Variation in Pea (Pisum sativum L.) Seed Quality Traits Defined by Physicochemical Functional Properties

 ,
,  ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Physicochemical Characteristics and Cooking Time
2.3. Pasting Properties, Resistant Starch, and Basic Composition
2.4. Seed Trait Classification
2.5. Statistical Analysis
3. Results and Discussion
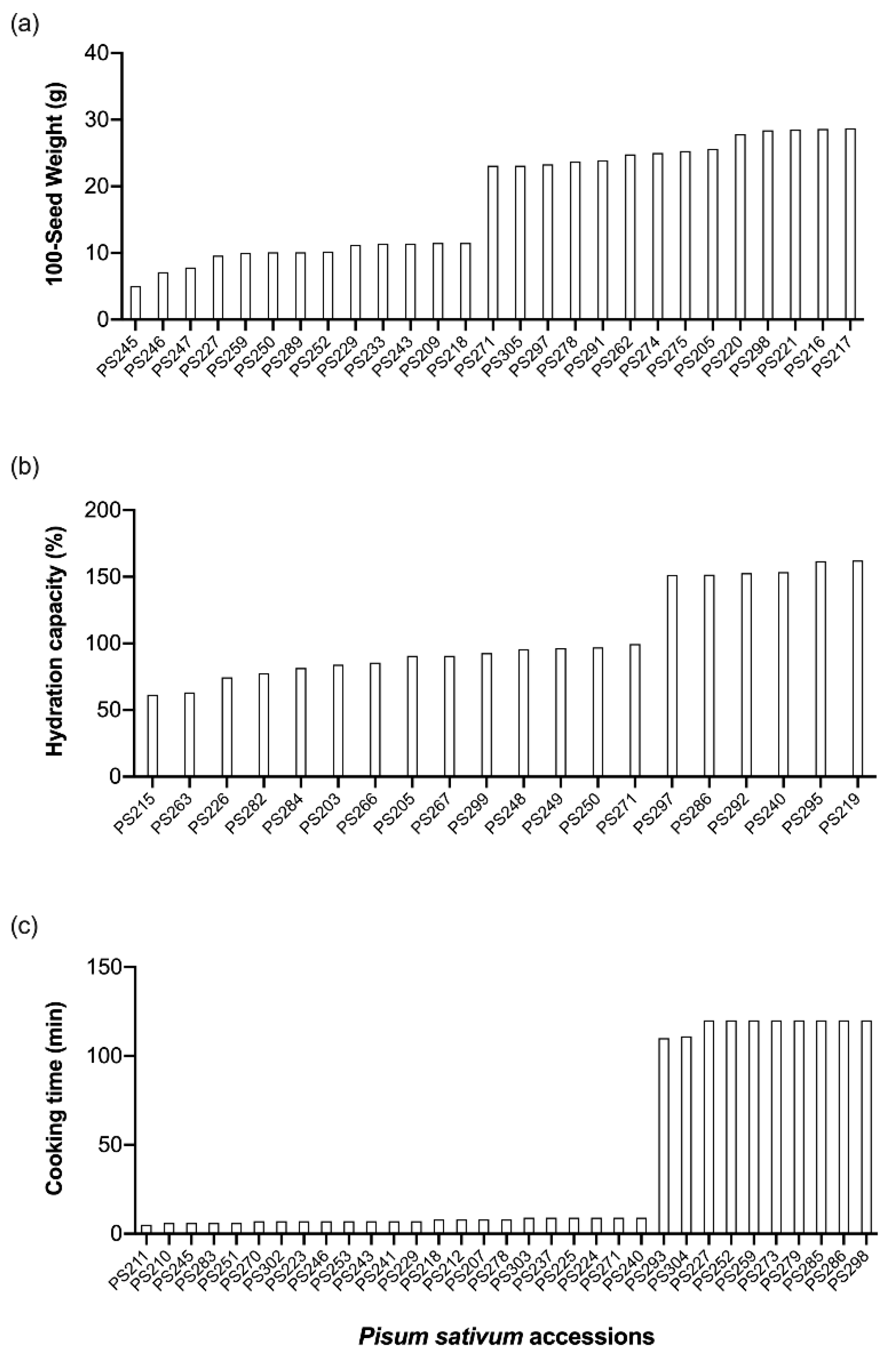
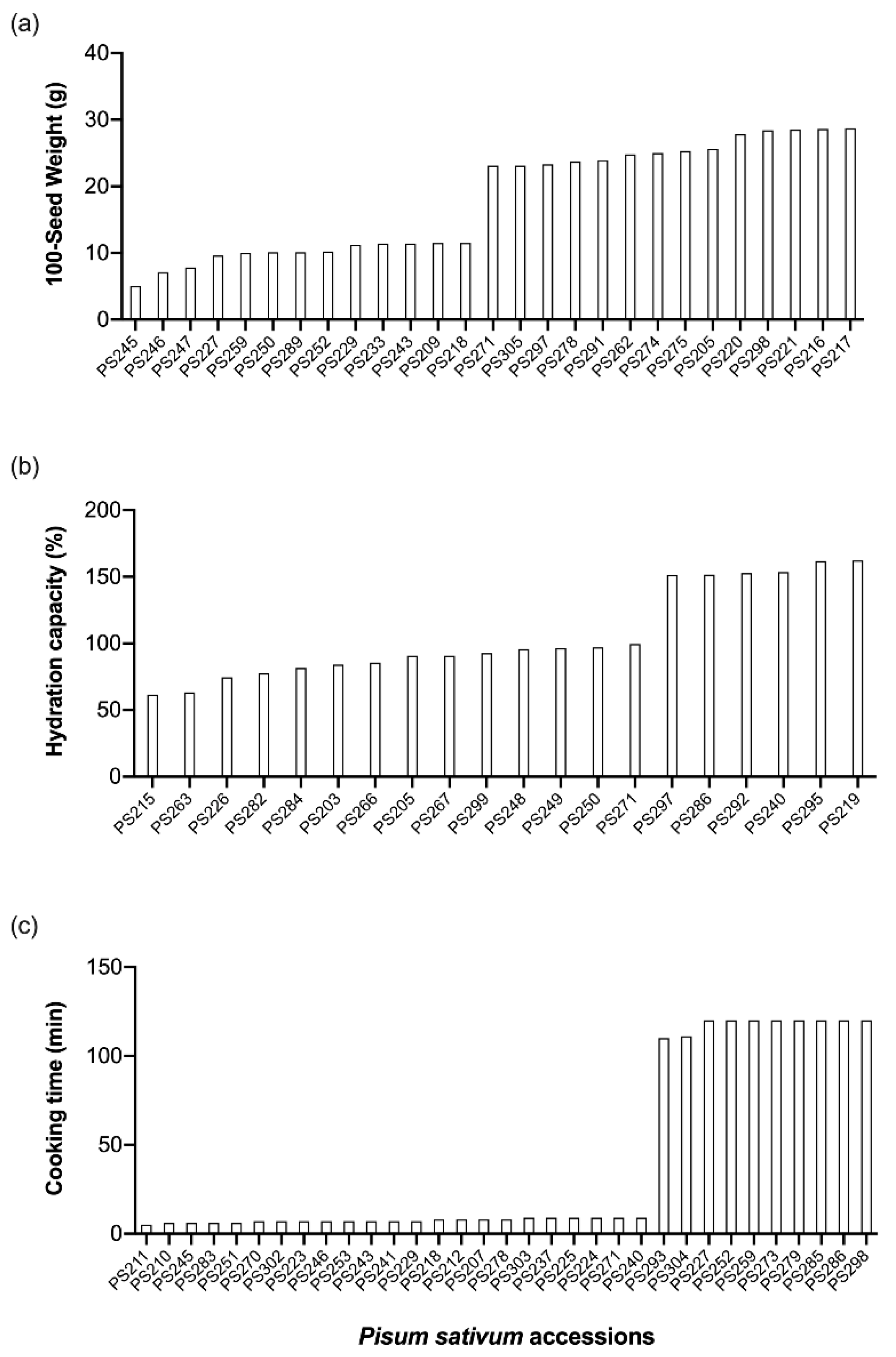
3.1. Seed and Flour Variation in Pea Germplasm
3.2. Correlation Analysis of Pasting, Physicochemical, and Basic Composition Parameters
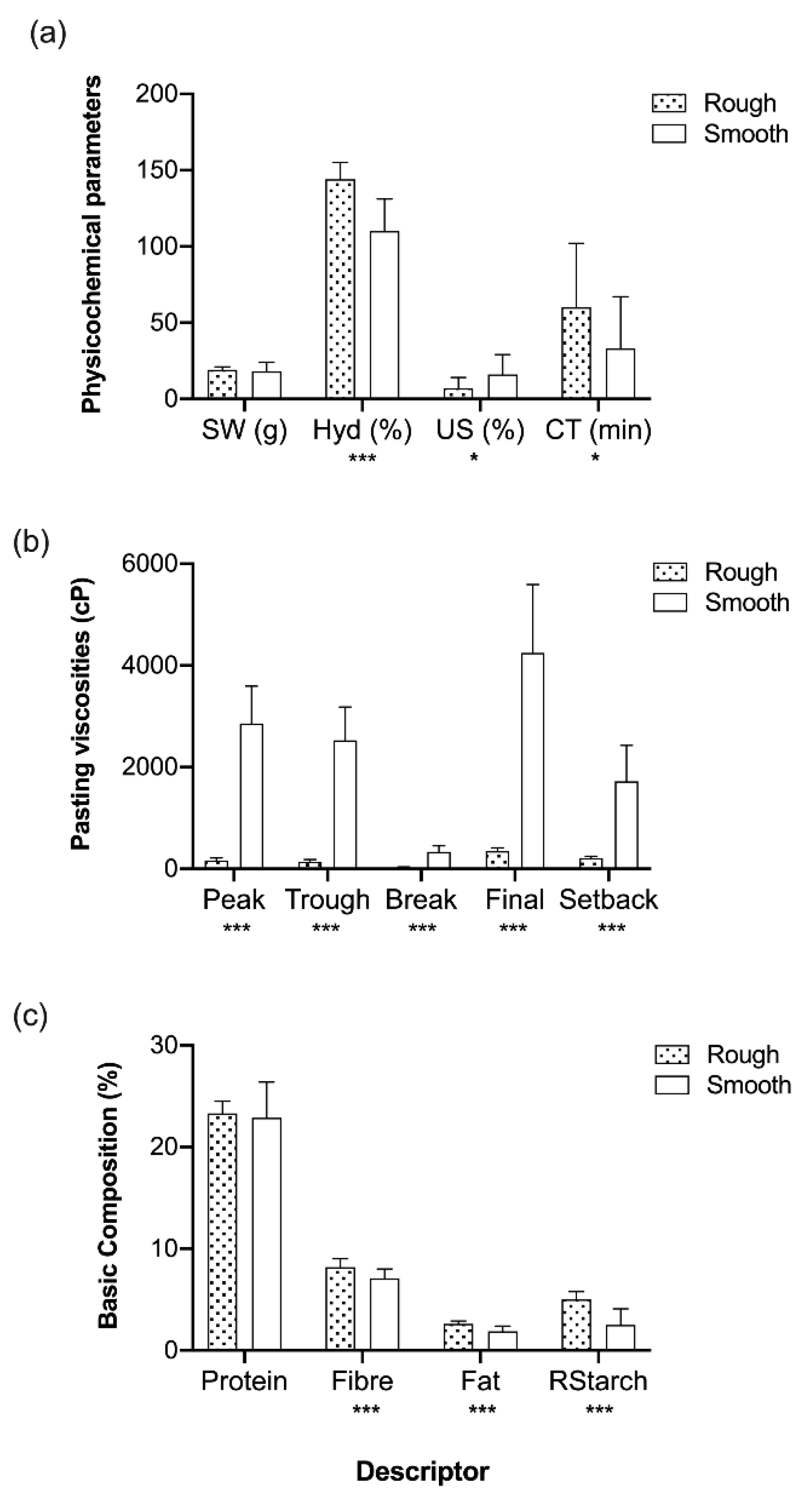
3.3. Seed Trait Variation
3.4. Characterisation of Allelic Variation at the r and rb Genetic Loci
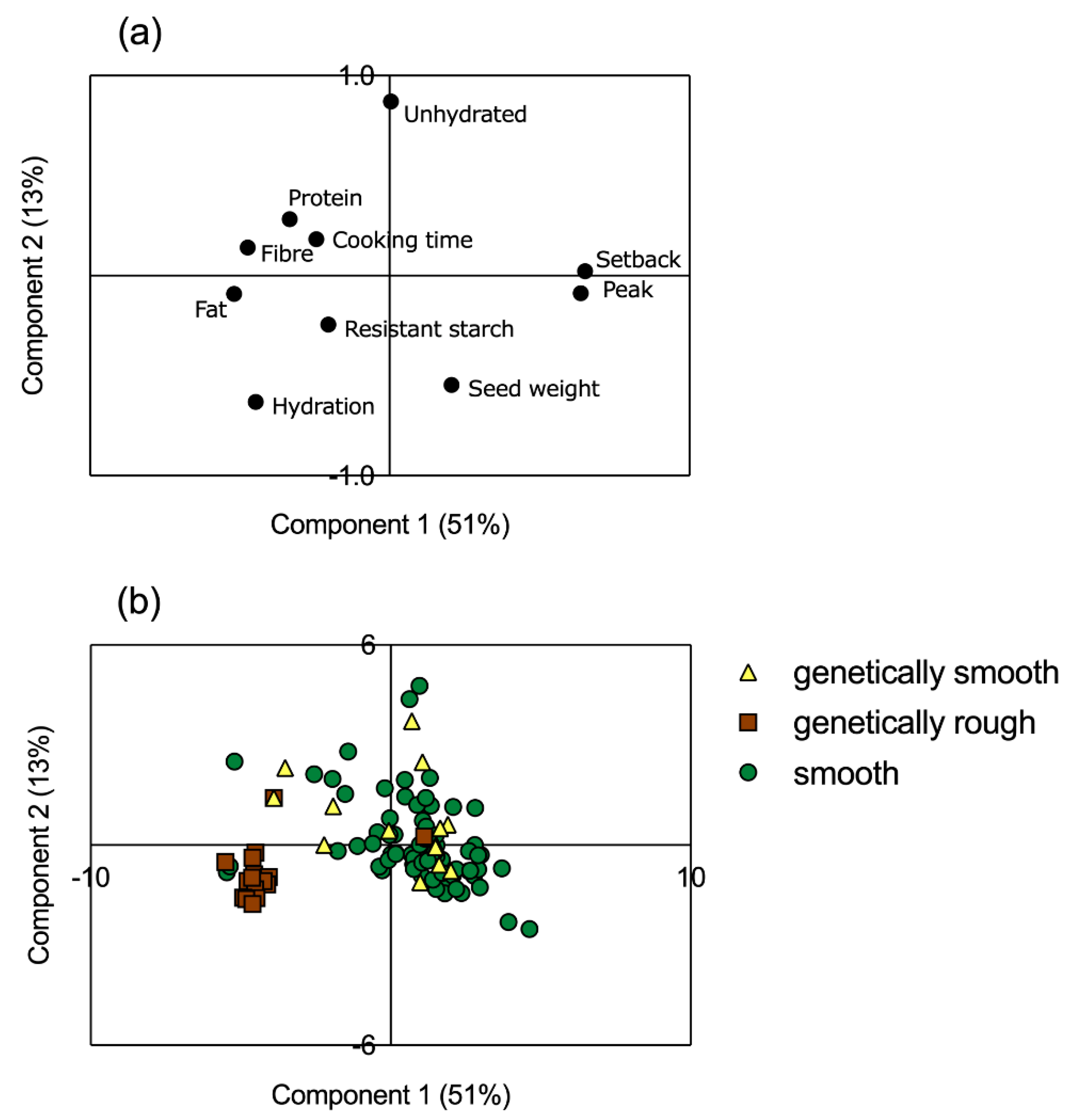
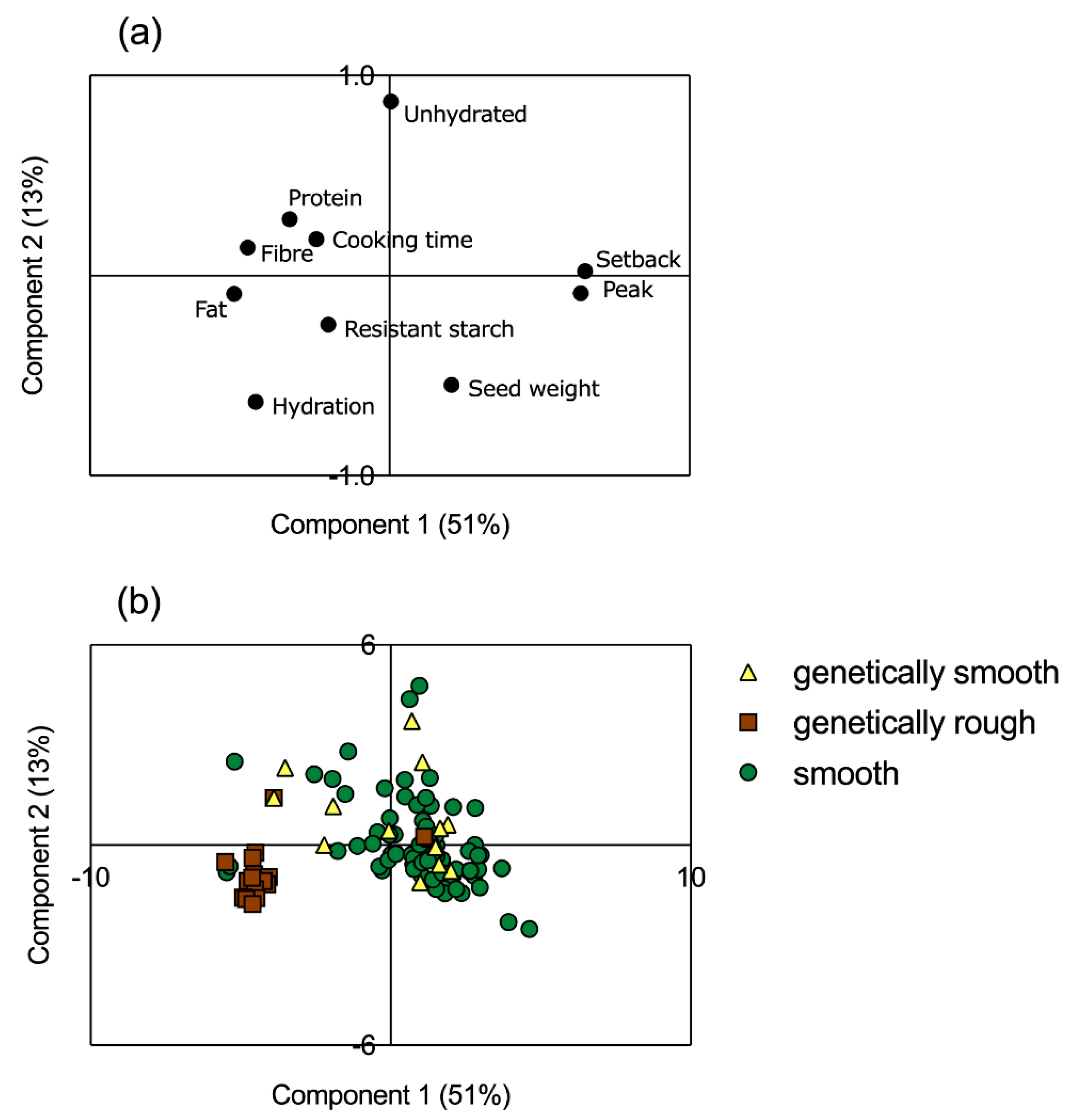
3.5. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Peas, Dry, World’s Area Harvested and Production Quantity. Available online: http://www.fao.org/faostat/en/?#data/QC (accessed on 4 November 2019).
- Eurostat. Eurostat Statistics Explained. Dry Pulses in EU Agriculture—Statistics on Cultivation, Production and Economic Value. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Dry_pulses_in_EU_agriculture_-_statistics_on_cultivation,_production_and_economic_value#Structure_of_production (accessed on 10 April 2019).
- Foschia, M.; Horstmann, S.W.; Arendt, E.K.; Zannini, E. Legumes as functional ingredients in gluten-free bakery and pasta products. Annu. Rev. Food Sci. Technol. 2017, 8, 75–96. [Google Scholar] [CrossRef]
- Rempel, C.; Geng, X.; Zhang, Y. Industrial scale preparation of pea fractions with enhanced nutritive composition by dry fractionation. Food Chem. 2019, 276, 119–128. [Google Scholar] [CrossRef]
- Cunha, L.M.; Fonseca, S.C.; Lima, R.C.; Loureiro, J.; Pinto, A.S.; Vaz Patto, M.C.; Brites, C. Consumer-driven improvement of maize bread formulations with legume fortification. Foods 2019, 8, 235. [Google Scholar] [CrossRef]
- Tosh, S.M.; Farnworth, E.R.; Brummer, Y.; Duncan, A.M.; Wright, A.J.; Boye, J.I.; Marcotte, M.; Benali, M. Nutritional profile and carbohydrate characterization of spray-dried lentil, pea and chickpea ingredients. Foods 2013, 2, 338–349. [Google Scholar] [CrossRef]
- Vaz Patto, M.C.; Amarowicz, R.; Aryee, A.N.A.; Boye, J.I.; Chung, H.J.; Martín-Cabrejas, M.A.; Domoney, C. Achievements and challenges in improving the nutritional quality of food legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Shevkani, K.; Singh, N.; Kaur, A. Bioactive constituents in pulses and their health benefits. J. Food Sci. Technol. 2017, 54, 858–870. [Google Scholar] [CrossRef]
- Santos, C.S.; Carbas, B.; Castanho, A.; Bronze, M.R.; Serrano, C.; Vasconcelos, M.W.; Vaz Patto, M.C.; Brites, C. Relationship between seed traits and pasting and cooking behaviour in a pulse germplasm collection. Crop Pasture Sci. 2018, 69, 892–903. [Google Scholar] [CrossRef]
- Tripathi, S.; Sridhar, V.; Jukanti, A.K.; Suresh, K.; Rao, B.V.; Gowda, C.L.L.; Gaur, P.M. Genetic variability and interrelationships of phenological, physicochemical and cooking quality traits in chickpea. Plant Genet. Resour. Charact. Util. 2012, 10, 194–201. [Google Scholar] [CrossRef]
- Serrano, C.; Carbas, B.; Castanho, A.; Soares, A.; Vaz Patto, M.C.; Brites, C. Characterisation of nutritional quality traits of a chickpea (Cicer arietinum) germplasm collection exploited in chickpea breeding in Europe. Crop Pasture Sci. 2017, 68, 1031–1040. [Google Scholar] [CrossRef]
- Wang, S.; Sharp, P.; Copeland, L. Structural and functional properties of starches from field peas. Food Chem. 2011, 126, 1546–1552. [Google Scholar] [CrossRef]
- Simsek, S.; Tulbek, M.C.; Yao, Y.; Schatz, B. Starch characteristics of dry peas (Pisum sativum L.) grown in the USA. Food Chem. 2009, 115, 832–838. [Google Scholar] [CrossRef]
- Liu, C.; Wang, S.; Copeland, L.; Wang, S. Physicochemical properties and in vitro digestibility of starches from field peas grown in China. LWT Food Sci. Technol. 2015, 64, 829–836. [Google Scholar] [CrossRef]
- Maharjan, P.; Penny, J.; Partington, D.L.; Panozzo, J.F. Genotype and environment effects on the chemical composition and rheological properties of field peas. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Kumar, A.; Mamidi, S.; McPhee, K. Genetic diversity and population structure among pea (Pisum sativum L.) cultivars as revealed by simple sequence repeat and novel genic markers. Mol. Biotechnol. 2014, 56, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Rayner, T.; Moreau, C.; Ambrose, M.; Isaac, P.G.; Ellis, N.; Domoney, C. Genetic variation controlling wrinkled seed phenotypes in Pisum: How lucky was Mendel? Int. J. Mol. Sci. 2017, 18, 1205. [Google Scholar] [CrossRef] [PubMed]
- AACC International. AACC Approved Methods of Analysis: Method 56-35.01—Method for Determining Water Hydration Capacity and Percentage of Unhydrated Seeds of Pulses, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, October 2007. [Google Scholar]
- AACC International. AACC Approved Methods of Analysis: Method 76-21.02—General Pasting Method for Wheat or Rye Flour or Starch Using the Rapid Visco Analyser, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, November 1999. [Google Scholar]
- AACC International. AACC Approved Methods of Analysis: Method 32-40.01—Resistant Starch in Starch Samples and Plant Materials, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, October 2002. [Google Scholar]
- International Organization for Standardizatio (ISO). ISO 16634. Food Products—Determination of the Total Nitrogen Content by Combustion According to Dumas Principle and Calculation of the Crude Protein Content; International Organization for Standardization: London, UK, 2016. [Google Scholar]
- International Organization for Standardizatio (ISO). ISO 6492. Animal Feeding Stuffs—Determination of Fat Content; International Organization for Standardization: London, UK, 1999. [Google Scholar]
- International Organization for Standardizatio (ISO). ISO 6865. Animal Feeding Stuffs—Determination of Crude Fibre Content. Method with Intermediate Filtration; International Organization for Standardization: London, UK, 2000. [Google Scholar]
- Rakotomalala, R. TANAGRA: Un Logiciel Gratuit Pour L’enseignement et la Recherche; Actes de EGC ’2005RNTI-E-3; EGC: Lyon, France, 2005; Volume 2, pp. 697–702. [Google Scholar]
- Sharma, S.; Singh, N.; Virdi, A.S.; Rana, J.C. Quality traits analysis and protein profiling of field pea (Pisum sativum) germplasm from Himalayan region. Food Chem. 2015, 172, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Khatkar, B.S. Thermal, pasting and morphological properties of starch granules of wheat (Triticum aestivum L.) varieties. J. Food Sci. Technol. 2017, 54, 2403–2410. [Google Scholar] [CrossRef]
- Adebowale, A.R.A.; Sanni, S.A.; Oladapo, F.O. Chemical, functional and sensory properties of instant yam-breadfruit flour. Niger. Food J. 2008, 26, 2–12. [Google Scholar] [CrossRef]
- Li, L.; Yuan, T.Z.; Setia, R.; Raja, R.B.; Zhang, B. Characteristics of pea, lentil and faba bean starches isolated from air-classified flours in comparison with commercial starches. Food Chem. 2019, 276, 599–607. [Google Scholar] [CrossRef]
- Nikolopoulou, D.; Grigorakis, K.; Stasini, M.; Alexis, M.N.; Iliadis, K. Differences in chemical composition of field pea (Pisum sativum) cultivars: Effects of cultivation area and year. Food Chem. 2007, 103, 847–852. [Google Scholar] [CrossRef]
- Ratnayake, W.S.; Hoover, R.; Warkentin, T. Pea starch: Composition, structure and properties—A Review. Starch 2002, 54, 217–234. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Martini, M.; Saporita, T.C.; Nucci, D.; Gianfredi, V.; Maddalo, F.; Capua, A.D.; Tovani, F.; Marensi, L. Chapter 17—Nutraceutical and functional food regulations in the European Union. In Developing New Functional Food and Nutraceutical Products; Bagchi, D., Nair, S., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 309–322. [Google Scholar]
- Abdel-Aal, E.M.; Ragaee, S.; Rabalski, I.; Warkentin, T.; Vandenberg, A. Nutrient content and ciscosity of Saskatchewan-grown pulses in relation to their cooking quality. Can. J. Plant Sci. 2019, 99, 67–77. [Google Scholar] [CrossRef]
- Carbas, B.; Pathania, S.; Castanho, A.; Lourenço, D.; Veiga, I.M.; Vaz Patto, M.C.; Brites, C. Elucidating potential utilization of Portuguese commnon bean varieties in rice based processed foods. J. Food Sci. Technol. 2018, 55, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.; Zhang, L.; Arntfield, S. Understanding water uptake from the induced changes occurred during processing: Chemistry of pinto and navy bean seed coats. Int. J. Food Prop. 2010, 13, 631–647. [Google Scholar] [CrossRef]
- Bhattacharyya, M.K.; Smith, A.M.; Ellis, T.H.N.; Hedley, C.; Martin, C. The wrinkled-seed character of pea described by Mendel is caused by a transposon-like insertion in a gene encoding starch-branching enzyme. Cell 1990, 60, 115–122. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptor | Mean ± RSD | Range |
|---|---|---|
| Physicochemical parameters | ||
| 100-seed weight (g) | 18 ± 28 | 5–29 |
| Hydration capacity (%) | 116 ± 17 | 62–162 |
| Unhydrated seeds (%) | 10 ± 88 | 2–50 |
| Cooking time (min) | 33 ± 106 | 5–120 |
| Pasting viscosities (cP *) | ||
| Peak | 2643 ± 50 | 83–4836 |
| Trough | 2358 ± 49 | 58–4066 |
| Breakdown | 285 ± 67 | 8–867 |
| Final | 4018 ± 50 | 212–7471 |
| Setback | 1660 ± 54 | 151–3489 |
| Basic composition (%) | ||
| Protein | 22 ± 0.1 | 16–30 |
| Fibre | 7 ± 0.1 | 6–10 |
| Fat | 2 ± 0.3 | 1–3 |
| Resistant Starch | 3 ± 50 | 1–7 |
| Trough (cP) | Break (cP) | FV (cP) | SB (cP) | SW (g) | HC (%) | US (%) | CT (min) | Protein | Fibre | Fat | RS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Peak | 1.000 ** | 0.804 ** | 0.989 ** | 0.949 ** | 0.218 * | −0.631 ** | −0.005 | −0.391 ** | −0.443 ** | −0.619 ** | −0.759 ** | −0.269 * |
| Trough | 0.743 ** | 0.989 ** | 0.944 ** | 0.192 * | −0.627 ** | −0.009 | −0.397 ** | −0.421 ** | −0.613 ** | −0.761 ** | −0.281 * | |
| Break | 0.764 ** | 0.768 ** | 0.333 ** | −0.518 ** | 0.015 | −0.264 * | −0.475 ** | −0.516 ** | −0.572 ** | −0.136 | ||
| FV | 0.982 ** | 0.249 ** | −0.610 ** | −0.031 | −0.373 ** | −0.495 ** | −0.610 ** | −0.741 ** | −0.263 * | |||
| SB | 0.317 ** | −0.568 ** | −0.059 | −0.329 ** | −0.577 ** | −0.587 ** | −0.690 ** | −0.230 * | ||||
| SW | −0.011 | −0.214 * | −0.007 | −0.408 ** | −0.303 ** | −0.058 | −0.068 | |||||
| HC | −0.578 ** | 0.035 | 0.327 ** | 0.393 ** | 0.493 ** | 0.359 ** | ||||||
| US | 0.138 | 0.139 | 0.118 | 0.013 | −0.055 | |||||||
| CT | −0.130 | 0.362 ** | 0.258 * | 0.146 | ||||||||
| Protein | 0.211 * | 0.113 | 0.040 | |||||||||
| Fibre | 0.666 ** | 0.217 * | ||||||||||
| Fat | 0.132 |
| Peak | Trough | Break | FV | Setback | SW | HC | US | CT | |
|---|---|---|---|---|---|---|---|---|---|
| Shape | |||||||||
| Ellipsoid (n = 42) | 3252 a | 2910 a | 342 a | 4938 a | 2028 a | 17 a | 109 a | 10 a | 22 a |
| Cylindrical (n = 24) | 3066 a | 2726 a | 340 a | 4632 a | 1906 ab | 19 a | 112 a | 11 a | 27 ab |
| Rhomboid (n = 11) | 1953 b | 1716 b | 236 ab | 2944 b | 1228 b | 16 a | 121 ab | 10 a | 51 bc |
| Irregular (n = 16) | 582 c | 496 c | 86 b | 918 c | 422 c | 18 a | 135 b | 13 a | 67 c |
| n.d. (n = 12) | 3046 a | 2759 a | 287 a | 4684 a | 1925 ab | 19 a | 115 a | 9 a | 16 ab |
| Colour | |||||||||
| Cream yellow (n = 9) | 3107 ab | 2765 ab | 342 a | 4727 ab | 1962 ab | 19 a | 111 ab | 8 ab | 11 a |
| Yellow green (n = 24) | 3120 a | 2777 a | 342 a | 4736 a | 1958 a | 19 a | 112 a | 9 a | 23 a |
| Light green (n = 14) | 2200 ab | 1933 ab | 267 a | 3365 ab | 1432 ab | 19 a | 126 ab | 8 a | 34 a |
| Dark green (n = 8) | 2677 ab | 2420 ab | 257 a | 3957 ab | 1537 ab | 11 b | 103 a | 15 ab | 51 a |
| Green (n = 23) | 1744 b | 1548 b | 196 a | 2609 b | 1061 b | 18 a | 128 b | 9 a | 44 a |
| Army green (n = 5) | 3172 ab | 2800 ab | 372 a | 4849 ab | 2049 ab | 16 ab | 99 a | 23 b | 52 a |
| Brown (n = 8) | 2938 ab | 2608 ab | 331 a | 4491 ab | 1883 ab | 17 ab | 103 a | 16 ab | 39 a |
| Orange brown (n = 2) | 3210 ab | 3014 ab | 197 a | 5255 ab | 2241 ab | 21 ab | 113 a | 9 ab | 64 a |
| n.d. (n = 12) | 3046 ab | 2759 ab | 287 a | 4684 ab | 1925 ab | 19 a | 115 ab | 9 ab | 16 a |
| Surface | |||||||||
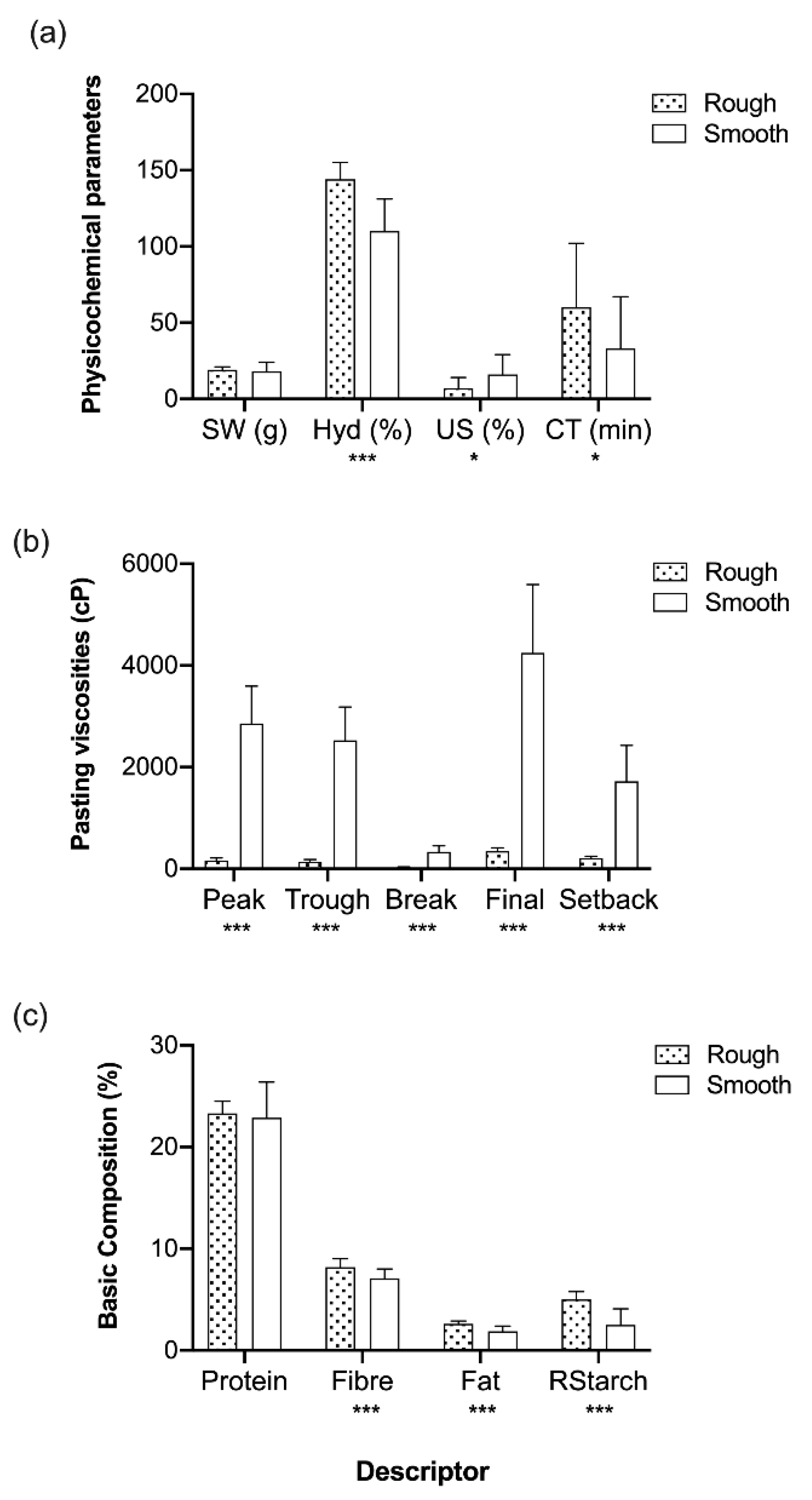
| Rough (n = 29) | 1475 a | 1302 a | 172 a | 2249 a | 947 a | 19 a | 127 a | 11 a | 47 a |
| Smooth (n = 64) | 3097 b | 2761 b | 336 b | 4694 b | 1933 b | 17 a | 110 b | 10 a | 30 ab |
| n.d. (n = 12) | 3046 b | 2759 b | 287 ab | 4684 b | 1925 b | 19 a | 115 ab | 9 a | 16 b |
| Protein (%) | Fibre (%) | Fat (%) | RS (%) | |
|---|---|---|---|---|
| Shape | ||||
| Ellipsoid (n = 42) | 22 ± 2.0 a | 7 ± 0.91 a | 2 ± 0.41 a | 3 ± 1.7 a |
| Cylindrical (n = 24) | 22 ± 1.6 a | 7 ± 0.75 a | 2 ± 0.28 a | 3 ± 1.5 a |
| Rhomboid (n = 11) | 23 ± 3.1 ab | 8 ± 0.95 a | 2 ± 0.57 a | 4 ± 1.7 a |
| Irregular (n = 16) | 24 ± 2.1 b | 8 ± 0.66 a | 2 ± 0.42 a | 5 ± 1.1 b |
| n.d. (n = 12) | 23 ± 3.4 ab | 7 ± 1.3 a | 2 ± 0.60 a | 3 ± 1.7 a |
| Colour | ||||
| Cream yellow (n = 9) | 21 ± 1.4 ac | 6 ± 0.67 a | 2 ± 0.30 a | 3 ± 0.82 ab |
| Yellow green (n = 24) | 22 ± 1.7 ab | 7 ± 0.77 a | 2 ± 0.41 a | 3 ± 1.8 ab |
| Light green (n = 14) | 22 ± 1.7 abc | 8 ± 0.89 a | 2 ± 0.61 a | 5 ± 1.5 a |
| Dark green (n = 8) | 24 ± 2.7 b | 8 ± 0.75 a | 2 ± 0.36 a | 3 ± 1.0 ab |
| Green (n = 23) | 23 ± 1.9 bc | 8 ± 1. 1 a | 2 ± 0.53 a | 4 ± 1.8 ab |
| Army green (n = 5) | 22 ± 3.3 ab | 7 ± 0.71 a | 2 ± 0.29 a | 3 ± 2.2 ab |
| Brown (n = 8) | 22 ± 3.3 ab | 7 ± 0.93 a | 2 ± 0.35 a | 3 ± 1.5 b |
| Orange-brown (n = 2) | 21 ± 0.69 ab | 7 ± 0.67 a | 2 ± 12 a | 2 ± 0.40 ab |
| n.d. (n = 12) | 23 ± 3.4 ab | 7 ± 1.3 a | 2 ± 60 a | 4 ± 1.7 ab |
| Surface | ||||
| Rough (n = 29) | 23 ± 2.5 a | 8 ± 0.90 a | 2 ± 0.52 a | 4 ± 1.7 a |
| Smooth (n = 64) | 22 ± 1.8 b | 7 ± 0.93 a | 2 ± 0.42 a | 3 ± 1.6 a |
| n.d. (n = 12) | 23 ± 3.4 a | 7 ± 1.3 a | 2 ± 0.60 a | 4 ± 1.7 a |
| Accession Number | r Allele | rb Allele | Starch Granule | DNA Assayed | Surface |
|---|---|---|---|---|---|
| 213 | r | Rb | compound | seed meal | |
| 222 | R | Rb | simple | seed meal | |
| 226 | R | Rb | simple | leaf | |
| 240 | R | Rb | simple | seed meal | |
| 241 | R | Rb | simple | seed meal | |
| 242 | R | Rb | simple | seed meal | |
| 244 | R | Rb | simple | seed meal | |
| 246 | R | Rb | simple | seed meal | |
| 247 | R | Rb | simple | leaf | |
| 256 | r | Rb | compound | seed meal | |
| 257 | r | Rb | compound | seed meal | |
| 258 | r | Rb | compound | seed meal | |
| 259 | r | Rb | compound | seed meal | |
| 261 | r | Rb | compound | seed meal | |
| 272 | r | Rb | compound | seed meal | |
| 273 | r | Rb | compound | seed meal | |
| 279 | r | Rb | compound | seed meal | |
| 282 | R | Rb | simple | seed meal | |
| 285 | r | Rb | compound | seed meal | |
| 286 | r | Rb | compound | seed meal | |
| 287 | R | Rb | simple | seed meal | |
| 288 | r | Rb | compound | seed meal | |
| 289 | r | Rb | compound | seed meal | |
| 291 | R | Rb | simple | seed meal | |
| 292 | r | Rb | compound | seed meal | |
| 293 | r | Rb | compound | seed meal | |
| 294 | r | Rb | compound | seed meal | |
| 295 | r | Rb | compound | seed meal | |
| 296 | r | Rb | compound | seed meal | |
| 297 | r | Rb | compound | seed meal | |
| 298 | R | Rb | simple | seed meal | |
| 299 | R | Rb | simple | leaf | |
| Control (JI 2822) | R | rb | simple | leaf | rough |
| Control (JI 1194) | r | Rb | compound | leaf | rough |
| Control (JI 281) | R | Rb | simple | leaf | smooth |
| Control (JI 399) | R | rb | simple | leaf | rough |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, C.S.; Carbas, B.; Castanho, A.; Vasconcelos, M.W.; Vaz Patto, M.C.; Domoney, C.; Brites, C. Variation in Pea (Pisum sativum L.) Seed Quality Traits Defined by Physicochemical Functional Properties. Foods 2019, 8, 570. https://doi.org/10.3390/foods8110570
Santos CS, Carbas B, Castanho A, Vasconcelos MW, Vaz Patto MC, Domoney C, Brites C. Variation in Pea (Pisum sativum L.) Seed Quality Traits Defined by Physicochemical Functional Properties. Foods. 2019; 8(11):570. https://doi.org/10.3390/foods8110570
Chicago/Turabian StyleSantos, Carla S., Bruna Carbas, Ana Castanho, Marta W. Vasconcelos, Maria Carlota Vaz Patto, Claire Domoney, and Carla Brites. 2019. "Variation in Pea (Pisum sativum L.) Seed Quality Traits Defined by Physicochemical Functional Properties" Foods 8, no. 11: 570. https://doi.org/10.3390/foods8110570
APA StyleSantos, C. S., Carbas, B., Castanho, A., Vasconcelos, M. W., Vaz Patto, M. C., Domoney, C., & Brites, C. (2019). Variation in Pea (Pisum sativum L.) Seed Quality Traits Defined by Physicochemical Functional Properties. Foods, 8(11), 570. https://doi.org/10.3390/foods8110570