cyd-trihex(sin) Cyd-trihex Cyd | Cyd-gal-xyl-glc(sin) | 10.25 | 36.000 |
| BC 4 | 919 581 287 | Cyd-trihex(fer) Cyd-dihex Cyd | Cyd-gal-xyl-glc(fer) | 42.35 | 31.000 |
| BC 5 | - | Cyd-trihex(cum) | Cyd-gal-xyl-glc(cum) | 15.42 | 34.000 |
| - | - | BC-ACY | - | sum | average |
| - | - | - | - | 100 | 3250 |
| - | Compound | λmax (pH1) | ß (mg/L)a | Absorption Coefficient εa Mean ± SD | ß (mg/L)b | Absorption Coefficient εb Mean ± SD | ß (mg/L)c | Absorption Coefficient εc Mean ± SD | ß (mg/L)d | Absorption Coefficient εd Mean ± SD |
|---|---|---|---|---|---|---|---|---|---|---|
| Non-acylated anthocyanins | - | - | - | - | - | - | ||||
| BC 1 | A | - | 53.3 | 30,152.8 | - | - | - | - | - | - |
| B | - | 87.8 | 34,427.3 | - | - | - | - | - | - | |
| C | - | 89.7 | 32,900.0 | - | - | - | - | - | - | |
| average | 513 | - | 32,493.4 ± 2166.1 | - | - | - | - | - | - | |
| BC 2 | A | - | 355.7 | 31,297.4 | - | - | - | - | - | - |
| B | - | 351.3 | 33,040.2 | - | - | - | - | - | - | |
| average | 514 | - | 32,168.8 ± 1232.3 | - | - | - | - | - | - | |
| Acylated anthocyanins | - | - | - | - | - | - | ||||
| BC 3 | A | - | 127.0 ± 6.7 | - | 125.9 | 35,395.1 | 134.2 | 33,202.2 | 121.0 | 36,840.8 |
| B | - | 122.3 ± 3.0 | - | 121.6 | 36,898.3 | 125.6 | 35,731.7 | 119.7 | 37,492.1 | |
| - | - | - | - | - | 36,146.7 ± 1062.9 | - | 34,467.0 ± 1788.6 | - | 37,166.5 ± 460.5 | |
| average | 528 | - | 35,926.7 ± 1363.1 | - | - | - | - | - | - | |
| BC 4 | A | - | 908.2 ± 11.7 | - | 908.2 | 32,119.5 | 919.9 | 31,711.9 | 896.5 | 32,537.8 |
| B | - | 914.3 ± 20.8 | - | 912.7 | 30,296.2 | 935.8 | 29,548.4 | 894.3 | 30,919.7 | |
| - | - | - | - | - | 31,207.9 ± 1289.3 | 30,630.2 | 1529.8 ± 5.0 | - | 31,728.8 ± 1144.2 | |
| average | 526 | - | 31,188.9 ± 549.5 | - | - | - | - | - | - | |
| BC 5 | A | - | 250.4 ± 8.9 | - | 251.5 | 33,023.2 | 258.7 | 32,107.0 | 241.0 | 34,453.6 |
| B | - | 233.9 ± 4.4 | - | 236.3 | 34,734.8 | 228.9 | 35,862.0 | 236.6 | 34,696.7 | |
| - | - | - | - | - | 33,879.0 ± 1210.3 | -a | 33,984.5 ± 2655.2 | - | 34,575.2 ± 171.9 | |
| average | 525 | - | 34,146.2 ± 375.2 | - | - | - | - | - | - | |

| Inhibitor | R2 Mixed Inhibition According to Equation (4) | R2 Competitive Inhibition According to Equation (5) |
|---|---|---|
| Plg-3-glc | 0.99 ± 0.00 | 0.98 ± 0.01 |
| Cyd-3-glc | 0.97 ± 0.02 | 0.96 ± 0.03 |
| Dpd-3-glc | 0.98 ± 0.00 | 0.97 ± 0.00 |
| Peo-3-glc | 0.98 ± 0.02 | 0.97 ± 0.02 |

References
- de Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef] [PubMed]
- Anderson, O.M.; Jordheim, M. Basic Anthocyanin Chemistry and Dietary Sourcres. In Anthocyanins in Health and Disease; Wallace, T.C., Giusti, M.M., Eds.; CRC Press: Hoboken, NJ, USA, 2013; pp. 13–90. ISBN 978-1-4398-9471-2. [Google Scholar]
- Kammerer, D.; Carle, R.; Schieber, A. Quantification of anthocyanins in black carrot extracts (Daucus carota ssp. sativus var. atrorubens Alef.) and evaluation of their color properties. Eur. Food Res. Technol. 2004, 219, 479–486. [Google Scholar] [CrossRef]
- Algarra, M.; Fernandes, A.; Mateus, N.; de Freitas, V.; Esteves da Silva, J.C.G.; Casado, J. Anthocyanin profile and antioxidant capacity of black carrots (Daucus carota L. ssp. sativus var. atrorubens Alef.) from Cuevas Bajas, Spain. J. Food Compos. Anal. 2014, 33, 71–76. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Gläßgen, W.E.; Seitz, H.U.; Metzger, J.W. High-performance liquid chromatography/electrospray mass spectrometry and tandem mass spectrometry of anthocyanins from plant tissues and cell cultures of Daucus carota L. Biol. Mass Spectrom. 1992, 21, 271–277. [Google Scholar] [CrossRef]
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef]
- American Diabetes Association. The prevention or delay of type 2 diabetes. Diabetes Care 2002, 25, 742–749. [Google Scholar] [CrossRef]
- Hanefeld, M. The Role of Acarbose in the Treatment of Non–Insulin-Dependent Diabetes Mellitus. J. Diabetes Complicat. 1998, 12, 228–237. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-Anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef]
- Homoki, J.R.; Nemes, A.; Fazekas, E.; Gyémánt, G.; Balogh, P.; Gál, F.; Al-Asri, J.; Mortier, J.; Wolber, G.; Babinszky, L.; et al. Anthocyanin composition, antioxidant efficiency, and α-amylase inhibitor activity of different Hungarian sour cherry varieties (Prunus cerasus L.). Food Chem. 2016, 194, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Matsui, T.; Ueda, T.; Oki, T.; Sugita, K.; Terahara, N.; Matsumoto, K. alpha-Glucosidase inhibitory action of natural acylated anthocyanins. 1. Survey of natural pigments with potent inhibitory activity. J. Agric. Food Chem. 2001, 49, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Ostberg-Potthoff, J.J.; Berger, K.; Richling, E.; Winterhalter, P. Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism. Nutrients 2019, 11, 1166. [Google Scholar] [CrossRef]
- Kaeswurm, J.A.H.; Claasen, B.; Fischer, M.-P.; Buchweitz, M. Interaction of Structurally Diverse Phenolic Compounds with Porcine Pancreatic α-Amylase. J. Agric. Food Chem. 2019, 67, 11108–11118. [Google Scholar] [CrossRef]
- Soares, S.; Santos Silva, M.; García-Estévez, I.; Brandão, E.; Fonseca, F.; Ferreira-da-Silva, F.; Teresa Escribano-Bailón, M.; Mateus, N.; de Freitas, V. Effect of malvidin-3-glucoside and epicatechin interaction on their ability to interact with salivary proline-rich proteins. Food Chem. 2019, 276, 33–42. [Google Scholar] [CrossRef]
- Lehoczki, G.; Szabó, K.; Takács, I.; Kandra, L.; Gyémánt, G. Simple ITC method for activity and inhibition studies on human salivary α-amylase. J. Enzym. Inhib. Med. Chem. 2016, 31, 1648–1653. [Google Scholar] [CrossRef]
- Todd, M.J.; Gomez, J. Enzyme kinetics determined using calorimetry: A general assay for enzyme activity? Anal. Biochem. 2001, 296, 179–187. [Google Scholar] [CrossRef]
- Lachenmeier, D.W.; Teipel, J.; Scharinger, A.; Kuballa, T.; Walch, S.G.; Grosch, F.; Bunzel, M.; Okaru, A.O.; Schwarz, S. Fully automated identification of coffee species and simultaneous quantification of furfuryl alcohol using NMR spectroscopy. J. AOAC Int. 2020, in press. [Google Scholar] [CrossRef]
- Okaru, A.O.; Scharinger, A.; Rajcic de Rezende, T.; Teipel, J.; Kuballa, T.; Walch, S.G.; Lachenmeier, D.W. Validation of a Quantitative Proton Nuclear Magnetic Resonance Spectroscopic Screening Method for Coffee Quality and Authenticity (NMR Coffee Screener). Foods 2020, 9, 47. [Google Scholar] [CrossRef]
- Ackermann, S.M.; Dolsophon, K.; Monakhova, Y.B.; Kuballa, T.; Reusch, H.; Thongpanchang, T.; Bunzel, M.; Lachenmeier, D.W. Automated Multicomponent Analysis of Soft Drinks Using 1D 1H and 2D 1H-1H J-resolved NMR Spectroscopy. Food Anal. Methods 2017, 10, 827–836. [Google Scholar] [CrossRef]
- Yung-Chi, C.; Prusoff, W.H. Relationship between the inhibition constant (KI) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem. Pharmacol. 1973, 22, 3099–3108. [Google Scholar] [CrossRef]
- Bisswanger, H. Enzymkinetik. Theorie und Methoden; völlig neu bearb. Aufl.; Wiley-Interscience: Hoboken, NJ, USA, 2000; ISBN 9783527300969. [Google Scholar]
- D’Amico, S.; Sohier, J.S.; Feller, G. Kinetics and energetics of ligand binding determined by microcalorimetry: Insights into active site mobility in a psychrophilic alpha-amylase. J. Mol. Biol. 2006, 358, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Freyer, M.W.; Lewis, E.A. Isothermal Titration Calorimetry: Experimental Design, Data Analysis, and Probing Macromolecule/Ligand Binding and Kinetic Interactions. Methods Cell Biol. 2008, 84, 79–113. [Google Scholar] [CrossRef] [PubMed]
- Lo Piparo, E.; Scheib, H.; Frei, N.; Williamson, G.; Grigorov, M.; Chou, C.J. Flavonoids for controlling starch digestion: structural requirements for inhibiting human alpha-amylase. J. Med. Chem. 2008, 51, 3555–3561. [Google Scholar] [CrossRef] [PubMed]
- Wiese, S.; Gärtner, S.; Rawel, H.M.; Winterhalter, P.; Kulling, S.E. Protein interactions with cyanidin-3-glucoside and its influence on α-amylase activity. J. Sci. Food Agric. 2009, 89, 33–40. [Google Scholar] [CrossRef]
- Buchweitz, M. Natural Solutions for Blue Colors in Food. In Handbook on Natural Pigments in Food and Beverages: Industrial Applications for Improving Food Color; Carle, R., Schweiggert, R.M., Eds.; Woodhead Publishing is an imprint of Elsevier: Duxford, UK, 2016; pp. 355–384. ISBN 0081003714. [Google Scholar]
- Stintzing, F.C.; Stintzing, A.S.; Carle, R.; Frei, B.; Wrolstad, R.E. Color and antioxidant properties of cyanidin-based anthocyanin pigments. J. Agric. Food Chem. 2002, 50, 6172–6181. [Google Scholar] [CrossRef]
- Timberlake, C.F.; Bridle, P. Flavylium salts, anthocyanidins and anthocyanins. I. Structural transformations in acid solutions. J. Sci. Food Agric. 1967, 18, 473–478. [Google Scholar] [CrossRef]
- Sondheimer, E. On the Relation between Spectral Changes and pH of the Anthocyanin Pelargonidin 3-Monoglucoside 1. J. Am. Chem. Soc. 1953, 75, 1507–1508. [Google Scholar] [CrossRef]
- Figueiredo, P.; Elhabiri, M.; Toki, K.; Saito, N.; Dangles, O.; Brouillard, R. New aspects of anthocyanin complexation. Intramolecular copigmentation as a means for colour loss? Phytochemistry 1996, 41, 301–308. [Google Scholar] [CrossRef]
- Sadilova, E.; Stintzing, F.C.; Carle, R. Thermal Degradation of Acylated and Nonacylated Anthocyanins. J. Food Sci. 2006, 71, C504–C512. [Google Scholar] [CrossRef]
- Matsui, T.; Ueda, T.; Oki, T.; Sugita, K.; Terahara, N.; Matsumoto, K. α-Glucosidase Inhibitory Action of Natural Acylated Anthocyanins. 2. α-Glucosidase Inhibition by Isolated Acylated Anthocyanins. J. Agric. Food Chem. 2001, 49, 1952–1956. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, E.C.; Geissman, T.A. The chemistry of flower pigmentation in Antirrhinum majus color genotypes. III. Relative anthocyanin and aurone concentrations. Arch. Biochem. Biophys. 1955, 55, 389–402. [Google Scholar] [CrossRef]
- Sadilova, E.; Carle, R.; Stintzing, F.C. Thermal degradation of anthocyanins and its impact on color and in vitro antioxidant capacity. Mol. Nutr. Food Res. 2007, 51, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Rodríguez-Saona, L.E.; Wrolstad, R.E. Molar absorptivity and color characteristics of acylated and non-acylated pelargonidin-based anthocyanins. J. Agric. Food Chem. 1999, 47, 4631–4637. [Google Scholar] [CrossRef] [PubMed]
- Boulekbache-Makhlouf, L.; Medouni, L.; Medouni-Adrar, S.; Arkoub, L.; Madani, K. Effect of solvents extraction on phenolic content and antioxidant activity of the byproduct of eggplant. Ind. Crops Prod. 2013, 49, 668–674. [Google Scholar] [CrossRef]
- Somers, T.C. Grape phenolics: The anthocyanins ofVitis vinifera, variety shiraz. J. Sci. Food Agric. 1966, 17, 215–219. [Google Scholar] [CrossRef]







| - | (i) Inhibition Constants Obtained by the Global Fit (Equation (4)) | (ii) Independent Fit for Vmax/Vmaxapp and Km and Kmapp (Equation (6), Michaelis Menten, Table S2) and Calculation of the Inhibition Constants (Equations (7) and (8)) | ||||
|---|---|---|---|---|---|---|
| Inhibitor | Kic (µM) | Kiu (µM) | Kic/Kiu | Kic (µM) | Kiu (µM) | Kic/Kiu |
| Plg-glc | 57 ± 15 a | 151 ± 59 a | 0.39 ± 0.05 a | 69 ± 1 a | 158 ± 53 a | 0.46 ± 0.13 a |
| Cyd-glc | 81 ± 10 ***a | 184 ± 56 ***a | 0.45 ± 0.08 ***a | 134 *a | 228 *a | 0.56 *a |
| Dpd-glc | 62 ± 25 a | 204 ± 68 a | 0.34 ± 0.24 a | 63 ± 19 a | 270 ± 33 a | 0.26 ± 0.13 a |
| Peo-glc | 109 ± 39 a | 258 ± 78 a | 0.42 ± 0.23 a | 118 ± 42 **a | 254 ± 82 **a | 0.43 ± 0.04 **a |
| Mlv-glc # | 44 ± 4 a | 143 ± 44 a | 0.32 ± 0.07 a | 69 ± 17 a | 147 ± 50 a | 0.58 ± 0.35 a |
| - | (i) Kic and Kiu Obtained by the Global Fit (Equation (4)) | (ii) Independent Fit for vmax/vmaxapp and Km and Kmapp (Equation (6), Michaelis Menten) and Calculation of Kic and Kiu by Equations (7) and (8) | ||||||
|---|---|---|---|---|---|---|---|---|
| I | [S] µM | [S] µM | ||||||
| - | 10 | 100 | 103 | 106 | 10 | 100 | 103 | 106 |
| Plg-glc | 58 ± 15 a | 71 ± 20 a | 117 ± 41 ab | 151 ± 59 a | 71 ± 2 a | 84 ± 7 a | 127 ± 31 a | 158 ± 53 a |
| Cyd-glc | 83 ± 11 ***a | 97 ± 15 ***a | 147 ± 36 ***ab | 184 ± 56 ***a | 137 *a | 153 *a | 201 *a | 228 *a |
| Dpd-glc | 64 ± 25 a | 77 ± 25 a | 134 ± 1 ab | 203 ± 68 a | 65 ± 20 **a | 83 ± 24 **a | 167 ± 20 **a | 270 ± 32 **a |
| Peo-glc | 112 ± 40 a | 134 ± 46 a | 209 ± 65 b | 258 ± 78 a | 121 ± 42 a | 142 ± 49 a | 212 ± 69 a | 254 ± 82 a |
| Mlv-glc # | 46 ± 4 a | 58 ± 7 a | 106 ± 24 a | 143 ± 44 a | 71 ± 16 a | 83 ± 10 a | 121 ± 21 a | 147 ± 50 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaeswurm, J.A.H.; Könighofer, L.; Hogg, M.; Scharinger, A.; Buchweitz, M. Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides. Foods 2020, 9, 367. https://doi.org/10.3390/foods9030367
Kaeswurm JAH, Könighofer L, Hogg M, Scharinger A, Buchweitz M. Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides. Foods. 2020; 9(3):367. https://doi.org/10.3390/foods9030367
Chicago/Turabian StyleKaeswurm, Julia A. H., Lisa Könighofer, Melanie Hogg, Andreas Scharinger, and Maria Buchweitz. 2020. "Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides" Foods 9, no. 3: 367. https://doi.org/10.3390/foods9030367
APA StyleKaeswurm, J. A. H., Könighofer, L., Hogg, M., Scharinger, A., & Buchweitz, M. (2020). Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine α-Amylase by Anthocyanin-3-Glycosides. Foods, 9(3), 367. https://doi.org/10.3390/foods9030367