Pistacia terebinthus Resin as Yeast Immobilization Support for Alcoholic Fermentation


 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strain
2.2. Yeast Cells Immobilization within Pistacia Terebinthus Resin
2.3. Monitor of Yeast Cells’ Immobilization
2.4. Successive Batch Fermentations of Various Sugars at Different Temperatures
2.5. Ethanol, Residual Sugar, and Major Volatiles Analyses
2.6. Phenolic Content Determination
2.7. Head space (HS) Solid Phase Microextraction (SPME) Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.8. Preliminary Sensory Evaluation
2.9. Experimental Design and Statistical Analysis
3. Results and Discussion
3.1. Yeast Cells Immobilization
3.2. Successive Fermentations Batches by Various Sugars at Different Temperatures
3.3. Formation of Major Volatile By-Products
3.4. SPME GC-MS Analysis
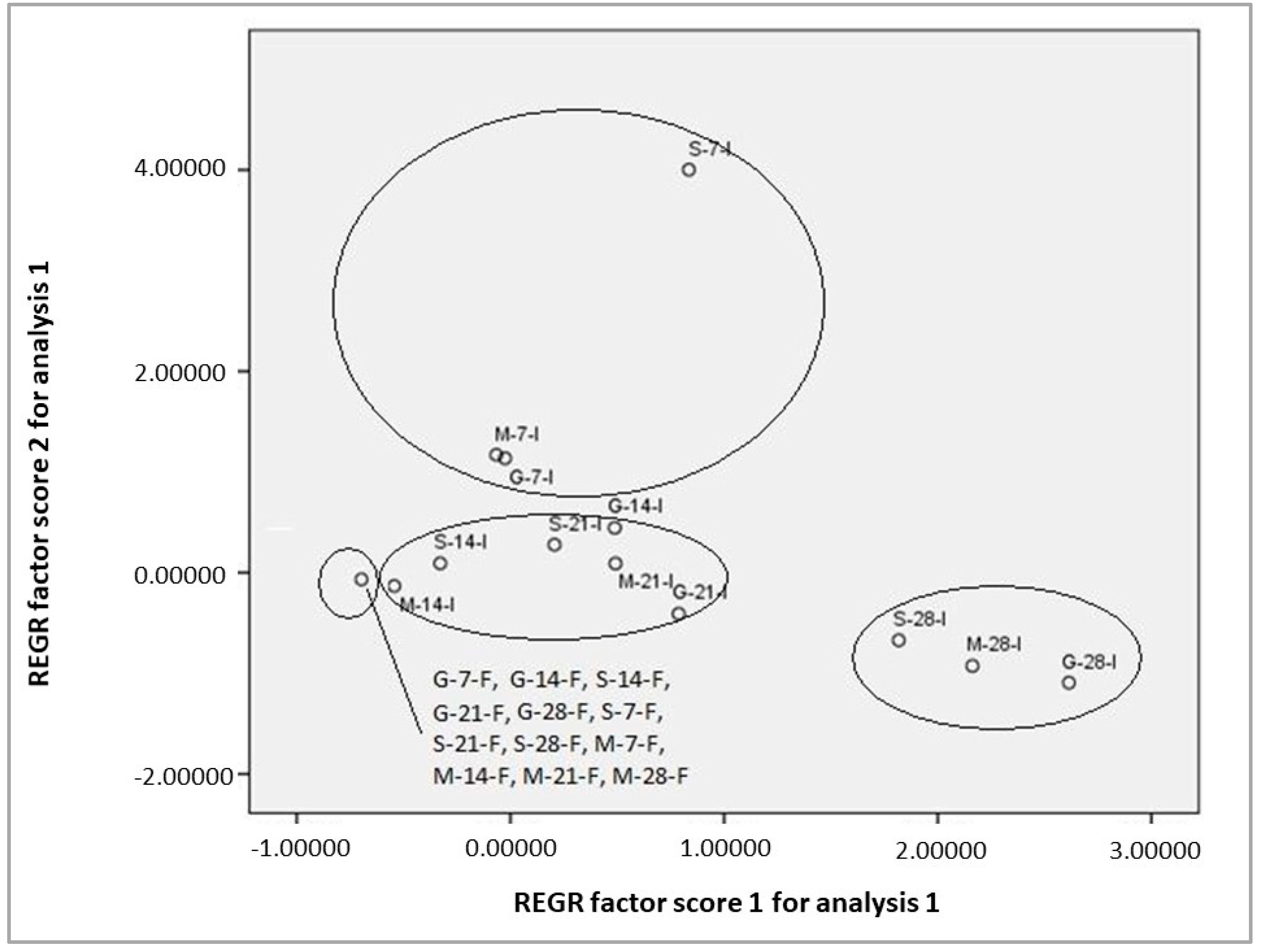
3.5. Chemometrics
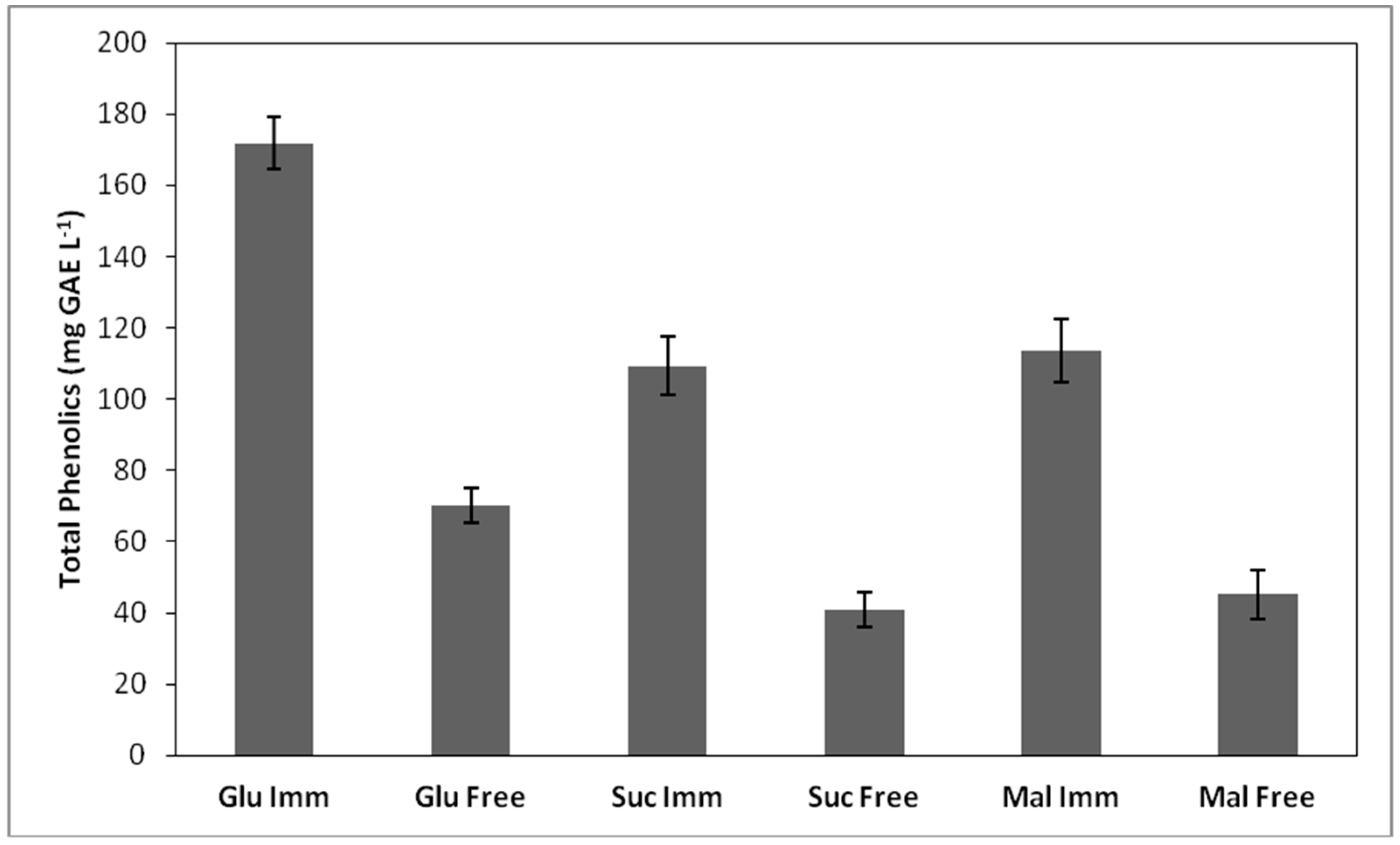
3.6. Phenolic Content
3.7. Preliminary Sensory Evaluation and Resistance to Spoilage
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Orhan, I.E.; Senol, F.S.; Gulpinar, A.R.; Sekeroglu, N.; Kartal, M.; Sener, B. Neuroprotective potential of some terebinth coffee brands and the unprocessed fruits of Pistacia terebinthus L. and their fatty and essential oil analyses. Food Chem. 2012, 130, 882–888. [Google Scholar] [CrossRef]
- Schoina, V.; Terpou, A.; Bosnea, L.; Kanellaki, M.; Nigam, P.S. Entrapment of Lactobacillus casei ATCC393 in the viscus matrix of Pistacia terebinthus resin for functional myzithra cheese manufacture. LWT 2018, 89, 441–448. [Google Scholar] [CrossRef]
- Rauf, A.; Patel, S.; Uddin, G.; Siddiqui, B.S.; Ahmad, B.; Muhammad, N.; Mabkhot, Y.N.; Hadda, T.B. Phytochemical, ethnomedicinal uses and pharmacological profile of genus Pistacia. Biomed. Pharmacother. 2017, 86, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Kavak, D.D.; Altıok, E.; Bayraktar, O.; Ülkü, S. Pistacia terebinthus extract: As a potential antioxidant, antimicrobial and possible β-glucuronidase inhibitor. J. Mol. Catal. B Enzym. 2010, 64, 167–171. [Google Scholar] [CrossRef]
- Sargin, S.A.; Büyükcengiz, M. Plants used in ethnomedicinal practices in Gulnar district of Mersin, Turkey. J. Herb. Med. 2018. [Google Scholar] [CrossRef]
- Buriani, A.; Fortinguerra, S.; Sorrenti, V.; Dall’Acqua, S.; Innocenti, G.; Montopoli, M.; Gabbia, D.; Carrara, M. Human Adenocarcinoma Cell Line Sensitivity to Essential Oil Phytocomplexes from Pistacia Species: A Multivariate Approach. Molecules 2017, 22, 1336. [Google Scholar] [CrossRef] [PubMed]
- Duru, M.E.; Cakir, A.; Kordali, S.; Zengin, H.; Harmandar, M.; Izumi, S.; Hirata, T. Chemical composition and antifungal properties of essential oils of three Pistacia species. Fitoterapia 2003, 74, 170–176. [Google Scholar] [CrossRef]
- Giner-Larza, E.M.; Máñez, S.; Recio, M.C.; Giner, R.M.; Prieto, J.M.; Cerdá-Nicolás, M.; Ríos, J.L. Oleanonic acid, a 3-oxotriterpene from Pistacia, inhibits leukotriene synthesis and has anti-inflammatory activity. Eur. J. Pharmacol. 2001, 428, 137–143. [Google Scholar] [CrossRef]
- Lamiri, A.; Lhaloui, S.; Benjilali, B.; Berrada, M. Insecticidal effects of essential oils against Hessian fly, Mayetiola destructor (Say). Field Crop. Res. 2001, 71, 9–15. [Google Scholar] [CrossRef]
- Mouhajir, F.; Hudson, J.B.; Rejdali, M.; Towers, G.H.N. Multiple antiviral activities of endemic medicinal plants used by Berber peoples of Morocco. Pharm. Biol. 2001, 39, 364–374. [Google Scholar] [CrossRef]
- Tuzlaci, E.; Aymaz, P.E. Turkish folk medicinal plants, Part IV: Gonen (Balikesir). Fitoterapia 2001, 72, 323–343. [Google Scholar] [CrossRef]
- Mohagheghzadeh, A.; Faridi, P.; Ghasemi, Y. Analysis of Mount Atlas mastic smoke: A potential food preservative. Fitoterapia 2010, 81, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Schoina, V.; Terpou, A.; Angelika-Ioanna, G.; Koutinas, A.; Kanellaki, M.; Bosnea, L. Use of Pistacia terebinthus resin as immobilization support for Lactobacillus casei cells and application in selected dairy products. J. Food Sci. Technol. 2015, 52, 5700–5708. [Google Scholar] [CrossRef] [PubMed]
- Genisheva, Z.; Teixeira, J.A.; Oliveira, J.M. Immobilized cell systems for batch and continuous winemaking. Trends Food Sci. Technol. 2014, 40, 33–47. [Google Scholar] [CrossRef]
- Kourkoutas, Y.; Bekatorou, A.; Banat, I.M.; Marchant, R.; Koutinas, A.A. Immobilization technologies and support materials suitable in alcohol beverages production: A review. Food Microbiol. 2004, 21, 377–397. [Google Scholar] [CrossRef]
- Ganatsios, V.; Koutinas, A.A.; Bekatorou, A.; Kanellaki, M.; Nigam, P. Promotion of maltose fermentation at extremely low temperatures using a cryotolerant Saccharomyces cerevisiae strain immobilized on porous cellulosic material. Enzym. Microb. Technol. 2014, 66, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Armani, M.; Morozova, K.; Scampicchio, M. Immobilization of Saccharomyces cerevisiae on nylon-6 nanofibrous membranes for grape juice fermentation. LWT 2018. [Google Scholar] [CrossRef]
- Ejaz, U.; Ahmed, A.; Sohail, M. Statistical optimization of immobilization of yeast cells on corncob for pectinase production. Biocatal. Agric. Biotechnol. 2018, 14, 450–456. [Google Scholar] [CrossRef]
- Gialleli, A.I.; Kallis, M.; Bekatorou, A.; Kanellaki, M.; Koutinas, A.A. Continuous Cold Pasteurisation of Contaminated Wine Using Nano- and Micro-Tubular Cellulose. Food Bioprocess Technol. 2014, 8, 539–547. [Google Scholar] [CrossRef]
- Terpou, A.; Nigam, P.S.; Bosnea, L.; Kanellaki, M. Evaluation of Chios mastic gum as antimicrobial agent and matrix forming material targeting probiotic cell encapsulation for functional fermented milk production. LWT 2018. [Google Scholar] [CrossRef]
- de Souza Júnior, W.C.; de Rezende, S.T.; Viana, P.A.; Falkoski, D.L.; Reis, A.P.; Machado, S.G.; Barros, E.G.D.; Guimarães, V.M. Treatment of soy milk with Debaryomyces hansenii cells immobilised in alginate. Food Chem. 2009, 114, 589–593. [Google Scholar] [CrossRef]
- Bosnea, L.A.; Kopsahelis, N.; Kokkali, V.; Terpou, A.; Kanellaki, M. Production of a novel probiotic yogurt by incorporation of L. casei enriched fresh apple pieces, dried raisins and wheat grains. Food Bioprod. Process. 2017, 102, 62–71. [Google Scholar] [CrossRef]
- Nikolaou, A.; Tsakiris, A.; Kanellaki, M.; Bezirtzoglou, E.; Akrida-Demertzi, K.; Kourkoutas, Y. Wine production using free and immobilized kefir culture on natural supports. Food Chem. 2019, 272, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Mallouchos, A.; Reppa, P.; Aggelis, G.; Kanellaki, M.; Koutinas, A.A.; Komaitis, M. Grape skins as a natural support for yeast immobilization. Biotechnol. Lett. 2002, 24, 1331–1335. [Google Scholar] [CrossRef]
- Genisheva, Z.; Vilanova, M.; Mussatto, S.I.; Teixeira, J.A.; Oliveira, J.M. Consecutive alcoholic fermentations of white grape musts with yeasts immobilized on grape skins—Effect of biocatalyst storage and SO2 concentration on wine characteristics. LWT Food Sci. Technol. 2014, 59, 1114–1122. [Google Scholar] [CrossRef]
- Kopsahelis, N.; Kanellaki, M.; Bekatorou, A. Low temperature brewing using cells immobilized on brewer’s spent grains. Food Chem. 2007, 104, 480–488. [Google Scholar] [CrossRef]
- Walker, G.M.; Walker, R.S.K. Enhancing Yeast Alcoholic Fermentations. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Argiriou, T.; Kalliafas, A.; Psarianos, C.; Kana, K.; Kanellaki, M.; Koutinas, A.A. New alcohol resistant strains of Saccharomyces cerevisiae species for potable alcohol production using molasse. Appl. Biochem. Biotechnol. 1992, 36, 153–161. [Google Scholar] [CrossRef]
- Kopsahelis, N.; Nisiotou, A.; Kourkoutas, Y.; Panas, P.; Nychas, G.J.E.; Kanellaki, M. Molecular characterization and molasses fermentation performance of a wild yeast strain operating in an extremely wide temperature range. Bioresour. Technol. 2009, 100, 4854–4862. [Google Scholar] [CrossRef]
- Ganatsios, V.; Koutinas, A.A.; Bekatorou, A.; Panagopoulos, V.; Banat, I.M.; Terpou, A.; Kopsahelis, N. Porous cellulose as promoter of oil production by the oleaginous yeast Lipomyces starkeyi using mixed agroindustrial wastes. Bioresour. Technol. 2017, 244, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Bardi, E.P.; Koutinas, A.A. Immobilization of Yeast on Delignified Cellulosic Material for Room Temperature and Low-Temperature Wine Making. J. Agric. Food Chem. 1994, 42, 221–226. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total Phenol Analysis: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Tsakiris, A.; Kourkoutas, Y.; Dourtoglou, V.G.; Koutinas, A.A.; Psarianos, C.; Kanellaki, M. Wine produced by immobilized cells on dried raisin berries in sensory evaluation comparison with commercial products. J. Sci. Food Agric. 2006, 86, 539–543. [Google Scholar] [CrossRef]
- Iconomopoulou, M.; Psarianos, K.; Kanellaki, M.; Koutinas, A.A. Low temperature and ambient temperature wine making using freeze dried immobilized cells on gluten pellets. Process Biochem. 2002, 37, 707–717. [Google Scholar] [CrossRef]
- Bardi, E.P.; Bakoyianis, V.; Koutinas, A.A.; Kanellaki, M. Room temperature and low temperature wine making using yeast immobilized on gluten pellets. Process Biochem. 1996, 31, 425–430. [Google Scholar] [CrossRef]
- Argiriou, T.; Kaliafas, A.; Psarianos, K.; Kanellaki, M.; Voliotis, S.; Koutinas, A.A. Psychrotolerant Saccharomyces cerevisiae strains after an adaptation treatment for low temperature wine making. Process Biochem. 1996, 31, 639–643. [Google Scholar] [CrossRef]
- Kourkoutas, Y.; Douma, M.; Koutinas, A.A.; Kanellaki, M.; Banat, I.M.; Marchant, R. Continuous winemaking fermentation using quince-immobilized yeast at room and low temperatures. Process Biochem. 2003, 39, 143–148. [Google Scholar] [CrossRef]
- Mallouchos, A.; Komaitis, M.; Koutinas, A.; Kanellaki, M. Wine fermentations by immobilized and free cells at different temperatures. Effect of immobilization and temperature on volatile by-products. Food Chem. 2003, 80, 109–113. [Google Scholar] [CrossRef]
- Terpou, A.; Bosnea, L.; Kanellaki, M. Effect of Mastic Gum (Pistacia Lentiscus via Chia) as a Probiotic Cell Encapsulation Carrier for Functional Whey Beverage Production. SCIOL Biomed. 2017, 1, 1–10. [Google Scholar]
- Regodón Mateos, J.A.; Pérez-Nevado, F.; Ramírez Fernández, M. Influence of Saccharomyces cerevisiae yeast strain on the major volatile compounds of wine. Enzym. Microb. Technol. 2006, 40, 151–157. [Google Scholar] [CrossRef]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.-P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef]
- Antonelli, A.; Castellari, L.; Zambonelli, C.; Carnacini, A. Yeast Influence on Volatile Composition of Wines. J. Agric. Food Chem. 1999, 47, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Kopsahelis, N.; Bosnea, L.; Kanellaki, M.; Koutinas, A.A. Volatiles Formation from Grape Must Fermentation Using a Cryophilic and Thermotolerant Yeast. Appl. Biochem. Biotechnol. 2012, 167, 1183–1198. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Minnaar, P.; Nyobo, L.; Jolly, N.; Ntushelo, N.; Meiring, S. Anthocyanins and polyphenols in Cabernet Franc wines produced with Saccharomyces cerevisiae and Torulaspora delbrueckii yeast strains: Spectrophotometric analysis and effect on selected sensory attributes. Food Chem. 2018, 268, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Gironi, F.; Piemonte, V. Temperature and solvent effects on polyphenol extraction process from chestnut tree wood. Chem. Eng. Res. Des. 2011, 89, 857–862. [Google Scholar] [CrossRef]
- Bouyahya, A.; Chadon Assemian, I.C.; Mouzount, H.; Bourais, I.; Et-Touys, A.; Fellah, H.; Benjouad, A.; Dakka, N.; Bakri, Y. Could volatile compounds from leaves and fruits of Pistacia lentiscus constitute a novel source of anticancer, antioxidant, antiparasitic and antibacterial drugs? Ind. Crop. Prod. 2019, 128, 62–69. [Google Scholar] [CrossRef]
- Garaguso, I.; Nardini, M. Polyphenols content, phenolics profile and antioxidant activity of organic red wines produced without sulfur dioxide/sulfites addition in comparison to conventional red wines. Food Chem. 2015, 179, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Bąkowska, A.; Kucharska, A.Z.; Oszmiański, J. The effects of heating, UV irradiation, and storage on stability of the anthocyanin–polyphenol copigment complex. Food Chem. 2003, 81, 349–355. [Google Scholar] [CrossRef]
- Papageorgiou, V.P.; Assimopoulou, A.N.; Yannovits-Argiriadis, N. Chemical composition of the essential oil of chios turpentine. J. Essent. Oil Res. 1999, 11, 367–368. [Google Scholar] [CrossRef]
- Monaco, P.; Previtera, L.; Mangoni, L. Terpenes in Pistacia-plants—A possible defense role for monoterpenes against gall-forming aphids. Phytochemistry 1982, 21, 2408–2410. [Google Scholar] [CrossRef]
- Kotan, R.; Kordali, S.; Cakir, A. Screening of antibacterial activities of twenty-one oxygenated monoterpenes. Z. Nat. Sect. C J. Biosci. 2007, 62, 507–513. [Google Scholar] [CrossRef]
- Jabra-Rizk, M.A.; Meiller, T.F.; James, C.E.; Shirtliff, M.E. Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob. Agents Chemother. 2006, 50, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Leite, B.; Teixeira, P.; Cerca, N.; Azeredo, J.; Oliveira, R. Farnesol as antibiotics adjuvant in staphylococcus epidermidis control in vitro. Am. J. Med. Sci. 2011, 341, 191–195. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Batch No. | T (°C) | Fermentation Time (h) | Ethanol (g L−1) | Ethanol Productivity (g L−1 Day−1) | Residual Sugar (g L−1) | Sugar Conversion (%) | |
|---|---|---|---|---|---|---|---|
| Free cells | Glucose | ||||||
| 1−8 | 28 | 29.0 ± 3.2 | 43.45 ± 5.5 | 35.9 ± 1.7 | 3.4 ± 1.2 | 97.5 ± 2.5 | |
| 9−13 | 21 | 42.0 ± 3.7 | 40.29 ± 5.5 | 19.9 ± 2.3 | 3.3 ± 1.5 | 98.2 ± 1.7 | |
| 14−16 | 14 | 133.3 ± 6.1 | 36.34 ± 3.9 | 9.0 ± 0.3 | 5.9 ± 0.7 | 95.3 ± 0.3 | |
| 17−18 | 7 | 1140 ± 84.9 | 32.39 ± 2.4 | 0.7 ± 0.1 | 22.3 ± 2.1 | 83.1 ± 0.2 | |
| Sucrose | |||||||
| 1−8 | 28 | 32.1 ± 2.9 | 46.60 ± 5.5 | 34.8 ± 5.6 | 19.9 ± 6.4 | 85.0 ± 4.9 | |
| 9−13 | 21 | 60.2 ± 2.6 | 40.30 ± 5.5 | 16.2 ± 2.4 | 20.5 ± 1.9 | 84.5 ± 1.5 | |
| 14−16 | 14 | 146 ± 16.7 | 36.34 ± 3.9 | 6.0 ± 1.3 | 20.9 ± 4.6 | 84.2 ± 3.5 | |
| 17−18 | 7 | 1520 ± 84.9 | 41.08 ± 2.4 | 0.7 ± 0.1 | 22.4 ± 3.3 | 83.0 ± 2.5 | |
| Maltose | |||||||
| 1−8 | 28 | 26.1 ± 1.4 | 46.61 ± 5.5 | 42.5 ± 4.8 | 3.1 ± 1.0 | 97.6 ± 4.0 | |
| 9−13 | 21 | 50.8 ± 3.1 | 32.40 ± 3.2 | 15.3 ± 0.9 | 5.4 ± 3.5 | 96.1 ± 2.8 | |
| 14−16 | 14 | 167.3 ± 14.6 | 32.39 ± 2.4 | 4.7 ± 0.7 | 13.3 ± 3.1 | 90.1 ± 2.0 | |
| 17−18 | 7 | 2160 ± 99.0 | 41.10 ± 12.6 | 0.5 ± 0.1 | 16.1 ± 1.3 | 88.3 ± 0.4 | |
| Immobilized cells | Glucose | ||||||
| 1−8 | 28 | 19.6 ± 2.7 | 37.30 ± 2.4 | 45.8 ± 4.7 | 9.4 ± 6.5 | 93.0 ± 5.4 | |
| 9−13 | 21 | 23.9 ± 3.5 | 37.30 ± 1.6 | 37.5 ± 5.7 | 10.1 ± 2.3 | 92.7 ± 7.1 | |
| 14−16 | 14 | 29.9 ± 1.9 | 45.03 ± 2.4 | 36.4 ± 1.9 | 9.1 ± 3.4 | 93.1 ± 4.2 | |
| 17−18 | 7 | 376 ± 24.0 | 39.50 ± 0.8 | 2.6 ± 0.2 | 20.3 ± 3.3 | 84.7 ± 2.3 | |
| Sucrose | |||||||
| 1−8 | 28 | 24.2 ± 4.3 | 42.66 ± 6.3 | 43.4 ± 8.0 | 20.6 ± 11.8 | 84.4 ± 8.9 | |
| 9−13 | 21 | 33.3 ± 4.6 | 42.66 ± 3.9 | 30.9 ± 4.6 | 17.2 ± 6.3 | 86.9 ± 4.8 | |
| 14−16 | 14 | 53.5 ± 1.0 | 44.24 ± 4.7 | 19.9 ± 1.8 | 13.9 ± 10.5 | 89.5 ± 9.0 | |
| 17−18 | 7 | 705 ± 59.8 | 50.56 ± 2.4 | 1.7 ± 0.2 | 13.6 ± 1.4 | 89.5 ± 0.9 | |
| Maltose | |||||||
| 1−8 | 28 | 24.0 ± 3.9 | 39.50 ± 4.7 | 39.7 ± 7.6 | 21.1 ± 6.2 | 84.1 ± 4.8 | |
| 9−13 | 21 | 22.7 ± 4.5 | 41.90 ± 3.9 | 45.6 ± 5.6 | 15.4 ± 5.8 | 89.1 ± 4.9 | |
| 14−16 | 14 | 34.7 ± 3.3 | 37.13 ± 5.5 | 25.8 ± 6.0 | 5.3 ± 2.4 | 96.4 ± 2.4 | |
| 17−18 | 7 | 792 ± 30.2 | 34.76 ± 1.6 | 1.0 ± 0.1 | 21.3 ± 4.3 | 83.8 ± 2.7 | |
| Substrate | Batch No. | T (°C) | MeOH (mg L−1) | Acetaldehyde (mg L−1) | Ethyl Acetate (mg L−1) | Propanol (mg L−1) | Isobutyl Alcohol (mg L−1) | Amyl Alcohols (mg L−1) | Total Polyphenols (mg GAE L−1) |
|---|---|---|---|---|---|---|---|---|---|
| Glucose | Free cells | ||||||||
| 1−8 | 28 | 16.3 ± 6.0 e,a,b,f,g | 49.8 ± 14.6 e,a,d | 9.9 ± 1.7 e,a,d,f,h | Tr a | 2.9 ± 0.3 e | 15.8 ± 2.6 e,a,b,d,g,h | 48.5 ± 4.3 e,g | |
| 9−13 | 21 | 12.9 ± 3.6 f,a,b,e | 32.0 ± 11.2 f,a,b,c,d,g,h | 8.6 ± 1.9 f,a,b,c,d,e,h | 0.5 ± 0.2 c | 1.0 ± 0.3 f,b,c,d | 5.3 ± 1.5 f,c | 70.2 ± 4.7 f,d | |
| 14−16 | 14 | 24.1 ± 7.3 g,a,b,c,d,e | 18.4 ± 3.7 g,b,c,f,h | 5.6 ± 1.1 g,a,b,c | 1.3 ± 0.6 b | 4.3 ± 0.7 g,a,h | 13.8 ± 2.6 g,b,c,d,e,h | 47.1 ± 4.7 g,e | |
| 17−18 | 7 | 46.9 ± 10.6 h | 23.0 ± 3.2 h,a,b,c,f,g | 10.3 ± 3.5 h,a,d,e,f | Tr a | 4.8 ± 1.2 h,a,g | 18.2 ± 3.9 h,a,d,e,g | 24.3 ± 4.7 h | |
| Immobilized cells | |||||||||
| 1−8 | 28 | 21.1 ± 5.8 a,b,c,e,f,g | 37.0 ± 10.1 a,b,c,d,e,f,h | 7.5 ± 1.1 a,b,c,d,e,f,g,h | Tr a | 4.2 ± 0.8 a,g,h | 19.7 ± 4.1 a,d,e,h | 169.3 ± 6.8 a,b | |
| 9−13 | 21 | 21.7 ± 5.3 b,a,c,e,f,g | 20.8 ± 2.2 b,a,c,f,g,h | 6.9 ± 1.2 b,a,c,f,g | 1.0 ± 0.4 b | 1.7 ± 0.9 b,d,f | 10.7 ± 2.2 b,c,d,e,g | 171.8 ± 7.3 b,a | |
| 14−16 | 14 | 28.5 ± 6.7 c,a,b,d,g | 23.5 ± 4.4 c,a,b,d,f,g,h | 6.3 ± 1.4 c,a,b,f,g | Tr a | Tr c,d,f | 9.8 ± 3.1 c,b,f,g | 104.3 ± 5.5 c | |
| 17−18 | 7 | 31.9 ± 7.1 d,c,g | 40.6 ± 7.3 d,a,c,e,f | 9.9 ± 2.9 d,a,e,f,h | Tr a | 0.9 ± 0.3 d,b,c,f | 15.5 ± 3.7 d,a,b,e,g,h | 74.3 ± 4.3 d,f | |
| Sucrose | Free cells | ||||||||
| 1−8 | 28 | 33.5 ± 7.5 e,a,f,g | 38.9 ± 7.1 e,d,f,h | 3.2 ± 1.1 e,a,c,d,g,h | Tr d | 5.0 ± 1.4 e,a | 15.5 ± 4.9 a | 40.2 ± 5.5 e,f,g,h | |
| 9−13 | 21 | 36.0 ± 8.8 f,a,e,g | 30.7 ± 7.3 f,a,b,d,e,g | 9.7 ± 4.5 f,b | Tr d | Tr f,c,d,g,h | Tr b | 41.0 ± 4.9 f,e,g,h | |
| 14−16 | 14 | 39.7 ± 7.4 g,d,e,f,h | 25.1 ± 6.7 g,a,b,c,f | Tr g,e | Tr d | Tr g,c,d,f,h | Tr b | 49.3 ± 5.4 g,e,f | |
| 17−18 | 7 | 50.4 ± 11.4 h,d,g | 44.3 ± 11.3 h,d,e | 4.5 ± 1.2 h,a,c,d,e | Tr d | 1.2 ± 0.6 h,b,c,d,f,g | Tr b | 34.3 ± 4.7 h,e,f | |
| Immobilized cells | |||||||||
| 1−8 | 28 | 24.7 ± 5.1 a,f | 24.0 ± 5.3 a,b,c,f,g | 5.0 ± 1.7 a,c,d,e,h | 1.6 ± 0.6 a,b,c | 4.2 ± 1.4 a,b,e | 13.3 ± 2.2 a | 101.8 ± 7.1 a,b | |
| 9−13 | 21 | 21.3 ± 3.7 b,a,c | 21.3 ± 2.9 b,a,c,f,g | 8.9 ± 1.4 b,c,f | 1.3 ± 0.5 b,a | 2.7 ± 1.1 b,a,c,h | 15.1 ± 3.3 a | 109.3 ± 8.2 b,a | |
| 14−16 | 14 | 19.2 ± 3.5 c,a | 15.2 ± 6.0 c,a,b,g | 5.9 ± 1.4 c,a,b,d,e,h | 2.3 ± 1.1 c,a | 1.2 ± 0.3 c,b,d,f,g,h | 11.3 ± 2.9 a | 88.5 ± 5.9 c | |
| 17−18 | 7 | 48.3 ± 11.1 d,g,h | 37.9 ± 5.8 d,e,f,h | 4.0 ± 1.2 d,a,c,e,h | Tr d | 0.4 ± 0.1 d,c,f,g,h | Tr b | 66.0 ± 5.1 d | |
| Maltose | Free cells | ||||||||
| 1−8 | 28 | 15.5 ± 3.4 e,a,c,f | 15.3 ± 4.7 e,a | 4.2 ± 2.1 a | 2.4 ± 0.8 e,a,b,c,f | 3.5 ± 1.0 e,a,c,f | 14.7 ± 3.2 e,a,b,c,d | 61.8 ± 5.1 e,c,d | |
| 9−13 | 21 | 14.5 ± 4.9 f,a,c,e | 23.3 ± 6.0 f,b | 6.0 ± 3.8 a | 1.2 ± 0.4 f,b,c,d,e,g,h | 1.7 ± 1.7 f,b,c,d,e,g,h | 5.6 ± 5.2 f,a,b,c,g,h | 45.2 ± 6.8 f | |
| 14−16 | 14 | 31.1 ± 4.2 g,b,d,f,h | 4.5 ± 1.2 g,c,d,h | 5.6 ± 1.3 a | Tr g,b,c,d,f,h | Tr g,d,f,h | Tr g,f,h | 30.2 ± 3.9 g,h | |
| 17−18 | 7 | 26.5 ± 4.6 h,b,c,g | 5.1 ± 1.3 h,c,d,g | 5.0 ± 1.4 a | Tr h,c,d,f,g | Tr h,d,f,g | Tr h,f,g | 32.7 ± 5.3 h,g | |
| Immobilized cells | |||||||||
| 1−8 | 28 | 17.1 ± 5.8 a,c,e,f | 16.0 ± 2.9 a,e | 5.8 ± 1.8 a | 3.0 ± 1.5 a,b,e | 4.0 ± 1.0 a,b,c,e | 13.3 ± 4.6 a,b,c,d,e | 96.8 ± 7.7 a | |
| 9−13 | 21 | 26.2 ± 5.7 b,c,g,h | 27.2 ± 4.9 b,f | 8.0 ± 1.9 a | 1.6 ± 1.0 b,a,c,d,e,f,g | 2.5 ± 1.3 b,a,c,e,f | 9.4 ± 4.1 b,a,c,e,f | 113.5 ± 8.9 b | |
| 14−16 | 14 | 21.2 ± 4.2 c,a,b,e,h | 2.4 ± 0.5 c,d,g,h | 7.3 ± 1.1 a | 0.8 ± 0.2 c,b,d,e,f,g,h | 2.5 ± 1.0 c,a,b,e,f | 10.7 ± 2.3 c,a,b,e,f | 71.8 ± 5.1 c,d,e | |
| 17−18 | 7 | 36.4 ± 6.1 d | 6.0 ± 1.1 d,c,g,h | 3.9 ± 1.0 a | Tr d,b,c,f,g,h | Tr d,f,g,h | 18.7 ± 5.6 d,a,e | 63.6 ± 5.4 d,c,e | |
| Compound | 28 °C | 21 °C | 14 °C | 7 °C | Total Compounds Detected | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GI | GF | SI | SF | MI | MF | GI | GF | SI | SF | MI | MF | GI | GF | SI | SF | MI | MF | GI | GF | SI | SF | MI | MF | ||
| Alcohols | 50 | 18 | 44 | 13 | 41 | 18 | 41 | 17 | 29 | 17 | 32 | 16 | 37 | 14 | 26 | 13 | 33 | 15 | 24 | 15 | 24 | 13 | 26 | 14 | 57 |
| terpenoids | 27 | 2 | 26 | 1 | 25 | 2 | 21 | 1 | 16 | 2 | 17 | 2 | 18 | 2 | 13 | 1 | 14 | 2 | 11 | 1 | 11 | 1 | 11 | 1 | 30 |
| Esters | 16 | 9 | 15 | 10 | 12 | 13 | 15 | 11 | 16 | 11 | 16 | 13 | 13 | 6 | 16 | 9 | 11 | 9 | 16 | 10 | 15 | 10 | 16 | 10 | 24 |
| terpenoids | 3 | 1 | 2 | - | 2 | 1 | 3 | 1 | 3 | 1 | 3 | 1 | 2 | 1 | 2 | - | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 3 |
| Organic acids | 10 | 9 | 10 | 10 | 9 | 10 | 10 | 10 | 9 | 7 | 7 | 9 | 2 | 4 | 5 | 5 | 2 | 3 | 4 | 2 | 6 | 4 | 2 | 4 | 10 |
| terpenoids | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Aldehydes | 6 | 2 | 3 | 2 | 5 | 1 | 11 | 3 | 9 | 2 | 12 | 1 | 4 | 1 | 5 | 3 | 3 | 1 | 2 | 1 | 3 | 1 | 3 | 1 | 13 |
| terpenoids | 4 | - | 2 | - | 4 | - | 3 | - | 2 | - | 3 | - | 2 | - | 2 | - | 2 | - | 2 | - | 2 | - | 2 | - | 4 |
| Ketones | 13 | 6 | 17 | 4 | 15 | 6 | 13 | 7 | 10 | 7 | 12 | 6 | 13 | 8 | 10 | 5 | 10 | 8 | 12 | 7 | 10 | 7 | 10 | 6 | 21 |
| terpenoids | 8 | - | 8 | - | 8 | - | 6 | - | 3 | - | 5 | - | 5 | - | 3 | - | 3 | - | 4 | - | 4 | - | 4 | - | 9 |
| Miscellaneous compounds | 16 | 10 | 18 | 7 | 15 | 9 | 16 | 10 | 18 | 8 | 19 | 8 | 12 | 5 | 12 | 8 | 13 | 10 | 16 | 10 | 14 | 10 | 15 | 11 | 22 |
| terpenoids | 8 | 1 | 8 | - | 8 | 1 | 7 | 1 | 8 | - | 8 | 1 | 4 | - | 4 | - | 5 | 1 | 7 | 1 | 7 | 1 | 7 | 1 | 11 |
| Total terpenoids | 50 | 4 | 46 | 1 | 47 | 4 | 40 | 3 | 32 | 3 | 36 | 4 | 31 | 3 | 24 | 1 | 26 | 4 | 26 | 3 | 26 | 3 | 26 | 3 | 57 |
| Total compounds | 111 | 54 | 107 | 46 | 97 | 57 | 106 | 58 | 91 | 52 | 98 | 53 | 81 | 38 | 74 | 43 | 72 | 46 | 74 | 45 | 72 | 36 | 72 | 46 | 147 |
| Compounds | KI | G7-I | G7-F | G14-I | G14-F | G21-I | G21-F | G28-I | G28-F | S7-I | S7-F | S14-I | S14-F | S21-I | S21-F | S28-I | S28-F | M7-I | M7-F | M14-I | M14-F | M21-I | M21-F | M28-I | M28-F |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p-menth-4(8)-ene | 946 | 23.90 | 18.36 | - | - | 24.86 | 18.57 | 1.18 | 5.45 | 25.00 | 1.17 | 1.46 | - | 1.24 | - | 1.66 | - | 1.84 | 0.97 | 0.81 | 1.14 | 1.28 | - | ||
| α-pinene | 994 | 23.30 | - | 47.47 | - | 51.39 | - | 7.79 | - | 386.54 | - | 1.92 | - | 14.35 | - | 9.98 | - | 1.90 | - | 4.83 | - | 13.07 | - | 36.24 | - |
| carvomenthol | 1024 | - | - | - | - | - | - | 5.38 | - | - | - | - | - | - | - | 4.84 | - | - | - | - | - | - | - | 9.30 | - |
| camphene | 1033 | 1.93 | - | - | - | - | - | - | - | 6.50 | - | - | - | - | - | - | - | 5.86 | - | - | - | - | - | - | - |
| trans-pinane | 1038 | - | - | - | - | - | - | 15.17 | - | - | - | - | - | 15.06 | - | 1.16 | - | - | - | - | - | 0.40 | - | 0.65 | - |
| β-pinene | 1085 | 0.52 | - | - | - | 0.13 | - | 0.06 | - | 8.79 | - | - | - | 0.93 | - | 1.03 | - | 3.77 | - | 0.12 | - | 0.54 | - | 0.33 | - |
| thuja-2,4(10)-diene | 1119 | 0.41 | - | - | - | - | - | - | - | 2.60 | - | - | - | - | - | - | - | 1.19 | - | - | - | - | - | - | - |
| 1,8-cineole | 1205 | - | - | - | - | 0.27 | - | 5.19 | - | - | - | - | - | 0.33 | - | 1.09 | - | - | - | - | - | 0.53 | - | 0.75 | - |
| p-menth-1-en-9-al | 1235 | - | - | - | - | 0.24 | - | 3.70 | - | - | - | - | - | - | - | 0.33 | - | - | - | - | - | 4.48 | - | 3.71 | - |
| verbenyl ethyl ether | 1333 | 4.06 | - | 4.80 | - | 0.20 | - | 5.30 | - | 15.09 | - | 0.31 | - | 1.11 | - | 0.55 | - | 0.76 | - | 0.29 | - | 2.02 | - | 1.43 | - |
| p-cymenene | 1410 | - | - | - | - | 0.24 | - | 0.27 | - | - | - | - | - | 0.35 | - | 1.09 | - | - | - | - | - | 1.05 | - | 2.88 | - |
| dehydro-p-cymene | 1438 | 4.96 | - | 2.68 | - | 3.55 | - | 3.42 | - | 14.94 | - | 1.53 | - | 199.91 | - | 5.95 | - | 9.38 | - | 0.62 | - | 2.37 | - | 18.81 | - |
| linalool oxide | 1464 | - | - | 2.13 | - | 0.85 | - | 0.51 | - | - | - | - | - | 0.39 | - | 0.43 | - | - | - | 0.21 | - | 0.33 | - | 0.29 | - |
| α-campholenal | 1484 | 1.21 | - | 0.09 | - | 0.17 | - | 0.15 | - | 1.46 | - | 0.31 | - | 0.31 | - | - | - | 0.18 | - | 0.24 | - | 1.41 | - | 0.78 | - |
| pinocarvone | 1521 | 2.01 | - | 2.58 | - | - | - | 2.39 | - | 5.46 | - | - | - | 1.27 | - | 1.73 | - | 39.62 | - | 0.25 | - | 1.05 | - | 2.95 | - |
| α-norborneol | 1525 | - | - | - | - | - | - | 0.34 | - | - | - | - | - | - | - | 0.23 | - | - | - | - | - | - | - | - | - |
| linalool | 1536 | 1.21 | - | 2.69 | - | 0.15 | - | 0.97 | - | 1.35 | - | 0.95 | - | 1.09 | - | 0.32 | - | 0.58 | - | 0.13 | - | 0.66 | - | 0.60 | - |
| fenchol | 1541 | - | - | 4.79 | - | 1.88 | - | 2.39 | - | - | - | 1.06 | - | 0.90 | - | 1.13 | - | - | - | 0.35 | - | 1.93 | - | 2.95 | - |
| bornyl acetate | 1550 | 5.98 | - | 6.10 | - | 0.63 | - | 0.86 | - | 15.65 | - | 1.04 | - | 2.00 | - | 0.94 | - | 0.90 | - | 0.33 | - | 1.57 | - | 1.44 | - |
| isopinocamphone | 1557 | - | - | 1.56 | - | - | - | 1.56 | - | - | - | 11.81 | - | - | - | 0.83 | - | - | - | - | - | 0.69 | - | 0.97 | - |
| 4-terpineol | 1562 | 2.10 | - | 3.47 | - | 2.48 | - | 5.83 | - | 3.18 | - | 1.21 | - | 1.25 | - | 4.88 | - | 5.17 | - | 0.36 | - | 2.76 | - | 3.71 | - |
| myrtenal | 1588 | 2.41 | - | 0.72 | - | 0.55 | - | 0.35 | - | 7.03 | - | 0.49 | - | 1.59 | - | - | - | 0.19 | - | 0.42 | - | 1.49 | - | 2.69 | - |
| 3,6,6-trimethyl-2-norpinanone | 1595 | - | - | 1.13 | - | 0.62 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1.09 | - |
| L-trans-pinocarveol | 1618 | 3.41 | - | 2.23 | - | 2.00 | - | 5.70 | - | 7.74 | - | 0.12 | - | 2.50 | - | 4.64 | - | 3.18 | - | 0.31 | - | 2.79 | - | 4.85 | - |
| α-phellandren-8-ol | 1626 | 0.86 | - | 0.93 | - | 0.41 | - | 1.27 | - | 3.45 | - | 4.62 | - | 3.97 | - | 5.83 | - | 1.46 | - | 0.17 | - | 1.29 | - | 1.73 | - |
| δ-terpineol | 1636 | - | - | - | - | - | - | 0.11 | - | - | - | - | - | - | - | 0.16 | - | - | - | - | - | - | - | 0.14 | - |
| bornylene | 1647 | - | - | 0.51 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| neral | 1652 | - | - | - | - | - | - | 0.50 | - | - | - | - | - | - | - | 0.24 | - | - | - | - | - | - | - | 0.34 | - |
| α-terpineol | 1665 | 8.32 | - | 11.89 | 0.48 | 17.40 | - | 34.65 | 0.43 | 15.09 | - | 8.50 | - | 6.92 | 0.52 | 23.67 | - | 7.08 | - | 3.96 | 0.23 | 17.96 | 1.16 | 22.93 | 0.66 |
| borneol | 1668 | 0.80 | - | 1.28 | - | 2.72 | - | 2.61 | - | 1.22 | - | 1.99 | - | 1.86 | - | 2.10 | - | 2.82 | - | 0.87 | - | 5.76 | - | 29.15 | - |
| isopinocampheol | 1675 | - | - | 1.20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| verbenol | 1679 | 3.59 | - | 1.84 | - | 0.84 | - | 1.59 | - | 16.76 | - | 0.87 | - | 1.29 | - | 2.36 | - | 1.17 | - | 0.78 | - | 0.61 | - | 0.52 | - |
| endo,endo-2,3-bornanediol | 1688 | - | - | 0.41 | - | 0.20 | - | 0.23 | - | - | - | - | - | 0.17 | - | 0.21 | - | - | - | - | - | - | - | 0.23 | - |
| exo-hydroxycineol | 1696 | 0.97 | - | 2.14 | - | 2.82 | - | 6.65 | - | 1.85 | - | 0.63 | - | 6.79 | - | 5.00 | - | 0.22 | - | 0.28 | - | 1.91 | - | 2.29 | - |
| myrcenol | 1696 | - | - | - | - | - | - | - | - | - | - | - | - | 1.41 | - | - | - | - | - | - | - | - | - | - | - |
| verbenone | 1705 | 4.58 | - | 2.53 | - | 2.73 | - | 4.64 | - | 17.00 | - | 0.87 | - | 2.96 | - | 3.94 | - | 4.82 | - | 1.18 | - | 0.31 | - | 0.14 | - |
| carvone | 1707 | 0.56 | - | - | - | 0.21 | - | 0.56 | - | 1.33 | - | - | - | - | - | 0.62 | - | 0.37 | - | - | - | 0.54 | - | 0.85 | - |
| p-methyl-acetophenone | 1754 | 1.55 | - | 1.79 | - | 1.71 | - | 2.07 | - | 3.84 | - | 1.19 | - | 2.02 | - | 2.19 | - | 2.81 | - | 0.71 | - | 2.94 | - | 2.88 | - |
| β-phellandren-8-ol | 1757 | - | - | 0.47 | - | 0.71 | - | 1.50 | - | - | - | - | - | - | - | 0.77 | - | - | - | - | - | 0.31 | - | 0.59 | - |
| myrtenol | 1767 | 5.59 | - | 3.20 | - | 3.95 | - | 3.54 | - | 8.70 | - | 1.05 | - | 2.90 | - | 3.69 | - | 5.92 | - | 1.09 | - | 2.71 | - | 1.42 | - |
| trans-carveol | 1812 | 3.45 | - | 2.67 | - | 1.88 | - | 2.44 | - | 6.62 | - | 4.66 | - | 2.60 | - | 3.09 | - | 5.31 | - | 1.79 | - | 2.18 | - | 2.19 | - |
| geraniol | 1821 | - | - | - | - | - | - | 0.15 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| p-cymen-8-ol | 1828 | - | - | - | - | 8.81 | - | 5.26 | - | - | - | 2.21 | - | - | - | 1.41 | - | - | - | 1.21 | - | 6.86 | - | 9.17 | - |
| geranyl acetate | 1835 | - | - | - | - | 0.28 | - | 0.56 | - | - | - | - | - | 0.09 | - | - | - | - | - | - | - | 0.88 | - | - | - |
| cis-carveol | 1845 | - | - | 3.15 | - | - | - | 5.97 | - | - | - | - | - | - | - | 6.93 | - | - | - | - | - | - | - | 6.31 | - |
| trans-myrtanol | 1854 | - | - | - | - | 2.18 | - | 1.19 | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.98 | - | 1.17 | - |
| cis-myrtanol | 1855 | - | - | 0.60 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| p-cymen-9-ol | 1878 | - | - | - | - | 0.80 | - | 0.70 | - | - | - | - | - | - | - | 0.71 | - | - | - | - | - | - | - | 0.91 | - |
| piperitenone | 1917 | - | - | - | - | 0.45 | - | 0.61 | - | - | - | - | - | - | - | 0.52 | - | - | - | - | - | - | - | 0.71 | - |
| geranyl isovalerate | 1957 | 1.50 | 1.08 | 1.12 | 0.82 | 0.15 | 0.91 | 1.48 | 0.72 | 0.72 | 0.23 | 0.09 | - | 0.65 | 1.17 | 1.44 | - | 0.18 | 0.13 | 0.09 | 0.12 | 0.28 | 0.33 | 0.24 | 0.30 |
| 3,6,6-trimethyl-2-norpinanol | 1992 | 1.25 | 0.89 | 1.39 | 1.03 | 0.59 | 1.96 | 1.44 | 0.72 | 1.10 | 0.50 | 0.13 | 0.35 | 2.52 | 1.13 | 3.54 | 1.23 | 0.64 | 0.66 | 1.32 | 0.87 | 0.42 | 1.51 | 1.08 | 1.14 |
| β-camphor | 2038 | - | - | - | - | - | - | 0.53 | - | - | - | - | - | - | - | 0.75 | - | - | - | - | - | - | - | - | - |
| p-cresol | 2071 | - | - | - | - | 0.38 | - | 0.86 | - | - | - | - | - | - | - | 1.05 | - | - | - | - | - | 0.98 | - | 0.75 | - |
| cuminyl alcohol | 2090 | - | - | - | - | 0.63 | - | 0.71 | - | - | - | - | - | - | - | 0.82 | - | - | - | - | - | - | - | 1.09 | - |
| pseudoionone | 2126 | - | - | - | - | 0.46 | - | 0.39 | - | - | - | - | - | - | - | 0.46 | - | - | - | - | - | - | - | 0.29 | - |
| longipinanol | 2140 | - | - | - | - | - | - | 0.22 | - | - | - | - | - | - | - | 0.13 | - | - | - | - | - | - | - | 0.23 | - |
| farnesol | 2323 | - | - | - | - | 0.55 | - | 0.57 | - | - | - | - | - | 0.40 | - | 0.43 | - | - | - | - | - | - | - | 0.74 | - |
| Total terpenes | 110.43 | 20.33 | 119.56 | 2.33 | 141.07 | 21.44 | 151.51 | 7.32 | 579.01 | 1.9 | 49.02 | 0.35 | 281.13 | 2.82 | 114.87 | 1.23 | 107.32 | 1.76 | 22.72 | 2.36 | 87.34 | 3.0 | 184.51 | 2.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kallis, M.; Sideris, K.; Kopsahelis, N.; Bosnea, L.; Kourkoutas, Y.; Terpou, A.; Kanellaki, M. Pistacia terebinthus Resin as Yeast Immobilization Support for Alcoholic Fermentation. Foods 2019, 8, 127. https://doi.org/10.3390/foods8040127
Kallis M, Sideris K, Kopsahelis N, Bosnea L, Kourkoutas Y, Terpou A, Kanellaki M. Pistacia terebinthus Resin as Yeast Immobilization Support for Alcoholic Fermentation. Foods. 2019; 8(4):127. https://doi.org/10.3390/foods8040127
Chicago/Turabian StyleKallis, Michalis, Konstantinos Sideris, Nikolaos Kopsahelis, Loulouda Bosnea, Yiannis Kourkoutas, Antonia Terpou, and Maria Kanellaki. 2019. "Pistacia terebinthus Resin as Yeast Immobilization Support for Alcoholic Fermentation" Foods 8, no. 4: 127. https://doi.org/10.3390/foods8040127
APA StyleKallis, M., Sideris, K., Kopsahelis, N., Bosnea, L., Kourkoutas, Y., Terpou, A., & Kanellaki, M. (2019). Pistacia terebinthus Resin as Yeast Immobilization Support for Alcoholic Fermentation. Foods, 8(4), 127. https://doi.org/10.3390/foods8040127