Narrow-Banded UVB Affects the Stability of Secondary Plant Metabolites in Kale (Brassica oleracea var. sabellica) and Pea (Pisum sativum) Leaves Being Added to Lentil Flour Fortified Bread: A Novel Approach for Producing Functional Foods

 , and
, and
Abstract
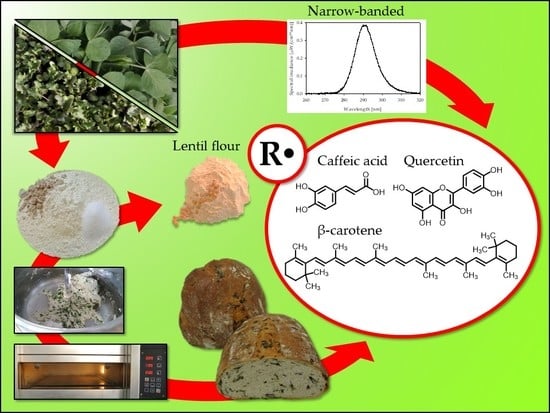
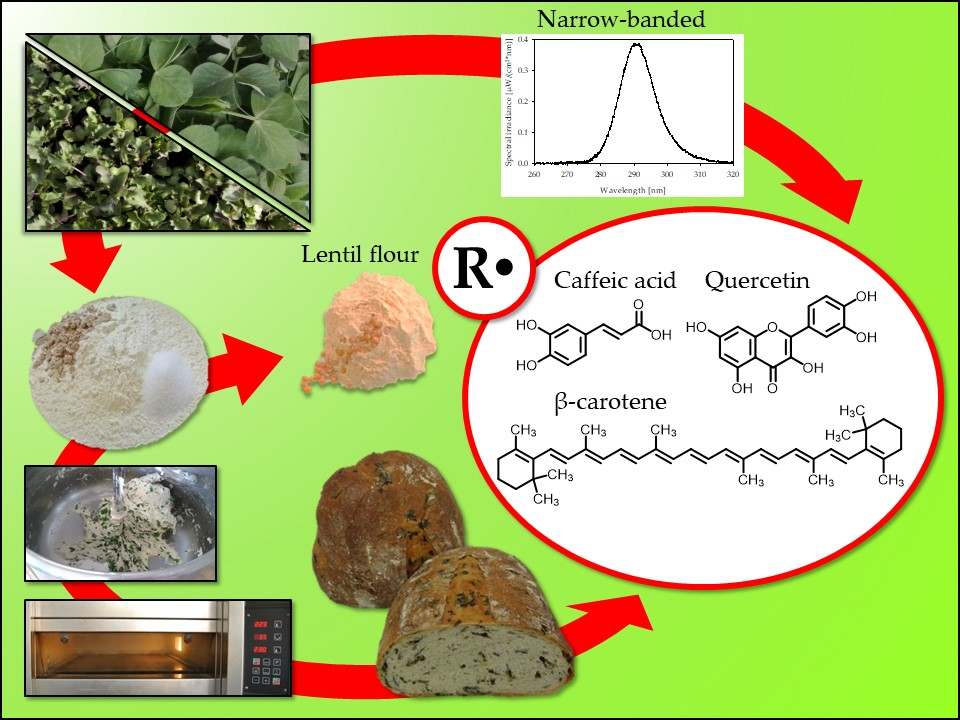
1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Plant Material
2.3. Breadmaking Experiments
2.4. Antioxidant Activity
2.4.1. Trolox Equivalent Antioxidant Activity (TEAC) Assay
2.4.2. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
2.5. Analysis of Carotenoids and Chlorophylls by Ultrahigh Perfomance Liquid Chromatography-Diode Array Detection-Time of Flight-Mass Spectrometry (UHPLC-DAD-ToF-MS)
2.6. Analysis of Flavonoid Glycosides and Hydroxycinnamic Acid Derivatives by High Perfomance Liquid Chromatography-Diode Array Detection-Electrospray Ionisation-Mass Spectrometry (HPLC-DAD-ESI-MSN)
2.7. Analysis of Protein Content by the Kjeldahl Method
2.8. Data Handling and Statistical Analysis
3. Results and Discussion
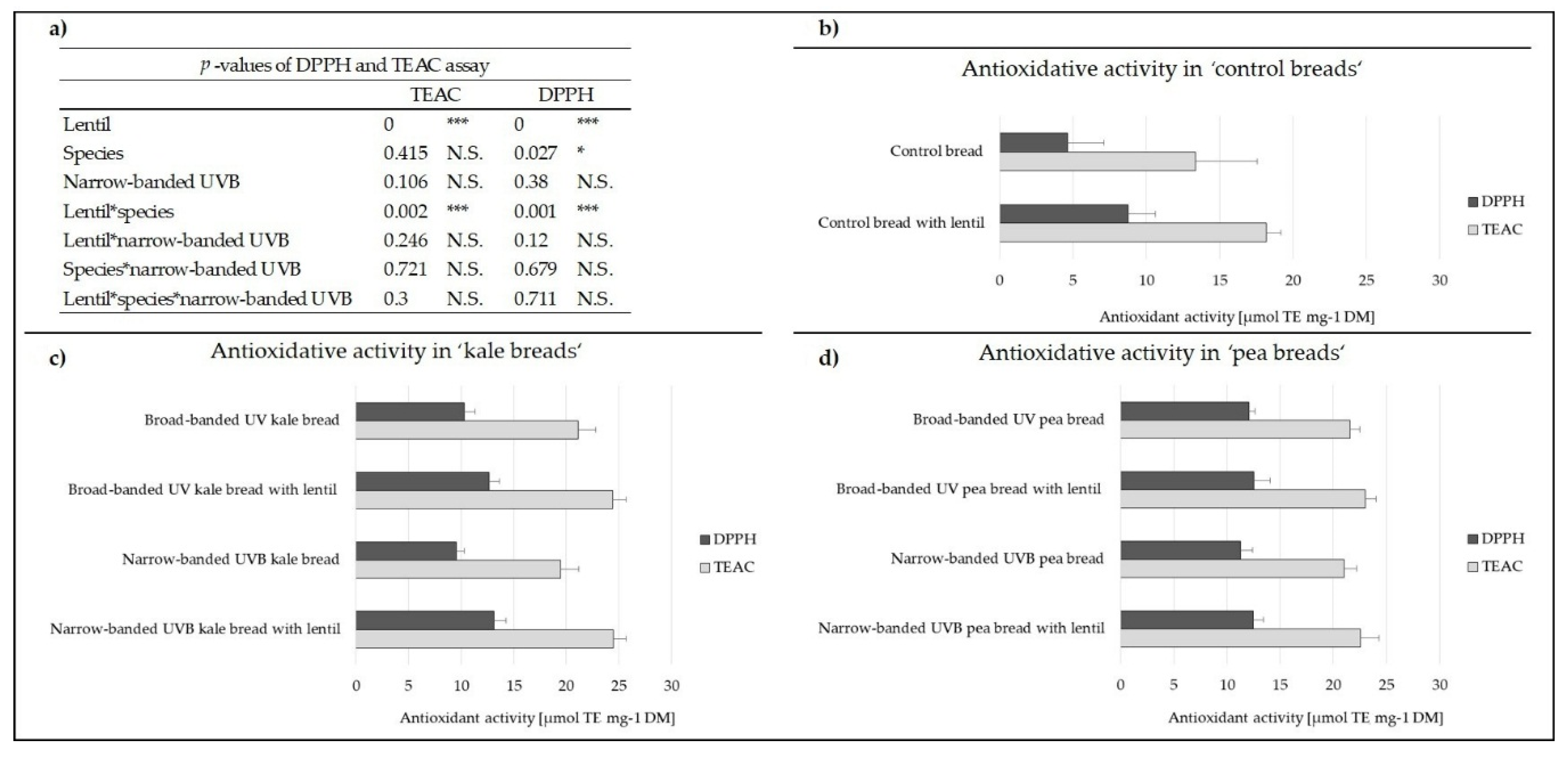
3.1. Antioxidant Activity of the Breads
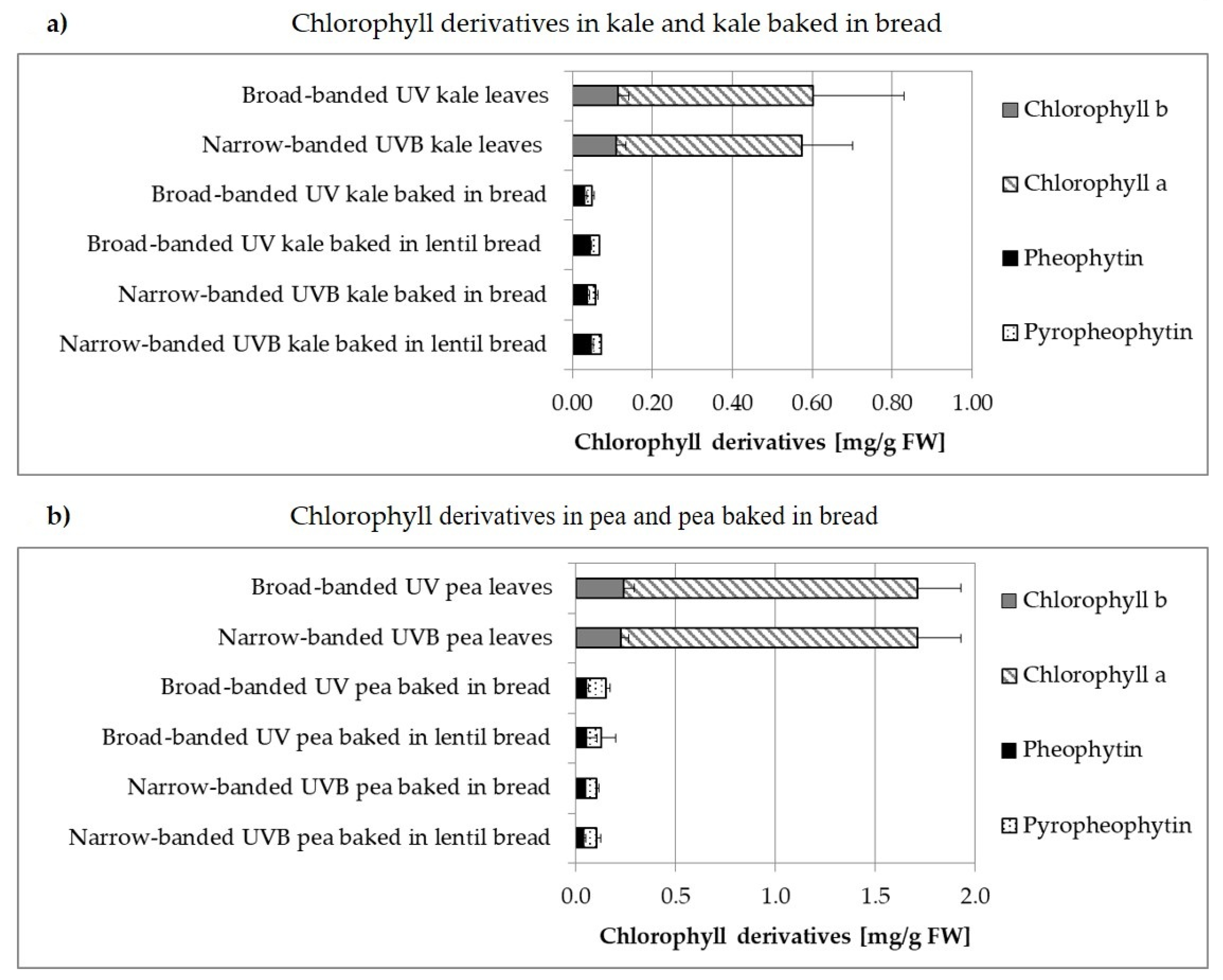
3.2. Carotenoids and Chlorophylls
3.2.1. Broad-Banded UV Light versus Narrow-Banded UVB Light
3.2.2. Non-Processed versus Processed
3.3. Flavonoid Glycosides and Hydroxycinnamic Acid Derivatives
3.3.1. Broad-Banded UV-Light versus Narrow-Banded UVB-Light
3.3.2. Non-Processed versus Processed
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martirosyan, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Food Health Dis. 2015, 5, 209–223. [Google Scholar] [CrossRef]
- Hsu, C.-Y.; Chao, P.-Y.; Hu, S.-P.; Yang, C.-M. The antioxidant and free radical scavenging activities of chlorophylls and pheophytins. Food Nutr. Sci. 2013, 4, 1. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects–A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.-W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Fung, T.T.; Hu, F.B.; Willett, W.C.; Longo, V.D.; Chan, A.T.; Giovannucci, E.L. Association of animal and plant protein intake with all-cause and cause-specific mortality. JAMA Intern. Med. 2016, 176, 1453–1463. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Polyphenol-rich lentils and their health promoting effects. Int. J. Mol. Sci. 2017, 18, 2390. [Google Scholar] [CrossRef]
- Ribas-Agusti, A.; Martin-Belloso, O.; Soliva-Fortuny, R.; Elez-Martinez, P. Food processing strategies to enhance phenolic compounds bioaccessibility and bioavailability in plant-based foods. Crit. Rev. Food Sci. Nutr. 2017, 58, 1–18. [Google Scholar] [CrossRef]
- Rohn, S. Possibilities and limitations in the analysis of covalent interactions between phenolic compounds and proteins. Food Res. Int. 2014, 65, 13–19. [Google Scholar] [CrossRef]
- Heinze, M.; Hanschen, F.S.; Wiesner-Reinhold, M.; Baldermann, S.; Gräfe, J.; Schreiner, M.; Neugart, S. Effects of Developmental Stages and Reduced UVB and Low UV Conditions on Plant Secondary Metabolite Profiles in Pak Choi (Brassica rapa subsp. chinensis). J. Agric. Food Chem. 2018, 66, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Rodríguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. UVA, UVB Light, and Methyl Jasmonate, Alone or Combined, Redirect the Biosynthesis of Glucosinolates, Phenolics, Carotenoids, and Chlorophylls in Broccoli Sprouts. Int. J. Mol. Sci. 2017, 18, 2330. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, M.; Mewis, I.; Huyskens-Keil, S.; Jansen, M.; Zrenner, R.; Winkler, J.; O’Brien, N.; Krumbein, A. UV-B-induced secondary plant metabolites-potential benefits for plant and human health. Crit. Rev. Plant Sci. 2012, 31, 229–240. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Fallik, E. Light quality manipulation improves vegetable quality at harvest and postharvest: A review. Environ. Exp. Bot. 2017, 139, 79–90. [Google Scholar] [CrossRef]
- Caldwell, M.M. Solar UV irradiation and the growth and development of higher plants. In Photophysiology; Academic Press: New York, NY, USA; London, UK, 1971; Volume 6, pp. 131–177. [Google Scholar]
- Neugart, S.; Schreiner, M. UVB and UVA as eustressors in horticultural and agricultural crops. Sci. Hortic. 2018, 234, 370–381. [Google Scholar] [CrossRef]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sust. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef]
- Ahmed, M.; Eun, J.-B. Flavonoids in fruits and vegetables after thermal and nonthermal processing: A review. Crit. Rev. Food Sci. Nutr. 2017, 58, 1–30. [Google Scholar] [CrossRef]
- Turkmen, N.; Poyrazoglu, E.S.; Sari, F.; Sedat Velioglu, Y. Effects of cooking methods on chlorophylls, pheophytins and colour of selected green vegetables. Int. J. Food Sci. Technol. 2006, 41, 281–288. [Google Scholar] [CrossRef]
- Klopsch, R.; Baldermann, S.; Hanschen, F.S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Brassica-enriched wheat bread: Unraveling the impact of ontogeny and breadmaking on bioactive secondary plant metabolites of pak choi and kale. Food Chem. 2019, 295, 412–422. [Google Scholar] [CrossRef]
- Klopsch, R.; Baldermann, S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Bread enriched with legume microgreens and leaves–ontogenetic and baking-driven changes in the profile of secondary plant metabolites. Front. Chem. 2018, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Buscemi, S.; Corleo, D.; Di Pace, F.; Petroni, M.; Satriano, A.; Marchesini, G. The Effect of Lutein on Eye and Extra-Eye Health. Nutrients 2018, 10, 1321. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Identification of complex, naturally occurring flavonoid glycosides in kale (Brassica oleracea var. sabellica) by high-performance liquid chromatography diode-array detection/electrospray ionization multi-stage mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2009–2022. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Rohn, S.; Schreiner, M. Identification of complex, naturally occurring flavonoid glycosides in Vicia faba and Pisum sativum leaves by HPLC-DAD-ESI-MS n and the genotypic effect on their flavonoid profile. Food Res. Int. 2015, 76, 114–121. [Google Scholar] [CrossRef]
- Csepregi, K.; Neugart, S.; Schreiner, M.; Hideg, É. Comparative evaluation of total antioxidant capacities of plant polyphenols. Molecules 2016, 21, 208. [Google Scholar] [CrossRef] [PubMed]
- Fiol, M.; Weckmüller, A.; Neugart, S.; Schreiner, M.; Rohn, S.; Krumbein, A.; Kroh, L.W. Thermal-induced changes of kale’s antioxidant activity analyzed by HPLC–UV/Vis-online-TEAC detection. Food Chem. 2013, 138, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Helrich, K., Ed.; Association of Analytical Chemists: Washington, DC, USA, 1990; Volume 15, p. 771. [Google Scholar]
- Llorente, B.; Martinez-Garcia, J.F.; Stange, C.; Rodriguez-Concepcion, M. Illuminating colors: Regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49–55. [Google Scholar] [CrossRef]
- Reif, C.; Arrigoni, E.; Berger, F.; Baumgartner, D.; Nyström, L. Lutein and β-carotene content of green leafy Brassica species grown under different conditions. LWT Food Sci. Technol. 2013, 53, 378–381. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Kopsell, D.E. Accumulation and bioavailability of dietary carotenoids in vegetable crops. Trends Plant Sci. 2006, 11, 499–507. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W. Carotenoid composition in sun and shade leaves of plants with different life forms. Plant Cell Environ. 1992, 15, 411–419. [Google Scholar] [CrossRef]
- Mewis, I.; Schreiner, M.; Nguyen, C.N.; Krumbein, A.; Ulrichs, C.; Lohse, M.; Zrenner, R. UV-B irradiation changes specifically the secondary metabolite profile in broccoli sprouts: Induced signaling overlaps with defense response to biotic stressors. Plant Cell Physiol. 2012, 53, 1546–1560. [Google Scholar] [CrossRef] [PubMed]
- Becatti, E.; Petroni, K.; Giuntini, D.; Castagna, A.; Calvenzani, V.; Serra, G.; Mensuali-Sodi, A.; Tonelli, C.; Ranieri, A. Solar UV-B radiation influences carotenoid accumulation of tomato fruit through both ethylene-dependent and-independent mechanisms. J. Agric. Food Chem. 2009, 57, 10979–10989. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, V.; Calvenzani, V.; Petroni, K.; Tonelli, C.; Castagna, A.; Ranieri, A. Carotenoid profiling and biosynthetic gene expression in flesh and peel of wild-type and hp-1 tomato fruit under UV-B depletion. J. Agric. Food Chem. 2012, 60, 4960–4969. [Google Scholar] [CrossRef] [PubMed]
- Canjura, F.L.; Schwartz, S.J.; Nunes, R.V. Degradation kinetics of chlorophylls and chlorophyllides. J. Food Sci. 1991, 56, 1639–1643. [Google Scholar] [CrossRef]
- Teng, S.; Chen, B. Formation of pyrochlorophylls and their derivatives in spinach leaves during heating. Food Chem. 1999, 65, 367–373. [Google Scholar] [CrossRef]
- Schwartz, S.; Von Elbe, J. Kinetics of chlorophyll degradation to pyropheophytin in vegetables. J. Food Sci. 1983, 48, 1303–1306. [Google Scholar] [CrossRef]
- Onkokesung, N.; Reichelt, M.; van Doorn, A.; Schuurink, R.C.; van Loon, J.J.; Dicke, M. Modulation of flavonoid metabolites in Arabidopsis thaliana through overexpression of the MYB75 transcription factor: Role of kaempferol-3, 7-dirhamnoside in resistance to the specialist insect herbivore Pieris brassicae. J. Exp. Bot. 2014, 65, 2203–2217. [Google Scholar] [CrossRef]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant. Sci. 2016, 7, 735. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Said, A.M.; Khalifa, S.A.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.-K.; Verpoorte, R. Biosynthesis, natural sources, dietary intake, pharmacokinetic properties, and biological activities of hydroxycinnamic acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef]
- Majer, P.; Neugart, S.; Krumbein, A.; Schreiner, M.; Hideg, É. Singlet oxygen scavenging by leaf flavonoids contributes to sunlight acclimation in Tilia platyphyllos. Environ. Exp. Bot. 2014, 100, 1–9. [Google Scholar] [CrossRef]
- Neugart, S.; Fiol, M.; Schreiner, M.; Rohn, S.; Zrenner, R.; Kroh, L.W.; Krumbein, A. Interaction of moderate UV-B exposure and temperature on the formation of structurally different flavonol glycosides and hydroxycinnamic acid derivatives in kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2014, 62, 4054–4062. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Rechner, O.; Neugart, S.; Schreiner, M.; Wu, S.; Poehling, H.-M. Different narrow-band light ranges alter plant secondary metabolism and plant defense response to aphids. J. Chem. Ecol. 2016, 42, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.; Brunetti, C.; Fini, A.; Agati, G.; Ferrini, F.; Gori, A.; Tattini, M. UV radiation promotes flavonoid biosynthesis, while negatively affecting the biosynthesis and the de-epoxidation of xanthophylls: Consequence for photoprotection? Environ. Exp. Bot. 2016, 127, 14–25. [Google Scholar] [CrossRef]
- Chaaban, H.; Ioannou, I.; Chebil, L.; Slimane, M.; Gérardin, C.; Paris, C.; Charbonnel, C.; Chekir, L.; Ghoul, M. Effect of heat processing on thermal stability and antioxidant activity of six flavonoids. J. Food Process. Preserv. 2017, 41, e13203. [Google Scholar] [CrossRef]
- Rohn, S.; Buchner, N.; Driemel, G.; Rauser, M.; Kroh, L.W. Thermal degradation of onion quercetin glucosides under roasting conditions. J. Agric. Food Chem. 2007, 55, 1568–1573. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| (a) | Kale Leaves | Kale Bread | Kale Bread with Lentil | |||
|---|---|---|---|---|---|---|
| mg/g FW in Plant Tissue | Recovery in Kale Bread (%) | |||||
| Lutein | broad-banded UV | 0.034 A | ± | 0.009 | 1.8 a | 2 a |
| narrow-banded UVB | 0.028 A | ± | 0.008 | 1.9 a | 2.1 a | |
| α-Carotene | broad-banded UV | 0.001 A | ± | 0.001 | N.D. | N.D. |
| narrow-banded UVB | 0.002 B | ± | 0.001 | |||
| ß-Carotene | broad-banded UV | 0.011 A | ± | 0.004 | N.D. | N.D. |
| narrow-banded UVB | 0.009 A | ± | 0.003 | |||
| Neoxanthin derivative 1 | broad-banded UV | 0.01 A | ± | 0.003 | N.D. | N.D. |
| narrow-banded UVB | 0.009 A | ± | 0.003 | |||
| Neoxanthin derivative 2 | broad-banded UV | 0.004 A | ± | 0.001 | N.D. | N.D. |
| narrow-banded UVB | 0.005 B | ± | 0.001 | |||
| Violaxanthin | broad-banded UV | 0.001 A | ± | 0.001 | N.D. | N.D. |
| narrow-banded UVB | 0.001 A | ± | 0.001 | |||
| Total carotenoids | broad-banded UV | 0.06 A | ± | 0.019 | 1.2 a | 1.36 a |
| narrow-banded UVB | 0.055 A | ± | 0.017 | 1 a | 1.1 a | |
| (b) | Pea Leaves | Pea Bread | Pea Bread with Lentil | |||
| mg/g FW in Plant Tissue | Recovery in Pea Bread (%) | |||||
| Lutein | broad-banded UV | 0.102 A | ± | 0.023 | 15.1 a | 11 a |
| narrow-banded UVB | 0.106 A | ± | 0.020 | 9 b | 11.3 b | |
| ß-Carotene | broad-banded UV | 0.052 A | ± | 0.008 | N.D. | N.D. |
| narrow-banded UVB | 0.052 A | ± | 0.009 | |||
| Neoxanthin derivative 1 | broad-banded UV | 0.023 A | ± | 0.006 | N.D. | N.D. |
| narrow-banded UVB | 0.021 A | ± | 0.005 | |||
| Neoxanthin derivative 2 | broad-banded UV | 0.003 A | ± | 0.001 | N.D. | N.D. |
| narrow-banded UVB | 0.003 A | ± | 0.001 | |||
| Violaxanthin derivative 1 | broad-banded UV | 0.037 A | ± | 0.015 | N.D. | N.D. |
| narrow-banded UVB | 0.038 A | ± | 0.012 | |||
| Violaxanthin derivative 2 | broad-banded UV | 0.005 A | ± | 0.001 | N.D. | N.D. |
| narrow-banded UVB | 0.006 A | ± | 0.001 | |||
| Total carotenoids | broad-banded UV | 0.223 A | ± | 0.053 | 7 a | 6.9 a |
| narrow-banded UVB | 0.226 A | ± | 0.047 | 4.2 a | 5.3 b | |
| (a) Hydroxycinnamic Acid Derivatives | Kale | Kale Bread | Kale Bread with Lentil | |||
|---|---|---|---|---|---|---|
| mg/g FW in Plant Tissue | Recovery in Kale Bread (%) | |||||
| 3-Caffeoylquinic acid | broad-banded UV | 0.309 A | ± | 0.065 | 14 a | 14 a |
| narrow-banded UVB | 0.259 A | ± | 0.044 | 15.7 b | 14.5 b | |
| Caffeoylglucoside | broad-banded UV | 0.078 A | ± | 0.020 | N.D. | N.D. |
| narrow-banded UVB | 0.072 A | ± | 0.009 | |||
| Sinapic acid-glucoside | broad-banded UV | 0.185 A | ± | 0.037 | 21 a | 21.2 a |
| narrow-banded UVB | 0.239 B | ± | 0.026 | 15.7 b | 16.8 b | |
| Sinapoyl-feruloylgentiobiose | broad-banded UV | 0.257 A | ± | 0.049 | 15.1 a | 15.7 a |
| narrow-banded UVB | 0.204 B | ± | 0.034 | 18.3 b | 19 b | |
| Disinapoyl-feruloylgentiobiose | broad-banded UV | 0.095 A | ± | 0.018 | 33.5 a | 33.8 a |
| narrow-banded UVB | 0.092 A | ± | 0.012 | 34.6 b | 34.7 a | |
| Sinapoyl-hydroxyferuloylgentiobiose | broad-banded UV | 0.089 A | ± | 0.018 | 38.5 a | 38.5 a |
| narrow-banded UVB | 0.082 A | ± | 0.015 | 39.8 b | 40.5 b | |
| Disinapoyl-gentiobiose | broad-banded UV | 0.268 A | ± | 0.051 | 17.2 a | 17.8 a |
| narrow-banded UVB | 0.263 A | ± | 0.038 | 17.7 a | 19.4 a | |
| Trisinapoyl-gentiobiose | broad-banded UV | 0.184 A | ± | 0.045 | 26 a | 25.6 a |
| narrow-banded UVB | 0.203 A | ± | 0.032 | 23.6 b | 26.2 a | |
| Hydroxycinnamic acid derivatives | broad-banded UV | 1.465 A | ± | 0.302 | 19.2 a | 19.4 a |
| narrow-banded UVB | 1.413 A | ± | 0.211 | 19.4 a | 20.2 a | |
| (b) Kaempferol Glycosides | Kale | Kale Bread | Kale Bread with Lentil | |||
| mg/g FW in Plant Tissue | Recovery in Kale Bread (%) | |||||
| K-3-O-dirha-7-O-rha | broad-banded UV | 0.153 A | ± | 0.024 | 39.9 a | 37.6 a |
| narrow-banded UVB | 0.128 B | ± | 0.026 | 48.8 b | 47.5 b | |
| K-3-O-soph-7-O-ᴅ-glc | broad-banded UV | 0.163 A | ± | 0.052 | 10.1 a | 10.7 a |
| narrow-banded UVB | 0.124 A | ± | 0.040 | 6.1 b | 7 b | |
| K-3-O-cou-soph-7-O-ᴅ-glc | broad-banded UV | 0.08 A | ± | 0.023 | 19.1 a | 20.2 a |
| narrow-banded UVB | 0.035 B | ± | 0.011 | 33.6 b | 31.8 b | |
| K-3-O-caf-soph-7-O-ᴅ-glc | broad-banded UV | 0.529 A | ± | 0.141 | 17.1 a | 16.8 a |
| narrow-banded UVB | 0.388 B | ± | 0.129 | 17.3 a | 17.1 a | |
| K-3-O-fer-soph-7-O-glc | broad-banded UV | 0.17 A | ± | 0.045 | 16.4 a | 17.9 a |
| narrow-banded UVB | 0.1 B | ± | 0.031 | 20.4 b | 21.7 b | |
| K-3-O-hfer-soph-7-O-glc | broad-banded UV | 1.204 A | ± | 0.302 | 11.4 a | 11.8 a |
| narrow-banded UVB | 0.936 B | ± | 0.192 | 9.2 b | 9.5 b | |
| K-3-O-sin-soph-7-O-glc | broad-banded UV | 0.26 A | ± | 0.091 | 19.9 a | 21.3 a |
| narrow-banded UVB | 0.235 A | ± | 0.076 | 13.6 b | 15.4 b | |
| Kaempferol glycosides | broad-banded UV | 2.56 A | ± | 0.678 | 15.6 a | 16 a |
| narrow-banded UVB | 1.945 B | ± | 0.504 | 14.8 a | 15.1 a | |
| (c) Quercetin glycosides | Kale | Kale Bread | Kale Bread with Lentil | |||
| mg/g FW in Plant Tissue | Recovery in Kale Bread (%) | |||||
| Q-3-O-triglc | broad-banded UV | 0.495 A | ± | 0.69 | 10.3 a | 10.2 a |
| narrow-banded UVB | 0.624 B | ± | 0.109 | 7.2 b | 8.5 b | |
| Q-3,7,4´-O-ᴅ-triglc | broad-banded UV | 0.145 A | ± | 0.034 | 23.6 a | 25.5 a |
| narrow-banded UVB | 0.132 A | ± | 0.040 | 23.1 a | 22.6 b | |
| Q-3-O-caf-soph-7-O-glc | broad-banded UV | 0.15 A | ± | 0.047 | 40.2 a | 35.1 a |
| narrow-banded UVB | 0.195 B | ± | 0.047 | 15.5 b | 16.1 b | |
| Q-3-O-fer-soph-7-O-ᴅ-glc | broad-banded UV | 0.094 A | ± | 0.024 | N.D. | N.D. |
| narrow-banded UVB | 0.109 A | ± | 0.036 | |||
| Q-3-O-hfer-sophtr-7-O-glc | broad-banded UV | 0.188 A | ± | 0.040 | 75.6 a | 77.2 a |
| narrow-banded UVB | 0.176 B | ± | 0.041 | 63.1 b | 72 a | |
| Q-3-O-sin-soph-7-O-ᴅ-glc | broad-banded UV | 0.131 A | ± | 0.044 | 38.2 a | 38.1 a |
| narrow-banded UVB | 0.175 B | ± | 0.091 | 18.7 b | 21.4 b | |
| Quercetin glycosides | broad-banded UV | 1.203 A | ± | 0.305 | 28.1 a | 27.9 a |
| narrow-banded UVB | 1.41 A | ± | 0.363 | 17.7 a | 19.7 b | |
| (a) Hydroxycinnamic Acid Glycosides | Pea | Pea Bread | Pea Bread with Lentil | ||||||
|---|---|---|---|---|---|---|---|---|---|
| mg/g FW in Plant Tissue | Recovery in Pea Bread (%) | ||||||||
| Coumaroyl-glucoside | broad-banded UV | 0.022 A | ± | 0.008 | 63.5 a | 84.5 a | |||
| narrow-banded UVB | 0.016 B | ± | 0.004 | 47.45 b | 79.1 a | ||||
| Caffeoyl-glucoside 1 | broad-banded UV | 0.006 A | ± | 0.001 | N.D. | N.D. | |||
| narrow-banded UVB | 0.006 A | ± | 0.001 | ||||||
| Caffeoyl-glucoside 2 | broad-banded UV | 0.007 A | ± | 0.001 | N.D. | N.D. | |||
| narrow-banded UVB | 0.007 A | ± | 0.002 | ||||||
| Caffeoyl-glucoside 3 | broad-banded UV | 0.006 A | ± | 0.001 | N.D. | N.D. | |||
| narrow-banded UVB | 0.005 A | ± | 0.001 | ||||||
| Hydroxycinnamic acid derivatives | broad-banded UV | 0.041 A | ± | 0.012 | 34.3 a | 39.7 a | |||
| narrow-banded UVB | 0.034 A | ± | 0.005 | 25.7 a | 37.2 b | ||||
| (b) Kaempferol Glycosides | Pea | Pea Bread | Pea Bread with Lentil | ||||||
| mg/g FW in Plant Tissue | Recovery in Pea Bread (%) | ||||||||
| K-3-O-sophtr 1 | broad-banded UV | 0.086 A | ± | 0.013 | 111 a | 94.4 a | |||
| narrow-banded UVB | 0.098 B | ± | 0.01 | 109.7 a | 107.6 b | ||||
| K-3-O-sophtr 2 | broad-banded UV | 0.024 A | ± | 0.014 | N.D. | N.D. | |||
| narrow-banded UVB | 0.011 B | ± | 0.005 | ||||||
| K-3-O-cou-sophtr 1 | broad-banded UV | 0.681 A | ± | 0.108 | 92.4 a | 77.5 a | |||
| narrow-banded UVB | 0.742 A | ± | 0.114 | 82.1 a | 85.5 b | ||||
| K-3-O-cou-sophtr 2 | broad-banded UV | 0.043 A | ± | 0.008 | 61.5 a | 66.5 a | |||
| narrow-banded UVB | 0.039 A | ± | 0.004 | 52.3 a | 71.2 a | ||||
| K-3-O-sin-sophtr | broad-banded UV | 0.089 A | ± | 0.019 | 99.9 a | 89.4 a | |||
| narrow-banded UVB | 0.093 A | ± | 0.013 | 86.8 b | 89.2 a | ||||
| Kaempferol glycosides | broad-banded UV | 0.922 A | ± | 0.128 | 91.1 a | 79 a | |||
| narrow-banded UVB | 0.984 A | ± | 0.13 | 81.6 b | 86.5 a | ||||
| (c) Quercetin Glycosides | Pea | Pea Bread | Pea Bread with Lentil | ||||||
| mg/g FW in Plant Tissue | Recovery in Pea Bread (%) | ||||||||
| Q-3-O-sophtr 1 | broad-banded UV | 0.354 A | ± | 0.061 | 111.1 a | 93.8 a | |||
| narrow-banded UVB | 0.409 B | ± | 0.02 | 106.6 a | 112 a | ||||
| Q-3-O-sophtr 2 | broad-banded UV | 0.065 A | ± | 0.028 | 178.9 a | 163.1 a | |||
| narrow-banded UVB | 0.07 A | ± | 0.018 | 168.5 a | 187.6 b | ||||
| Q-3-O-cou-sophtr 1 | broad-banded UV | 3.007 A | ± | 0.28 | 100.3 a | 92.4 a | |||
| narrow-banded UVB | 2.896 A | ± | 0.208 | 75.9 b | 91.9 a | ||||
| Q-3-O-cou-sophtr 2 | broad-banded UV | 0.114 A | ± | 0.016 | 109.6 a | 91.6 a | |||
| narrow-banded UVB | 0.122 A | ± | 0.041 | 99.1 a | 103.6 a | ||||
| Q-3-O-cou-sophtr 3 | broad-banded UV | 0.169 A | ± | 0.045 | 68.9 a | 78.9 a | |||
| narrow-banded UVB | 0.137 A | ± | 0.019 | 50.1 b | 72.3 a | ||||
| Q-3-O-caf-sophtr | broad-banded UV | 0.063 A | ± | 0.017 | 108.5 a | 89 a | |||
| narrow-banded UVB | 0.065 A | ± | 0.011 | 96.1 b | 99.7 a | ||||
| Quercetin glycosides | broad-banded UV | 3.772 A | ± | 0.362 | 101.7 a | 93.3 a | |||
| narrow-banded UVB | 3.697 A | ± | 0.261 | 80.3 a | 95.8 b | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klopsch, R.; Baldermann, S.; Voss, A.; Rohn, S.; Schreiner, M.; Neugart, S. Narrow-Banded UVB Affects the Stability of Secondary Plant Metabolites in Kale (Brassica oleracea var. sabellica) and Pea (Pisum sativum) Leaves Being Added to Lentil Flour Fortified Bread: A Novel Approach for Producing Functional Foods. Foods 2019, 8, 427. https://doi.org/10.3390/foods8100427
Klopsch R, Baldermann S, Voss A, Rohn S, Schreiner M, Neugart S. Narrow-Banded UVB Affects the Stability of Secondary Plant Metabolites in Kale (Brassica oleracea var. sabellica) and Pea (Pisum sativum) Leaves Being Added to Lentil Flour Fortified Bread: A Novel Approach for Producing Functional Foods. Foods. 2019; 8(10):427. https://doi.org/10.3390/foods8100427
Chicago/Turabian StyleKlopsch, Rebecca, Susanne Baldermann, Alexander Voss, Sascha Rohn, Monika Schreiner, and Susanne Neugart. 2019. "Narrow-Banded UVB Affects the Stability of Secondary Plant Metabolites in Kale (Brassica oleracea var. sabellica) and Pea (Pisum sativum) Leaves Being Added to Lentil Flour Fortified Bread: A Novel Approach for Producing Functional Foods" Foods 8, no. 10: 427. https://doi.org/10.3390/foods8100427
APA StyleKlopsch, R., Baldermann, S., Voss, A., Rohn, S., Schreiner, M., & Neugart, S. (2019). Narrow-Banded UVB Affects the Stability of Secondary Plant Metabolites in Kale (Brassica oleracea var. sabellica) and Pea (Pisum sativum) Leaves Being Added to Lentil Flour Fortified Bread: A Novel Approach for Producing Functional Foods. Foods, 8(10), 427. https://doi.org/10.3390/foods8100427