Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives

Abstract
1. The Importance of Bread in the Human Diet
2. RS Definition and Analytical Methods
3. Natural RS Ingredients in Bread-Making and Structural Basis of Their Resistant Digestion
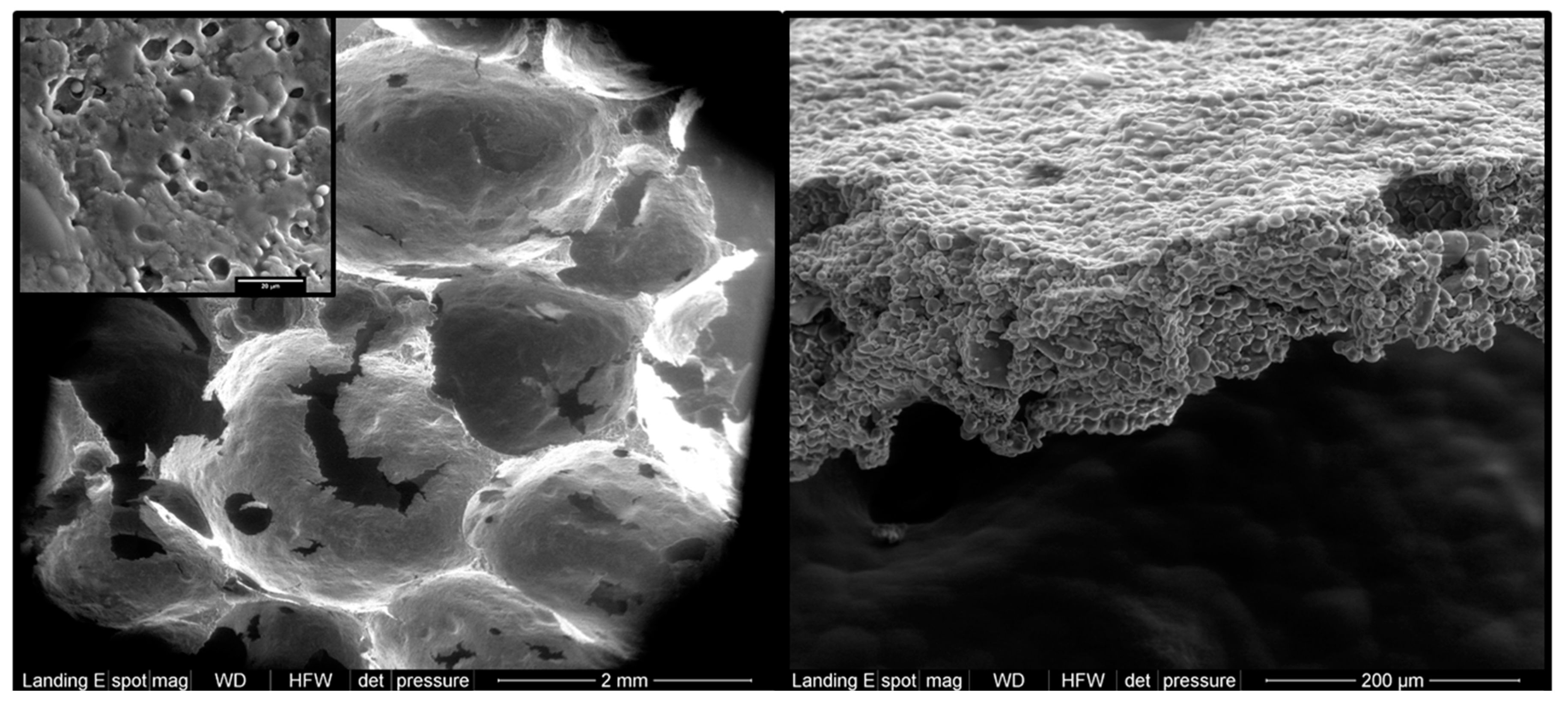
3.1. Physical Barriers Comprising Plant Cell Walls and/or the Food Matrix (RS1)
3.2. Granular Surface Properties (Granular Resistant Starch, RS2)
3.3. Dispersed Starch Molecules Forming Resistant Starch upon Cooling and Storage (RS3)
3.4. Introduction of Chemical Structures (Chemically Modified Resistant Starch, RS4)
3.5. Lipid Complexed Resistant Starch (RS5)
4. Effects of RS in Physicochemical Characteristics of the Breads
5. Current Legislation of RS Ingredients and Products in the Food Industry
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lafiandra, D.; Riccardi, G.; Shewry, P.R. Improving cereal grain carbohydrates for diet and health. J. Cereal Sci. 2014, 59, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Cust, A.E.; Skilton, M.R.; van Bakel, M.M.; Halkjaer, J.; Olsen, A.; Agnoli, C.; Psaltopoulou, T.; Buurma, E.; Sonestedt, E.; Chirlaque, M.D.; et al. Total dietary carbohydrate, sugar, starch and fibre intakes in the European Prospective Investigation into Cancer and Nutrition. Eur. J. Clin. Nutr. 2009, 63, S37–S60. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Norat, T. Whole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: Systematic review and dose-response meta-analysis of prospective studies. BMJ 2016, 353, i2716. [Google Scholar] [CrossRef] [PubMed]
- Pot, G.K.; Prynne, C.J.; Almoosawi, S.; Kuh, D.; Stephen, A.M. Trends in food consumption over 30 years: Evidence from a British birth cohort. Eur. J. Clin. Nutr. 2015, 69, 817. [Google Scholar] [CrossRef] [PubMed]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch–a review. Compr. Rev. Food Sci. F. 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Martinez, M.M.; Roman, L.; Gomez, M. Implications of hydration depletion in the in vitro starch digestibility of white bread crumb and crust. Food Chem. 2018, 239, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Thorvaldsson, K.; Skjöldebrand, C. Water diffusion in bread during baking. LWT 1998, 31, 658–663. [Google Scholar] [CrossRef]
- Primo-Martín, C.; van Nieuwenhuijzen, N.H.; Hamer, R.J.; van Vliet, T. Crystallinity changes in wheat starch during the bread-making process: Starch crystallinity in the bread crust. J. Cereal Sci. 2007, 45, 219–226. [Google Scholar] [CrossRef]
- Roman, L.; Gomez, M.; Hamaker, B.R.; Martinez, M.M. Banana starch and molecular shear fragmentation dramatically increase structurally driven slowly digestible starch in fully gelatinized bread crumb. Food Chem. 2019, 274, 664–671. [Google Scholar] [CrossRef]
- Ceriello, A.; Colagiuri., S. International Diabetes Federation guideline for management of postmeal glucose: A review of recommendations. Diabetic Med. 2008, 25, 1151–1156. [Google Scholar] [CrossRef]
- Brand-Miller, J.; McMillan-Price, J.; Steinbeck, K.; Caterson, I. Dietary glycemic index: Health implications. J. Am. Coll. Nutr. 2009, 28, 446S–449S. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Kendall, C.W.; Augustin, L.S.; Franceschi, S.; Hamidi, M.; Marchie, A.; Jenkins, A.L.; Axelsen, M. Glycemic index: Overview of implications in health and disease. Am. J. Clin. Nutr. 2002, 76, 266S–273S. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. The Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef]
- Bindels, L.B.; Walter, J.; Ramer-Tait, A.E. Resistant starches for the management of metabolic diseases. Curr. Opin. Clin. Nutr. 2015, 18, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Schalinske, M.R.K.; et al. Resistant Starch: Promise for Improving Human Health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef]
- Bird, A.R.; Lopez-Rubio, A.; Shrestha, A.K.; Gidley, M.J. Resistant starch in vitro and in vivo: Factors determining yield, structure, and physiological relevance. In Modern Biopolymer Science; Kasapis, S., Ubbink, J., Eds.; IT Norton, Academic Press: London, UK, 2009; pp. 449–510. [Google Scholar]
- Bird, A.; Conlon, M.; Christophersen, C.; Topping, D. Resistant starch, large bowel fermentation and a broader perspective of prebiotics and probiotics. Benef. Microbes 2010, 1, 423–431. [Google Scholar] [CrossRef]
- Dupuis, J.H.; Liu, Q.; Yada, R.Y. Methodologies for increasing the resistant starch content of food starches: A review. Compr. Rev. Food Sci. F. 2014, 13, 1219–1234. [Google Scholar] [CrossRef]
- Fuentes-Zaragoza, E.; Sánchez-Zapata, E.; Sendra, E.; Sayas, E.; Navarro, C.; Fernández-López, J.; Pérez-Alvarez, J.A. Resistant starch as prebiotic: A review. Starch-Stärke 2011, 63, 406–415. [Google Scholar] [CrossRef]
- Higgins, J.A.; Brown, I.L. Resistant starch: A promising dietary agent for the prevention/treatment of inflammatory bowel disease and bowel cancer. Curr. Opin. Gastroenterol. 2013, 29, 190–194. [Google Scholar] [CrossRef]
- Keenan, M.J.; Zhou, J.; Hegsted, M.; Pelkman, C.; Durham, H.A.; Coulon, D.B.; Martin, R.J. Role of resistant starch in improving gut health, adiposity, and insulin resistance. Adv. Nutr. 2015, 6, 198–205. [Google Scholar] [CrossRef]
- Koh, G.Y.; Rowling, M.J. Resistant starch as a novel dietary strategy to maintain kidney health in diabetes mellitus. Nutr. Rev. 2017, 75, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, S.; Nugent, A.P. Health effects of resistant starch. Nutr. Bull. 2017, 42, 10–41. [Google Scholar] [CrossRef]
- Qi, X.; Tester, R.F. Utilisation of dietary fibre (non-starch polysaccharide and resistant starch) molecules for diarrhoea therapy: A mini-review. Int. J. Biol. Macromol. 2019, 122, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Raigond, P.; Ezekiel, R.; Raigond, B. Resistant starch in food: A review. J. Sci. Food Agric. 2015, 1968–1978. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.D.; Bickerton, A.S.; Dennis, A.L.; Vidal, H.; Frayn, K.N. Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adipose tissue metabolism. Am. J. Clin. Nutr. 2005, 82, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.C.; Meng, Y.; Bai, H.; Shen, D.Q.; Wan, B.C.; Chen, L.Y. Meta-analysis indicates that resistant starch lowers serum total cholesterol and low-density cholesterol. Nutr. Res. 2018, 54, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Champ, M. Resistant starch. In Starch in Food: Structure, Function and Applications; Eliasson, A.C., Ed.; CRC Press: New York, NY, USA, 2004; pp. 560–574. [Google Scholar]
- Berry, C.S. Resistant starch formation: Formation and measurement of starch that survives exhaustive digestion with amylolytic enzymes during the determination of dietary fibre. J. Cereal Sci. 1986, 4, 301–314. [Google Scholar] [CrossRef]
- Englyst, H.; Wiggins, H.S.; Cummings, J.H. Determination of the non-starch polysaccharides in plant foods by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1982, 107, 307–318. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar]
- Englyst, H.N.; Kingman, S.M.; Hudson, G.J.; Cummings, J.H. Measurement of resistant starch in vitro and in vivo. Br. J. Nutr. 1996, 75, 749–755. [Google Scholar] [CrossRef]
- Englyst, K.N.; Hudson, G.J.; Englyst, H.N. Starch analysis in food. In Encyclopedia of Analytical Chemistry; Meyers, R.A., Ed.; John Wiley & Sons: Chichester, UK, 2000; pp. 4246–4262. [Google Scholar]
- McCleary, B.V.; Monaghan, D.A. Measurement of resistant starch. J. AOAC Int. 2002, 85, 665–675. [Google Scholar] [PubMed]
- AOAC. AOAC Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- AACC International. AACC International Approved Methods; AACC International: St. Paul, MN, USA, 2015. [Google Scholar]
- Akerberg, A.K.; Liljeberg, H.G.; Granfeldt, Y.E.; Drews, A.W.; Björck, I.M. An in vitro method, based on chewing, to predict resistant starch content in foods allows parallel determination of potentially available starch and dietary fiber. J. Nutr. 1998, 128, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Champ, M. Determination of resistant starch in foods and food products: Interlaboratory study. Eur. J. Clin. Nutr. 1992, 46, S51–S62. [Google Scholar] [PubMed]
- Champ, M.; Martin, L.; Noah, L.; Gratas, M. Analytical methods for resistant starch. In Complex Carbohydrates in Foods; Cho, S.S., Prosky, L., Dreher, M., Eds.; Marcel Dekker Inc: New York, NY, USA, 1999; pp. 169–187. [Google Scholar]
- Goni, I.; Garcia-Diz, L.; Manas, E.; Saura-Calixto, F. Analysis of Resistant Starch: A Method for Foods and Food Products. Food Chem. 1996, 56, 445–449. [Google Scholar] [CrossRef]
- Muir, J.G.; O’Dea, K. Validation of an in vitro assay for predicting the amount of starch that escapes digestion in the small intestine of humans. Am. J. Clin. Nutr. 1993, 57, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Prosky, L.; Asp, N.G.; Furda, I.; DeVries, J.W.; Schweizer, T.F.; Harland, B.F. Determination of total dietary fiber in foods and food products: Collaborative study. J. Assoc. Off. Anal. Chem. 1985, 68, 677–679. [Google Scholar] [PubMed]
- Lee, S.C.; Prosky, L. International survey on dietary fiber: Definition, analysis, and reference materials. J. AOAC. Int. 1995, 78, 22–36. [Google Scholar]
- McCleary, B.V. An integrated procedure for the measurement of total dietary fibre (including resistant starch), non-digestible oligosaccharides and available carbohydrates. Anal. Bioanal. Chem. 2007, 389, 291–308. [Google Scholar] [CrossRef]
- McCleary, B.V.; De Vries, J.W.; Rader, J.I.; Cohen, G.; Prosky, L.; Mugford, D.C.; Champ, M.; Okuma, K. Determination of total dietary fiber (CODEX definition) by enzymatic-gravimetric method and liquid chromatography: Collaborative study. J. AOAC Int. 2010, 93, 221–233. [Google Scholar] [CrossRef]
- Eerlingen, R.C.; Delcour, J.A. Formation, analysis, structure, and properties of type III enzyme resistant starch. J. Cereal Sci. 1995, 22, 129–138. [Google Scholar] [CrossRef]
- Brown, I.L.; Yotsuzuka, M.; Birkett, A.; Henriksson, A. Prebiotics, synbiotics and resistant starch. J. Japanese. Assoc. Dietary Fiber Res. 2006, 10, 1–9. [Google Scholar]
- Amaral, O.; Guerreiro, C.S.; Gomes, A.; Cravo, M. Resistant starch production in wheat bread: Effect of ingredients, baking conditions and storage. Eur Food Res Technol. 2016, 242, 1747–1753. [Google Scholar] [CrossRef]
- Liljeberg, H.; Akerberg, A.; Bjorck, I. Resistant starch formation in bread as influenced by choice of ingredients or baking conditions. Food Chem. 1996, 56, 389–394. [Google Scholar] [CrossRef]
- Wen, Q.B.; Lorenz, K.J.; Martin, D.J.; Stewart, B.G.; Sampson, D.A. Carbohydrate Digestibility and Resistant Starch of Steamed Bread. Starch-Stärke 1996, 48, 180–185. [Google Scholar] [CrossRef]
- Dhital, S.; Gidley, M.J.; Warren, F.J. Inhibition of α-amylase activity by cellulose: Kinetic analysis and nutritional implications. Carbohydr. Polym. 2015, 123, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, R.R.; Dhital, S.; Mense, A.; Gidley, M.J.; Shi, Y.C. Intact cellular structure in cereal endosperm limits starch digestion in vitro. Food. Hydrocoll. 2018, 81, 139–148. [Google Scholar] [CrossRef]
- Bhattarai, R.R.; Dhital, S.; Wu, P.; Chen, X.D.; Gidley, M.J. Digestion of isolated legume cells in a stomach-duodenum model: Three mechanisms limit starch and protein hydrolysis. Food Funct. 2017, 8, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Dhital, S.; Bhattarai, R.R.; Gorham, J.; Gidley, M.J. Intactness of cell wall structure controls the in vitro digestion of starch in legumes. Food Funct. 2016, 7, 1367–1379. [Google Scholar] [CrossRef]
- Edwards, C.H.; Grundy, M.M.; Grassby, T.; Vasilopoulou, D.; Frost, G.S.; Butterworth, P.J.; Berry, S.E.E.; Sanderson, J.; Ellis, P.R. Manipulation of starch bioaccessibility in wheat endosperm to regulate starch digestion, postprandial glycemia, insulinemia, and gut hormone responses: A randomized controlled trial in healthy ileostomy participants. Am. J. Clin. Nutr. 2015, 102, 791–800. [Google Scholar] [CrossRef]
- Livesey, G.; Wilkinson, J.A.; Roe, M.; Faulks, R.; Clark, S.; Brown, J.C.; Kennedy, H.; Elia, M. Influence of the physical form of barley grain on the digestion of its starch in the human small intestine and implications for health. Am. J. Clin. Nutr. 1995, 61, 75–81. [Google Scholar] [CrossRef]
- Roman, L.; Gomez, M.; Li, C.; Hamaker, B.R.; Martinez, M.M. Biophysical features of cereal endosperm that decrease starch digestibility. Carbohydr. Polym. 2017, 165, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Birkett, A.M.; Brown, I.L. Resistant starch and health. In Technology of Functional Cereal Products; Hamaker, B.R., Ed.; CRC Press: West Palm Beach, FL, USA, 2008; pp. 63–85. [Google Scholar]
- Al-Rabadi, G.J.S.; Gilbert, R.G.; Gidley, M.J. Effect of particle size on kinetics of starch digestion in milled barley and sorghum grains by porcine alpha amylase. J. Cereal Sci. 2009, 50, 198–204. [Google Scholar] [CrossRef]
- Granfeldt, Y.; Liljeberg, H.; Drews, A.; Newman, R.; Björck, I. Glucose and insulin responses to barley products: Influence of food structure and amylose-amylopectin ratio. Am. J.Clin..Nutr. 1994, 59, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Wesson, V.; Wolever, T.M.; Jenkins, A.L.; Kalmusky, J.; Guidici, S.; Csima, A.; Josse, R.G.; Wong, G.S. Wholemeal versus wholegrain breads: Proportion of whole or cracked grain and the glycaemic response. BMJ 1988, 297, 958–960. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.M.; Calviño, A.; Rosell, C.M.; Gomez, M. Effect of different extrusion treatments and particle size distribution on the physicochemical properties of rice flour. Food Bioprocess.Tech. 2014, 7, 2657–2665. [Google Scholar] [CrossRef]
- De La Hera, E.; Rosell, C.M.; Gomez, M. Effect of water content and flour particle size on gluten-free bread quality and digestibility. Food Chem. 2014, 151, 526–531. [Google Scholar] [CrossRef]
- Protonotariou, S.; Mandala, I.; Rosell, C.M. Jet milling effect on functionality, quality and in vitro digestibility of whole wheat flour and bread. Food Bioprocess. Tech. 2015, 8, 1319–1329. [Google Scholar] [CrossRef]
- Ezeogu, L.I.; Duodu, K.G.; Taylor, J.R.N. Effects of endosperm texture and cooking conditions on the in vitro starch digestibility of sorghum and maize flours. J. Cereal. Sci. 2005, 42, 33–44. [Google Scholar] [CrossRef]
- Zhang, G.; Hamaker, B.R. Low α-amylase starch digestibility of cooked sorghum flours and the effect of protein. Cereal. Chem. 1998, 75, 710–713. [Google Scholar] [CrossRef]
- Berti, C.; Riso, P.; Monti, L.D.; Porrini, M. In vitro starch digestibility and in vivo glucose response of gluten-free foods and their gluten counterparts. Eur. J. Nutr. 2004, 43, 198–204. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Thorne, M.J.; Wolever, T.M.; Jenkins, A.L.; Rao, A.V.; Thompson, L.U. The effect of starch-protein interaction in wheat on the glycemic response and rate of in vitro digestion. Am. J. Clin. Nutr. 1987, 45, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.R.; Taylor, J.; Campanella, O.H.; Hamaker, B.R. Functionality of the storage proteins in gluten-free cereals and pseudocereals in dough systems. J. Cereal. Sci. 2016, 67, 22–34. [Google Scholar] [CrossRef]
- Adams, C.A.; Novellie, L.; Liebenberg, N.V.D.W. Biochemical properties and ultrastructure of protein bodies isolated from selected cereals. Cereal. Chem. 1976, 53, 1–12. [Google Scholar]
- Batterman-Azcona, S.J.; Lawton, J.W.; Hamaker, B.R. Effect of specific mechanical energy on protein bodies and a-zeins in corn flour extrudates. Cereal. Chem. 1999, 76, 316–320. [Google Scholar] [CrossRef]
- Batterman-Azcona, S.J.; Hamaker, B.R. Changes occurring in protein body structure and a-zein during cornflake processing. Cereal. Chem. 1998, 75, 217–221. [Google Scholar] [CrossRef]
- Vamadevan, V.; Bertoft, E. Structure-function relationships of starch components. Starch-Stärke 2015, 67, 55–68. [Google Scholar] [CrossRef]
- Inan Eroglu, E.; Buyuktuncer, Z. The effect of various cooking methods on resistant starch content of foods. Nutr. Food Sci. 2017, 47, 522–533. [Google Scholar] [CrossRef]
- Faisant, N.; Buleon, A.; Colonna, P.; Molis, C.; Lartigue, S.; Galmiche, J.P.; Champ, M. Digestion of raw banana starch in the small intestine of healthy humans: Structural features of resistant starch. Br. J. Nutr. 1995, 73, 111–123. [Google Scholar] [CrossRef]
- Takeda, C.; Takeda, Y.; Hizukuri, S. Structure of the amylopectin fraction of amylomaize. Carbohydr. Res. 1993, 246, 273–281. [Google Scholar] [CrossRef]
- Planchot, V.; Colonna, P. Purification and characterization of extracellular alpha-amylase from Aspergillus fumigatus. Carbohydr. Res. 1995, 272, 97–109. [Google Scholar] [CrossRef]
- Gallant, D.J.; Bouchet, B.; Baldwin, P.M. Microscopy of starch: Evidence of a new level of granule organization. Carbohydr. Polym. 1997, 32, 177–191. [Google Scholar] [CrossRef]
- Dhital, S.; Warren, F.J.; Butterworth, P.J.; Ellis, P.R.; Gidley, M.J. Mechanisms of starch digestion by α-amylase—Structural basis for kinetic properties. Crit. Rev. Food Sci. Nutr. 2017, 57, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Haralampu, S. Resistant starch–a review of the physical properties and biological impact of RS3. Carbohydr. Polym. 2000, 41, 285–292. [Google Scholar] [CrossRef]
- Jacobs, H.; Delcour, J. Hydrothermal modifications of granular starch with retention of the granular structure: A review. J. Agric. Food Chem. 1998, 46, 2895–2905. [Google Scholar] [CrossRef]
- Htoon, A.; Shrestha, A.K.; Flanagan, B.M.; Lopez-Rubio, A.; Bird, A.R.; Gilbert, E.P.; Gidley, M.J. Effects of processing high amylose maize starches under controlled conditions on structural organization and amylase digestibility. Carbohydr. Polym. 2009, 75, 236–245. [Google Scholar] [CrossRef]
- Patel, H.; Royall, P.G.; Gaisford, S.; Williams, G.R.; Edwards, C.H.; Warren, F.J.; Flanagan, B.M.; Ellis, P.R.; Butterworth, P.J. Structural and enzyme kinetic studies of retrograded starch: Inhibition of α-amylase and consequences for intestinal digestion of starch. Carbohydr. Polym. 2017, 164, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Englyst, H.N.; Cummings, J.H. Digestion of the polysaccharides of some cereal foods in the human small intestine. Am. J. Clin. Nutr. 1985, 42, 778–787. [Google Scholar] [CrossRef]
- Vasanthan, T.; Bhatty, R.S. Enhancement of resistant starch (RS3) in amylomaize, barley, field pea and lentil starches. Starch-Stärke 1998, 50, 286–289. [Google Scholar] [CrossRef]
- Lopez-Rubio, A.; Flanagan, B.M.; Shrestha, A.K.; Gidley, M.J.; Gilbert, E.P. Molecular rearrangement of starch during in vitro digestion: Toward a better understanding of enzyme resistant starch formation in processed starches. Biomacromolecules 2008, 9, 1951–1958. [Google Scholar] [CrossRef]
- Cairns, P.; Sun, L.; Morris, V.J.; Ring, S.G. Physicochemical studies using amylose as an in vitro model for resistant starch. J. Cereal Sci. 1995, 21, 37–47. [Google Scholar] [CrossRef]
- Gidley, M.J.; Cooke, D.; Darke, A.H.; Hoffmann, R.A.; Russell, A.L.; Greenwell, P. Molecular order and structure in enzyme-resistant retrograded starch. Carbohydr. Polym. 1995, 28, 23–31. [Google Scholar] [CrossRef]
- Lopez-Rubio, A.; Htoon, A.; Gilbert, E.P. Influence of extrusion and digestion on the nanostructure of high-amylose maize starch. Biomacromolecules 2007, 8, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Dhital, S.; Flanagan, B.M.; Luckman, P.; Halley, P.J.; Gidley, M.J. Extrusion induced low-order starch matrices: Enzymic hydrolysis and structure. Carbohydr. Polym. 2015, 134, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Roman, L.; Campanella, O.; Martinez, M.M. Shear-induced molecular fragmentation decreases the bioaccessibility of fully gelatinized starch and its gelling capacity. Carbohydr. Polym. 2019, 215, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sofyan, M.; Hamaker, B.R. Slowly digestible state of starch: Mechanism of slow digestion property of gelatinized maize starch. J. Agric. Food Chem. 2008, 56, 4695–4702. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.M.; Li, C.; Okoniewska, M.; Mukherjee, N.; Vellucci, D.; Hamaker, B.R. Slowly digestible starch in fully gelatinized material is structurally driven by molecular size and A and B1 chain lengths. Carbohydr. Polym. 2018, 197, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Englyst, H.N.; Macfarlane, G.T. Breakdown of resistant and readily digestible starch by human gut bacteria. J. Sci. Food Agric. 1986, 37, 699–706. [Google Scholar] [CrossRef]
- Benmoussa, M.; Moldenhauer, K.A.; Hamaker, B.R. Rice amylopectin fine structure variability affects starch digestion properties. J. Agric. Food Chem. 2007, 55, 1475–1479. [Google Scholar] [CrossRef]
- Klucinec, J.D.; Thompson, D.B. Amylose and amylopectin interact in retrogradation of dispersed high-amylose starches. Cereal. Chem. 1999, 76, 282–291. [Google Scholar] [CrossRef]
- Matalanis, AM.; Campanella, O.H.; Hamaker, B.R. Storage retrogradation behavior of sorghum, maize and rice starch pastes related to amylopectin fine structure. J. Cereal Sci. 2009, 50, 74–81. [Google Scholar] [CrossRef]
- Maningat, C.C.; Seib, P.A. RS4-type resistant starch: Chemistry, functionality and health benefits. In Resistant Starch Sources, Applications and Health Benefits; Shi, Y.C., Maningat, C.C., Eds.; John Wiley: Hoboken, NJ, USA, 2013; pp. 43–77. [Google Scholar]
- Ingredion. Official website. Technical documents for Versafibe 1490 from Ingredion website. Available online: https://www.ingredion.us/content/dam/ingredion/technical-documents/na/VERSAFIBE%201490%20%20%2006400400%20%20%20Technical%20Specification.pdf (accessed on 26 June 2019).
- Roquette. Official website. Modified starches under the name “CLEARGUM”. Available online: https://www.roquette.com/product-finder-food-nutrition/ (accessed on 25 June 2019).
- MGP. Official website. Technical information on Fibersym RS4. Available online: https://www.mgpingredients.com/food-ingredients/products/line/fibersym (accessed on 26 June 2019).
- Cargill. Official website. Technical information on RS4 starches C☆PolarTex, C☆StabiTex, C☆Tex. Available online: https://www.cargill.com/food-bev/emea/starches-derivatives/stabilized-starches (accessed on 26 June 2019).
- Arp, C.G.; Correa, M.J.; Zuleta, Á.; Ferrero, C. Techno-functional properties of wheat flour-resistant starch mixtures applied to breadmaking. Int. J. Food Sci. Technol. 2017, 52, 550–558. [Google Scholar] [CrossRef]
- Lee Yeo, L.; Seib, P.A. White pan bread and sugar-snap cookies containing wheat starch phosphate, a cross-linked resistant starch. Cereal. Chem. 2009, 86, 210–220. [Google Scholar]
- Miller, R.A.; Bianchi, E. Effect of RS4 resistant starch on dietary fiber content of white pan bread. Cereal. Chem. 2017, 94, 185–189. [Google Scholar] [CrossRef]
- Miyazaki, M.; Van Hung, P.; Maeda, T.; Morita, N. Recent advances in application of modified starches for breadmaking. Trends. Food Sci. Technol. 2006, 17, 591–599. [Google Scholar] [CrossRef]
- Sciarini, L.S.; Bustos, M.C.; Vignola, M.B.; Paesani, C.; Salinas, C.N.; Perez, G.T. A study on fibre addition to gluten free bread: Its effects on bread quality and in vitro digestibility. J. Food Sci. Technol. 2017, 54, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P.; Morita, N. Dough properties and bread quality of flours supplemented with cross-linked corn starches. Food Res. Int. 2004, 37, 461–467. [Google Scholar] [CrossRef]
- Wepner, B.; Berghofer, E.; Miesenberger, E.; Tiefenbacher, K.; NK Ng, P. Citrate starch—application as resistant starch in different food systems. Starch-Stärke 1999, 51, 354–361. [Google Scholar] [CrossRef]
- Witczak, M.; Juszczak, L.; Ziobro, R.; Korus, J. Influence of modified starches on properties of gluten-free dough and bread. Part I: Rheological and thermal properties of gluten-free dough. Food Hydrocoll. 2012, 28, 353–360. [Google Scholar] [CrossRef]
- Ziobro, R.; Korus, J.; Witczak, M.; Juszczak, L. Influence of modified starches on properties of gluten-free dough and bread. Part II: Quality and staling of gluten-free bread. Food Hydrocoll. 2012, 29, 68–74. [Google Scholar] [CrossRef]
- Larsson, K.; Miezis, Y. On the possibility of dietary fiber formation by interaction in the intestine between starch and lipids. Starch-Stärke 1979, 31, 301–302. [Google Scholar] [CrossRef]
- Morrison, W.R.; Law, R.V.; Snape, C.E. Evidence for Inclusion Complexes of Lipids with V-amylose in Maize, Rice and Oat Starches. J. Cereal Sci. 1993, 18, 107–109. [Google Scholar] [CrossRef]
- Ai, Y.; Hasjim, J.; Jane, J. Effects of lipids on enzymatic hydrolysis and physical properties of starch. Carbohydr. Polym. 2013, 92, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Takato, S.; Sasaki, T.; Kajiwara, K. Complex formation, thermal properties, and in-vitro digestibility of gelatinized potato starch-fatty acid mixtures. Food Hydrocoll. 2012, 27, 228–234. [Google Scholar] [CrossRef]
- Lau, E.; Zhou, W.; Henry, C.J. Effect of fat type in baked bread on amylose–lipid complex formation and glycaemic response. Br. J. Nutr. 2016, 115, 2122–2129. [Google Scholar] [CrossRef] [PubMed]
- Biliaderis, C. Structural transitions and related physical properties of starch. In Starch. Chemistry and Technology; BeMiller, J., Whistler, R., Eds.; Academic Press: New York, NY, USA, 2009; pp. 293–372. [Google Scholar]
- Schirmer, M.; Jekle, M.; Becker, T. Starch gelatinization and its complexity for analysis. Starch-Stärke 2015, 67, 30–41. [Google Scholar] [CrossRef]
- Roman, L.; Belorio, M.; Gomez, M. Gluten-Free Breads: The Gap Between Research and Commercial Reality. Compr. Rev. Food Sci. F. 2019, 18, 690–702. [Google Scholar] [CrossRef]
- Pareyt, B.; Finnie, S.M.; Putseys, J.A.; Delcour, J.A. Lipids in bread making: Sources, interactions, and impact on bread quality. J. Cereal. Sci. 2011, 54, 266–279. [Google Scholar] [CrossRef]
- Hasjim, J.; Lee, SO.; Hendrich, S.; Setiawan, S.; Ai, Y.; Jane, J.L. Characterization of a novel resistant-starch and its effects on postprandial plasma-glucose and insulin responses. Cereal. Chem. 2010, 87, 257–262. [Google Scholar] [CrossRef]
- Segundo, C.; Roman, L.; Gomez, M.; Martinez, M.M. Mechanically fractionated flour isolated from green bananas (M. cavendishii var. nanica) as a tool to increase the dietary fiber and phytochemical bioactivity of layer and sponge cakes. Food Chem. 2017, 219, 240–248. [Google Scholar] [CrossRef]
- Segundo, C.; Roman, L.; Lobo, M.; Martinez, M.M.; Gomez, M. Ripe banana flour as a source of antioxidants in layer and sponge cakes. Plant. Food Hum. Nutr. 2017, 72, 365–371. [Google Scholar] [CrossRef]
- Mao, W.W.; Kinsella, J.E. Amylase activity in banana fruit: Properties and changes in activity with ripening. J. Food Sci. 1981, 46, 1400–1403. [Google Scholar] [CrossRef]
- Pico, J.; Xu, K.; Guo, M.; Mohamedshah, Z.; Ferruzzi, M.G.; Martinez, M.M. Manufacturing the ultimate green banana flour: Impact of drying and extrusion on phenolic profile and starch bioaccessibility. Food Chem. 2019, 297, 124990. [Google Scholar] [CrossRef] [PubMed]
- CODEX, Joint FAO/WHO Food Standards Programme, Secretariat of the CODEX Alimentarius Commission: CODEX Alimentarius (CODEX) Guidelines on Nutrition Labeling CAC/GL 2–1985 as Last Amended 2010. Available online: http://www.fao.org/ag/humannutrition/33309-01d4d1dd1abc825f0582d9e5a2eda4a74.pdf (accessed on 26 June 2019).
- Eerlingen, R.C.; Van Haesendonck, I.P.; De Paepe, G.; Delcour, J.A. Enzyme-resistant starch. III. The quality of straight-dough bread containing varying levels of enzyme-resistant starch. Cereal. Chem. 1994, 71, 165–169. [Google Scholar]
- Niba, L.L. Effect of storage period and temperature on resistant starch and beta-glucan content in cornbread. Food Chem. 2003, 83, 493–498. [Google Scholar] [CrossRef]
- Sullivan, W.R.; Hughes, J.G.; Cockman, R.W.; Small, D.M. The effects of temperature on the crystalline properties and resistant starch during storage of white bread. Food Chem. 2017, 228, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Arp, C.G.; Correa, M.J.; Ferrero, C. High-Amylose Resistant Starch as a Functional Ingredient in Breads: A Technological and Microstructural Approach. Food Bioprocess. Tech. 2018, 11, 2182–2193. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Wronkowska, M.; Soral-Śmietana, M.; Collar, C.; Haros, M. Impact of the addition of resistant starch from modified pea starch on dough and bread performance. Eur. Food Res. Technol. 2010, 231, 499–508. [Google Scholar] [CrossRef]
- Van Hung, P.; Yamamori, M.; Morita, N. Formation of Enzyme-Resistant Starch in Bread as Affected by High-Amylose Wheat Flour Substitutions. Cereal. Chem. J. 2005, 82, 690–694. [Google Scholar] [CrossRef]
- Almeida, E.L.; Chang, Y.K.; Steel, C.J. Dietary fibre sources in bread: Influence on technological quality. LWT 2013, 50, 545–553. [Google Scholar] [CrossRef]
- Sharma, A.; Yadav, B.S.; Ritika. Resistant starch: Physiological roles and food applications. Food Rev. Int. 2008, 24, 193–234. [Google Scholar] [CrossRef]
- Akerberg, A.; Liljeberg, H.; Bjorck, I. Effects of amylose/amylopectin ratio and baking conditions on resistant starch formation and glycaemic indices. J. Cereal. Sci. 1998, 28, 71–80. [Google Scholar] [CrossRef]
- Sarawong, C.; Gutierrez, Z.R.; Berghofer, E.; Schoenlechner, R. Effect of green plantain flour addition to gluten-free bread on functional bread properties and resistant starch content. Int. J. Food Sci. Technol. 2014, 49, 1825–1833. [Google Scholar] [CrossRef]
- Giuberti, G.; Fortunati, P.; Gallo, A. Can different types of resistant starch influence the in vitro starch digestion of gluten free breads? J. Cereal Sci. 2016, 100, 253–255. [Google Scholar] [CrossRef]
- Korus, J.; Witczak, M.; Ziobro, R.; Juszczak, L. The impact of resistant starch on characteristics of gluten-free dough and bread. Food Hydrocoll. 2009, 23, 988–995. [Google Scholar] [CrossRef]
- Ozturk, S.; Koksel, H.; Ng, P.K. Farinograph properties and bread quality of flours supplemented with resistant starch. Int. J. Food Sci. Nutr. 2009, 60, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Goesaert, H.; Leman, P.; Delcour, J.A. Model approach to starch functionality in bread making. J. Agric. Food Chem. 2008, 56, 6423–6431. [Google Scholar] [CrossRef] [PubMed]
- Tsatsaragkou, K.; Gounaropoulos, G.; Mandala, I. Development of gluten free bread containing carob flour and resistant starch. LWT 2014, 58, 124–129. [Google Scholar] [CrossRef]
- Barros, J.H.; Telis, V.R.; Taboga, S.; Franco, C.M. Resistant starch: Effect on rheology, quality, and staling rate of white wheat bread. J. Food Sci. Technol. 2018, 55, 4578–4588. [Google Scholar] [CrossRef] [PubMed]
- Collar, C.; Balestra, F.; Ancarani, D. Value added of resistant starch maize-based matrices in breadmaking: Nutritional and functional assessment. Food Bioprocess. Tech. 2014, 7, 3579–3590. [Google Scholar] [CrossRef]
- FDA 2018. U.S. Department of Health and Human Services, Food and Drug Administration, Center for Food Safety and Applied Nutrition. ‘Review of the scientific evidence on the physiological effects of non-digestible carbohydrates.’. June 2018. Available online: https://www.fda.gov/downloads/Food/LabelingNutrition/UCM610139.pdf. (accessed on 23 February 2019).
- European Commission 2011. Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to consumers, amending Regulations (EC) No 1924/2006 and (EC) No 1925/2006 of the European Parliament and of the Council, and repealing Commission Directive 87/250/EEC, Council Directive 90/496/EEC, Commission Directive 1999/10/EC, Directive 2000/13/EC of the European Parliament and of the Council, Commission Directives 2002/67/EC and 2008/5/EC and Commission Regulation (EC) No 608/2004. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32011R1169&from=EN. (accessed on 23 February 2019).
- Nugent, A.P. Health properties of resistant starch. Nutr. Bull. 2005, 30, 27–54. [Google Scholar] [CrossRef]
- European Commission 2006. Regulation (EC) No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on nutrition and health claims made on foods. Official Journal of the European Union, L404, 9–25. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:404:0009:0025:EN:PDF (accessed on 23 February 2019).
- Stephen, A.M.; Champ, M.M.J.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary fibre in Europe: Current state of knowledge on definitions, sources, recommendations, intakes and relationships to health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef] [PubMed]



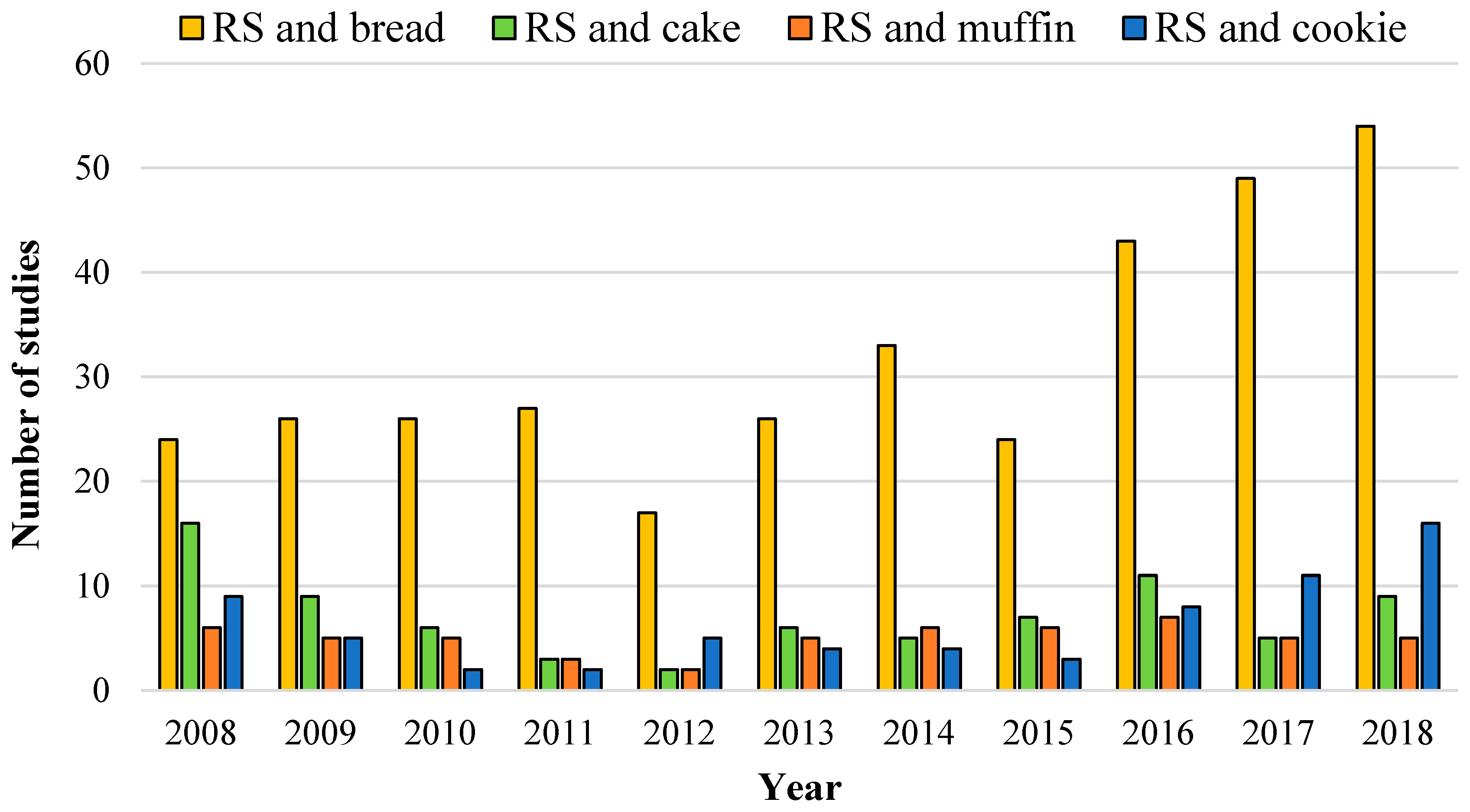{kind=link}
{kind=link}
| Classification | Structural Features Conferring the RS Property within Each Category | Detrimental Steps That May Decrease RS Content during Bread-Making | Assisting Steps That May Increase RS Content during Bread-Making |
|---|---|---|---|
| RS1 | Intact plant tissues | Milling, sieving, baking | - |
| Highly dense food matrices | - | Baking and cooling | |
| Confined starch within a continuous layer of certain proteins | - | Baking of starch materials containing specific layer forming proteins | |
| RS2 | Starch granules with an outer high-density shell structure | Baking (of note that high amylose RS2 is more heat-resistant) | - |
| RS3 | Retrograded amylose | - | Baking and cooling |
| High-density processed amylose | - | Extrusion of high amylose starch ingredients | |
| Retrograded amylopectin | Baking | Baking and cooling | |
| RS4 | Chemically substituted starches | - | - |
| Chemically cross-linked starches | - | - | |
| a Resistant maltodextrins | - | - | |
| RS5 | Amorphous amylose-lipid complexes (form I) | - | Baking and cooling |
| Crystalline amylose-lipid complexes (form II) | - | Baking and cooling |
| Ingredient | RS Content | Type of Bread | Substitution Level (%) | Evaluation Day | RS Content | In vitro RS Method | Effects on Bread Quality | Refs. |
|---|---|---|---|---|---|---|---|---|
| HA maize starch, Hi-Maize 260, Ingredion | 60% IDF Manufacturer (58.4% TDF) | Wheat flour (4.5% TDF) | 0 10 20 30 | n.a. crumb | 6.6% bb, db 9.5% bb, db 17.0% bb, db 26.6% bb, db | AOAC 985.29 | Increased hardness Decreased cohesiveness and resilience Decreased volume with 30% level Lighter crust Decreased number of cells in the crumb Decreased Cinf and estimated GI Increased consumer acceptability (20% level) | [130] |
| HA maize starch, Hi-Maize 260, Ingredion | 60% RS Manufacturer | Wheat flour | 0 10 15 20 | 2 h, Lyophilized crumb | 1.2% n.a. 3.9% n.a. 5.9% n.a. 11.1% n.a. | Goni et al. [40] | Decreased volume Increased hardness with 15% level Lighter crust Staling dependent on the level of replacement | [142] |
| HA, Amylo-maize starch N-400, Roquette | 40% TDF Manufacturer | GF flour mix (maize starch, rice flour, tapioca starch) | 0 20—RS2 20—RS3 a | n.a. bread | 1.2% bb, db 4.4% bb, db 7.6–9.2% bb, db | Englyst et al. [31] | Reduced in vitro glycemic index (RS3 > RS2) | [137] |
| HA maize starch, Hi-Maize 260, Ingredion | 56% TDF Manufacturer | Yellow maize flour | 0 20 | n.a. | 4.3% bb, db 12.0% bb, db | Modified AOAC 2002.02 | Specific volume and texture were not modified Decreased cell density Similar sensory evaluation SDS fraction was also increased eGI decreased from 85 to 71 | [143] |
| HA maize starch, Hi-Maize 260, Ingredion | 56% TDF Manufacturer | White maize flour | 0 20 | n.a. | 5.5% bb, db 11.3% bb, db | Modified AOAC 2002.02 | Specific volume and texture were not modified Same cell density Similar sensory evaluation SDS fraction was also increased eGI decreased from 83 to 72 | [143] |
| HA maize starch, Eurylon, Roquette | 83.2% RS2 | Wheat flour (14.1% RS) | 0 20 b | 24 h/7 days, Lyophilized crumb | 0.0%/4.4% bb, db 7.7%/10.2% bb, db | Modified Englyst et al. [31] | Decreased specific volume Decreased hardness No sensory differences | [127] |
| Extruded retrograded HA maize starch, EURESTA, Cerestar | 29.5% RS3 | Wheat flour (14.1% RS) | 0 20 b | 24 h/7 days, Lyophilized crumb | 0.0%/4.4% bb, db 8.4%/11.0% bb, db | Modified Englyst et al. [31] | Decreased specific volume Decreased hardness No sensory differences | [127] |
| HA maize starch, HylonVII, Ingredion | 53% RS2 | Wheat flour | 0 10 20 30 | 24 h, Lyophilized crumb | 1.2% bb, db 4.1% bb, db 8.1% bb, db 10.1% bb, db | AOAC 2002.02 | Decreased volume for 30% level Increased hardness for 30% level Paler crust color for 20% and 30% levels | [139] |
| HA maize starch, Novelose330, Ingredion | 46.5% RS3 | Wheat flour | 0 10 20 30 | 24 h, Lyophilized crumb | 1.2% bb, db 4.7% bb, db 9.7% bb, db 12.7% bb, db | AOAC 2002.02 | Decreased volume 20% and 30% levels Increased hardness for 20% and 30% levels Paler crust color for 30% | [139] |
| HA maize starch CrystaLean, SunOpta ingredients | 45% RS3 | Wheat flour | 0 10 20 30 | 24 h, Lyophilized crumb | 1.2% bb, db 4.4% bb, db 8.3% bb, db 12.4% bb, db | AOAC 2002.02 | Decreased volume for 30% level Increased hardness for 30% Paler crust color above 30% | [139] |
| HA maize starch, Hi-Maize 260, Ingredion | RS > 60% Manufacturer | Maize starch Potato starch (4:1) c | 0 10 15 20 | n.a. | 2.1% bb 3.9% bb 4.7% bb 5.0% bb | AOAC 991.43 | Decreased volume Similar initial hardness (20% level) Reduced hardening (48 h) | [138] |
| Tapioca starch, ActiStar 11700, Cargill | RS3 > 50% Manufacturer | Maize starch: Potato starch (4:1 mixture) d | 0 10 15 20 | n.a. | 2.1% bb 2.5% bb 2.8% bb 3.0% bb | AOAC 991.43 | Decreased volume Reduced initial hardness (20% level) Reduced hardening (48 h) | [138] |
| HA wheat flour, Okumoto Flour milling | 6.7% TDF | Wheat flour (3.4% TDF) | 0 10 30 50 | 2h | 0.9% bb, db 1.6% bb, db 2.6% bb, db 3.0% bb, db | AOAC 985.29 e | Decreased volume Increased hardness Similar staling (hardness) for 30 and 50% levels Higher staling for 10% level Increasing RS with storage and substitution | [132] |
| Green banana starch, Natural Evolution | 42.2% RS2 | Maize starch (0.8% RS): Rice flour (0.1% RS) (1:1 mixture) | 0 20—Native 20—Extruded f | 24 h, Crumb and crust | 1.5% cb – 0.3% ct, db 1.7% cb –5.7% ct, db 1.9% cb –0.7% ct, db | AOAC 2002.02 | Darker bread color Improved volume, reduced hardness, and improved sensory acceptance (native banana) Increased SDS fraction in crumb with native and extruded banana | [9] |
| Green plantain flour, Chiquita | 50.1% RS2 | Rice flour: GF wheat starch (1:1 mixture) | 0 35 | 24 h | 1.1% bb, db 2.3% bb, db | AOAC 2002.02 | Improved volume but increased firmness Darker bread crumb Lighter bread crust Optimization of water content, baking time and temperature for 30% replacement to maximize RS (3%) | [136] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roman, L.; Martinez, M.M. Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives. Foods 2019, 8, 267. https://doi.org/10.3390/foods8070267
Roman L, Martinez MM. Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives. Foods. 2019; 8(7):267. https://doi.org/10.3390/foods8070267
Chicago/Turabian StyleRoman, Laura, and Mario M. Martinez. 2019. "Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives" Foods 8, no. 7: 267. https://doi.org/10.3390/foods8070267
APA StyleRoman, L., & Martinez, M. M. (2019). Structural Basis of Resistant Starch (RS) in Bread: Natural and Commercial Alternatives. Foods, 8(7), 267. https://doi.org/10.3390/foods8070267