
Advanced Modification Strategies of Plant-Sourced Dietary Fibers and Their Applications in Functional Foods




Abstract
1. Introduction
2. Sources and Characteristics of PDF
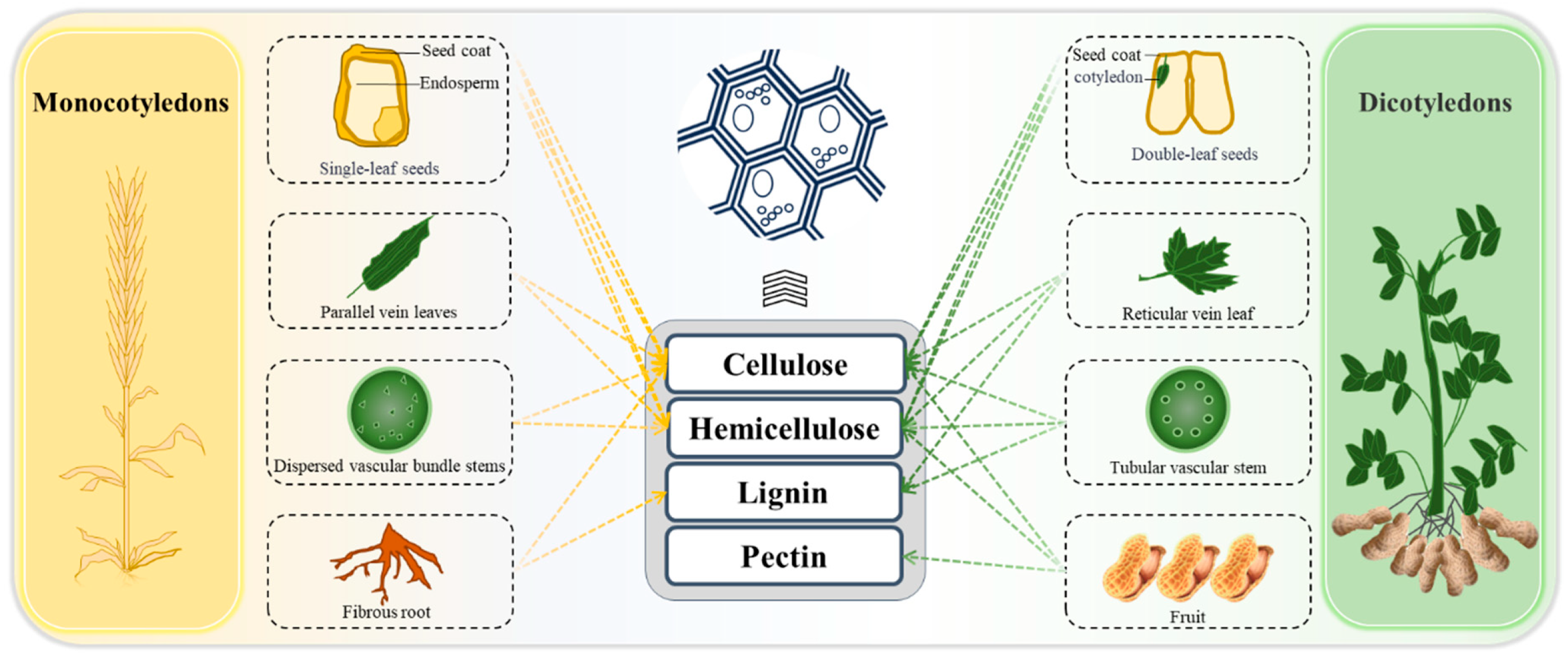
2.1. Tissue-Specific Distribution of Dietary Fiber in Plants
2.2. Physicochemical Properties of PDF
3. Innovative Modification Techniques for PDF
3.1. Physical Modification
3.2. Sustainable Chemical Modification
3.3. Bioengineering Modification
3.4. Hybrid Modification
3.5. Comparative Analysis of Modification Methods
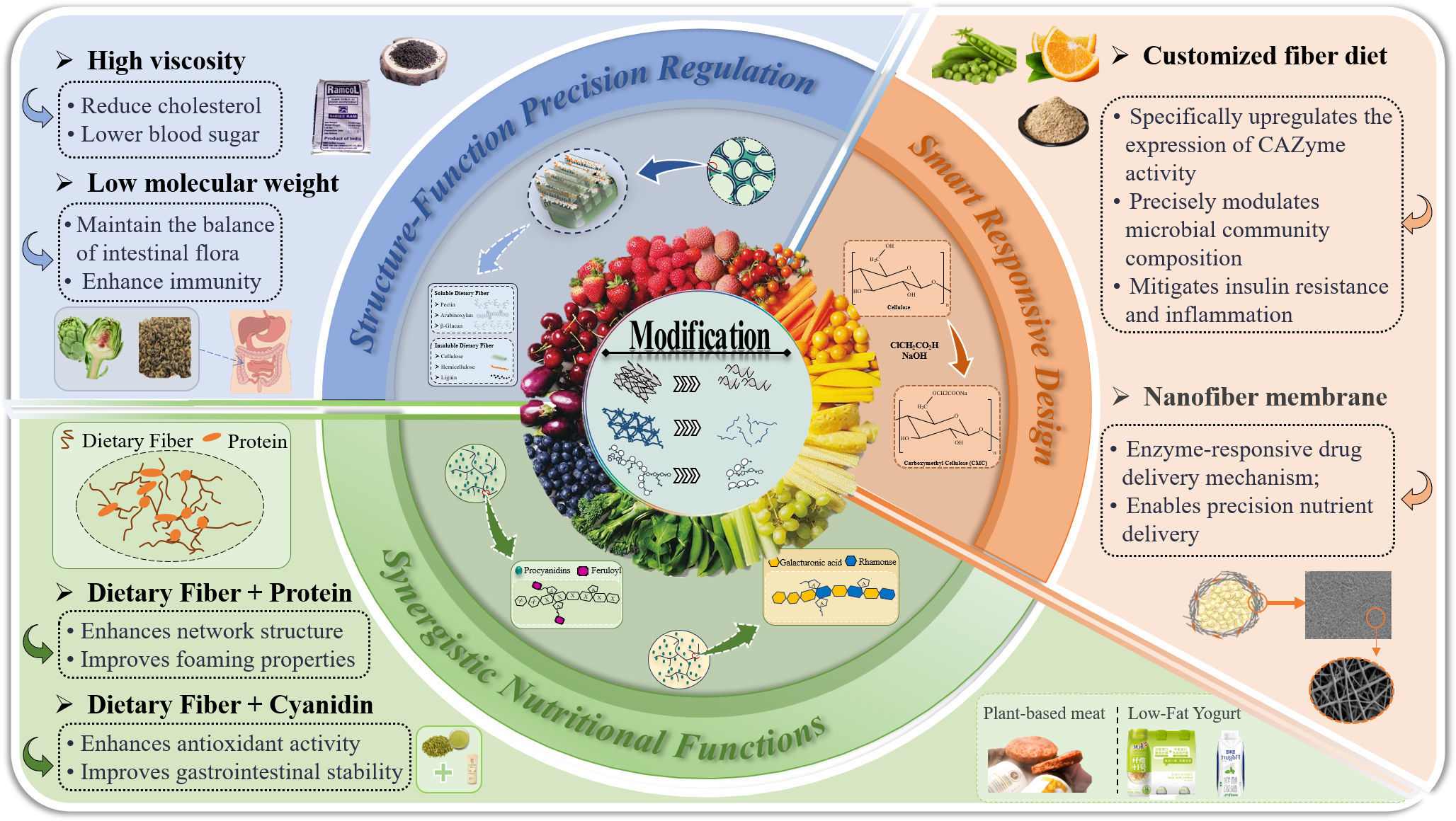
4. Mechanisms of Functional Improvement in Modified PDF
4.1. Structure–Function Precision Regulation
4.1.1. Modified Plant-Sourced Dietary Fiber Viscosity
4.1.2. Modified Plant-Sourced Dietary Fibers with Hierarchical Porosity
4.2. Synergistic Nutritional Functions
4.2.1. Polyphenol-Bound Plant-Sourced Dietary Fiber Complexes
4.2.2. Protein-Bound Modified Plant-Sourced Dietary Fiber Complexes
4.3. Smart Responsive Design
4.3.1. Smart pH-Sensitive Modification of Plant-Sourced Dietary Fibers
4.3.2. Combinatorial Customization of Modified Dietary Fiber for Personalized Gut-Targeted Delivery
5. Main Approaches for Functional Food Applications of PDF
5.1. Engineered Fiber Formulation Design
5.1.1. Low-Calorie Functional Foods
5.1.2. Microbiota-Targeting Functional Foods
5.2. Innovative Product Applications
5.2.1. 3D-Printed Personalized Nutrition Foods
5.2.2. Low-Glycemic-Index (GI) Energy Bar
5.3. Sensory Properties and Consumer Acceptance
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Plant-sourced Dietary Fiber |
References
- Fuller, S.; Beck, E.; Salman, H.; Tapsell, L. New Horizons for the Study of Dietary Fiber and Health: A Review. Plant Foods Hum. Nutr. 2016, 71, 1–12. [Google Scholar] [CrossRef]
- Chen, Z.; Mense, A.L.; Brewer, L.R.; Shi, Y.-C. Wheat bran layers: Composition, structure, fractionation, and potential uses in foods. Crit. Food Sci. 2024, 64, 6636–6659. [Google Scholar] [CrossRef]
- Chen, Z.; Mense, A.L.; Brewer, L.R.; Shi, Y.-C. Wheat bran arabinoxylans: Chemical structure, extraction, properties, health benefits, and uses in foods. Crit. Rev. Food Sci. Nutr. 2024, 23, 3. [Google Scholar] [CrossRef]
- Cyran, M.R.; Saulnier, L. Cell wall fractions isolated from outer layers of rye grain by sequential treatment with α-amylase and proteinase: Structural investigation of polymers in two ryes with contrasting breadmaking quality. J. Agric. Food Chem. 2005, 53, 9213–9224. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Ryden, P.; Edwards, C.H.; Grundy, M.M.L. Plant Cell Walls: Impact on Nutrient Bioaccessibility and Digestibility. Foods 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, E.; Tsopmo, A.; Oliviero, T.; Fogliano, V.; Udenigwe, C.C. Bioprocessing of common pulses changed seed microstructures, and improved dipeptidyl peptidase-IV and α-glucosidase inhibitory activities. Sci. Rep. 2019, 9, 15308. [Google Scholar] [CrossRef]
- Hemery, Y.M.; Anson, N.M.; Havenaar, R.; Haenen, G.R.M.M.; Noort, M.W.J.; Rouau, X. Dry-fractionation of wheat bran increases the bioaccessibility of phenolic acids in breads made from processed bran fractions. Food Res. Int. 2010, 43, 1429–1438. [Google Scholar] [CrossRef]
- Ma, C.; Bai, J.; Shao, C.; Liu, J.; Zhang, Y.; Li, X.; Yang, Y.; Xu, Y.; Wang, L. Degradation of blue honeysuckle polysaccharides, structural characteristics and antiglycation and hypoglycemic activities of degraded products. Food Res. Int. 2021, 143, 110281. [Google Scholar] [CrossRef]
- Arayici, M.E.; Mert-Ozupek, N.; Yalcin, F.; Basbinar, Y.; Ellidokuz, H. Soluble and Insoluble Dietary Fiber Consumption and Colorectal Cancer Risk: A Systematic Review and Meta-Analysis. Nutr. Cancer 2022, 74, 2412–2425. [Google Scholar] [CrossRef]
- Giuntini, E.B.; Sardá, F.A.H.; de Menezes, E.W. The Effects of Soluble Dietary Fibers on Glycemic Response: An Overview and Futures Perspectives. Foods 2022, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Moturi, J.; Hosseindoust, A.; Tajudeen, H.; Mun, J.Y.; Ha, S.H.; Kim, J.S. Influence of dietary fiber intake and soluble to insoluble fiber ratio on reproductive performance of sows during late gestation under hot climatic conditions. Sci. Rep. 2022, 12, 19749. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.R.; Zhang, W.X.; Cheng, J.; Lu, Z.B. Antioxidant and physicochemical properties of soluble dietary fiber from garlic straw as treated by energy-gathered ultrasound. Int. J. Food Prop. 2019, 22, 678–688. [Google Scholar] [CrossRef]
- Tan, W.S.K.; Chia, P.F.W.; Ponnalagu, S.; Karnik, K.; Henry, C.J. The Role of Soluble Corn Fiber on Glycemic and Insulin Response. Nutrients 2020, 12, 4. [Google Scholar] [CrossRef]
- Ul Ain, H.B.; Saeed, F.; Ahmed, A.; Khan, M.A.; Niaz, B.; Tufail, T. Improving the physicochemical properties of partially enhanced soluble dietary fiber through innovative techniques: A coherent review. J. Food Process. Preserv. 2019, 43, 4. [Google Scholar] [CrossRef]
- Tufail, T.; Saeed, F.; Tufail, T.; Ul Ain, H.B.; Hussain, M.; Noreen, S.; Shah, M.A. Exploring the cholesterol-lowering effects of cereal bran cell wall-enriched diets. Food Sci. Nutr. 2024, 12, 4944–4951. [Google Scholar] [CrossRef]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Saeed, F.; Javed, M.; Afzaal, M.; Niaz, B.; Imran, A.; Naz, A.; Umar, M. Extraction and characterization of cereal bran cell wall in relation to its end use perspectives. J. Food Sci. Technol. 2022, 59, 4615–4623. [Google Scholar] [CrossRef]
- de Almeida Costa, G.E.; da Silva Queiroz-Monici, K.; Pissini Machado Reis, S.M.; de Oliveira, A.C. Chemical composition, dietary fibre and resistant starch contents of raw and cooked pea, common bean, chickpea and lentil legumes. Food Chem. 2006, 94, 327–330. [Google Scholar] [CrossRef]
- Bajić, A.; Cvetković, B.; Mastilović, J.; Hadnađev, M.; Djordjević, M.; Djordjević, M.; Filipčev, B. Implementation of Plum Skin as a Structuring Agent in Plum Spread. Foods 2025, 14, 697. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Cheikhrouhou, S.; Renard, C.M.G.C.; Bureau, S.; Cuvelier, G.; Attia, H.; Ayadi, M.A. Characterization of pectins extracted from pomegranate peel and their gelling properties. Food Chem. 2017, 215, 318–325. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Wang, Y.; Liu, Z.; Ni, Y. Effects of extraction methods on the structural characteristics and functional properties of dietary fiber extracted from kiwifruit (Actinidia deliciosa). Food Hydrocoll. 2021, 110, 106162. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, N.; Wu, C.; Zhang, H.; Wang, C.; Zhang, T.; Lei, H. Physical modifications of dietary fibers from kiwifruit pomace: Physicochemical, structural and functional properties. Food Chem. 2025, 484, 144422. [Google Scholar] [CrossRef]
- Lin, D.; Long, X.; Huang, Y.; Yang, Y.; Wu, Z.; Chen, H.; Zhang, Q.; Wu, D.; Qin, W.; Tu, Z. Effects of microbial fermentation and microwave treatment on the composition, structural characteristics, and functional properties of modified okara dietary fiber. LWT 2020, 123, 109059. [Google Scholar] [CrossRef]
- Lesa, K.N.; Ahmad, N.; Mayangsari, Y.; Khandaker, M.U.; Iqbal, F.M.R.; Fibri, D.L.N.; Hassan, M.M.; Omara, T. Health Benefits of Okara for the Management of Diabetes Mellitus. J. Food Qual. 2023, 2023, 5540118. [Google Scholar] [CrossRef]
- He, Y.; Li, W.; Zhang, X.; Li, T.; Ren, D.; Lu, J. Physicochemical, functional, and microstructural properties of modified insoluble dietary fiber extracted from rose pomace. J. Food Sci. Technol. 2020, 57, 1421–1429. [Google Scholar] [CrossRef]
- Chen, Z.W.; Xia, Q.; Zha, B.P.; Sun, J.; Xu, B.; Chen, Z.X. Triboelectric separation of wheat bran tissues: Influence of tribo-material, water content, and particle size. J. Food Process Eng. 2020, 43, e13346. [Google Scholar] [CrossRef]
- Golbaghi, L.; Khamforoush, M.; Hatami, T. Carboxymethyl cellulose production from sugarcane bagasse with steam explosion pulping: Experimental, modeling, and optimization. Carbohydr. Polym. 2017, 174, 780–788. [Google Scholar] [CrossRef]
- Ma, C.; Ni, L.; Guo, Z.; Zeng, H.; Wu, M.; Zhang, M.; Zheng, B. Principle and Application of Steam Explosion Technology in Modification of Food Fiber. Foods 2022, 11, 3370. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.K.; Wu, L.X.; Cai, W.D.; Xiao, G.S.; Duan, Y.Q.; Zhang, H.H. Subcritical water extraction-based methods affect the physicochemical and functional properties of soluble dietary fibers from wheat bran. Food Chem. 2019, 298, 124987. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, J.; Yao, R.; Yan, S.; Wang, Q. Mechanism of lipid metabolism regulation by soluble dietary fibre from micronized and non-micronized powders of lotus root nodes as revealed by their adsorption and activity inhibition of pancreatic lipase. Food Chem. 2020, 305, 125435. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, B.; Shi, P.; Tian, H.; Li, Y.; Wang, X.; Wu, S.; Liang, P. The influences of acetylation, hydroxypropylation, enzymatic hydrolysis and crosslinking on improved adsorption capacities and in vitro hypoglycemic properties of millet bran dietary fibre. Food Chem. 2022, 368, 130883. [Google Scholar] [CrossRef] [PubMed]
- McRorie, J.W., Jr.; McKeown, N.M. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Xu, J.; Wu, W.; Wen, Y.; Lu, S.; El-Seedi, H.R.; Zhao, C. Structure-immunomodulatory activity relationships of dietary polysaccharides. Curr. Res. Food Sci. 2022, 5, 1330–1341. [Google Scholar] [CrossRef]
- Wang, K.; Wang, Y.; Chen, S.; Gu, J.; Ni, Y. Insoluble and Soluble Dietary Fibers from Kiwifruit (Actinidia deliciosa) Modify Gut Microbiota to Alleviate High-Fat Diet and Streptozotocin-Induced TYPE 2 Diabetes in Rats. Nutrients 2022, 14, 16. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Xiao, X.; Dong, Y.; Xu, T.; Wu, F. Dietary supplementation with Lactobacillus plantarum dy-1 fermented barley suppresses body weight gain in high-fat diet-induced obese rats. J. Sci. Food Agric. 2016, 96, 4907–4917. [Google Scholar] [CrossRef]
- Ang, W.S.; Law, J.W.F.; Letchumanan, V.; Hong, K.W.; Wong, S.H.; Ab Mutalib, N.S.; Chan, K.G.; Lee, L.H.; Tan, L.T.H. A Keystone Gut Bacterium Christensenella minuta—A Potential Biotherapeutic Agent for Obesity and Associated Metabolic Diseases. Foods 2023, 12, 13. [Google Scholar] [CrossRef]
- Bernaud, F.S.R.; Rodrigues, T.C. Fibra alimentar: Ingestão adequada e efeitos sobre a saúde do metabolismo. Arq. Bras. Endocrinol. Metab. 2013, 57, 6. [Google Scholar] [CrossRef]
- Farag, M.A.; Xiao, J.; Abdallah, H.M. Nutritional value of barley cereal and better opportunities for its processing as a value-added food: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2022, 62, 1092–1104. [Google Scholar] [CrossRef]
- Feng, Y.Q.; Song, Y.T.; Zhou, J.; Duan, Y.Q.; Kong, T.Y.; Ma, H.L.; Zhang, H.H. Recent progress of Lycium barbarum polysaccharides on intestinal microbiota, microbial metabolites and health: A review. Crit. Rev. Food Sci. Nutr. 2024, 64, 2917–2940. [Google Scholar] [CrossRef]
- Li, B.Y.; Xu, X.Y.; Gan, R.Y.; Sun, Q.C.; Meng, J.M.; Shang, A.; Mao, Q.Q.; Li, H.B. Targeting Gut Microbiota for the Prevention and Management of Diabetes Mellitus by Dietary Natural Products. Foods 2019, 8, 440. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.T.; Ding, Y.W.; Hu, Z.K.; Zhang, Z.H.; Fu, L.; Zhang, J.Y.; Zhu, Y.; Bai, J.; Xiao, X. Inter-individual variation in human microbiota drives differential impacts on the fermentability of insoluble bran by soluble (3-glucans from whole barley. Food Hydrocoll. 2025, 162, 111034. [Google Scholar] [CrossRef]
- Zhu, Y.; Bai, J.; Zhang, Y.; Xiao, X.; Dong, Y. Effects of bitter melon (Momordica charantia L.) on the gut microbiota in high fat diet and low dose streptozocin-induced rats. Int. J. Food Sci. Nutr. 2016, 67, 686–695. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Tuncil, Y.E. A perspective on the complexity of dietary fiber structures and their potential effect on the gut microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef]
- Geng, A.L.; Li, N.A.; Zayas-Garriga, A.; Xie, R.R.; Zhu, D.C.; Sun, J.Z. Direct Conversion of Minimally Pretreated Corncob by Enzyme-Intensified Microbial Consortia. Agriculture 2024, 14, 9. [Google Scholar] [CrossRef]
- Long, P.; Zeng, S.; Ying, R.; Huang, M. Mechanistic Insights Into the Inhibitory Effects of Arabinoxylan and (1,3)(1,4)-β-Glucan on Starch Digestive Enzymes. J. Food Sci. 2025, 90, e70221. [Google Scholar] [CrossRef]
- Hattori, K.; Arai, A. Preparation and Hydrolysis of Water-Stable Amorphous Cellulose. ACS Sustain. Chem. Eng. 2016, 4, 1180–1186. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Dou, Y.; Yu, X.; Luo, Y.; Chen, B.; Ma, D.; Zhu, J. Effect of Fructooligosaccharides Supplementation on the Gut Microbiota in Human: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhao, J.; Al-Ansi, W.; Wang, J.; Xue, L.; Liu, J.; Wang, Y.; Fan, M.; Qian, H.; Li, Y. Oat β-glucan alleviates DSS-induced colitis via regulating gut microbiota metabolism in mice. Food Funct. 2021, 12, 8976–8993. [Google Scholar] [CrossRef] [PubMed]
- Demuth, T.; Edwards, V.; Bircher, L.; Lacroix, C.; Nyström, L.; Geirnaert, A. In vitro Colon Fermentation of Soluble Arabinoxylan Is Modified Through Milling and Extrusion. Front. Nutr. 2021, 8, 707763. [Google Scholar] [CrossRef]
- Geng, N.; Song, J.; Zhang, K.; Dai, Z.; Li, D. Effect of dynamic high-pressure microfluidization on the physicochemical and structural properties of insoluble dietary fiber from fresh corn bract. J. Food Process. Pres. 2021, 45, 9. [Google Scholar] [CrossRef]
- Geng, N.; Song, J.; Luo, S.; Li, Y.; Wu, G.; Liu, C.; Wu, C. Ultrasound-assisted enzymatic extraction of soluble dietary fiber from fresh corn bract and its physio-chemical and structural properties. Qual. Assur. Saf. Crops Foods 2022, 14, 119–130. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhou, X.Y.; Zhang, Y.S.; Zhan, E.Q.; Ouyang, Z.; Bai, J.; Zhao, Y.S.; Xiao, X. Impacts of superfine grinding on structural characteristics and lipid-lowering effect of bitter melon polysaccharides. Int. J. Food Sci. Technol. 2024, 59, 3813–3822. [Google Scholar] [CrossRef]
- Wu, C.; Chen, P.; Li, S.; Jiang, L.; Wang, Z.; Liu, J. Effect of Cavitation Jets on Structure and Function of Okara Insoluble Dietary Fiber. Trans. Chin. Soc. Agric. Mach. 2021, 52, 350–356. [Google Scholar]
- Wu, C.L.; Teng, F.; McClements, D.J.; Zhang, S.; Li, Y.; Wang, Z.J. Effect of cavitation jet processing on the physicochemical properties and structural characteristics of okara dietary fiber. Food Res. Int. 2020, 134, 109251. [Google Scholar] [CrossRef]
- Li, B.; Yang, W.; Nie, Y.; Kang, F.; Goff, H.D.; Cui, S.W. Effect of steam explosion on dietary fiber, polysaccharide, protein and physicochemical properties of okara. Food Hydrocoll. 2019, 94, 48–56. [Google Scholar] [CrossRef]
- Huang, J.-Q.; Qi, R.-T.; Pang, M.-R.; Liu, C.; Li, G.-Y.; Zhang, Y. Isolation, chemical characterization, and immunomodulatory activity of naturally acetylated hemicelluloses from bamboo shavings. J. Zhejiang Univ. Sci. B 2017, 18, 138–151. [Google Scholar] [CrossRef]
- Yan, L.; Li, T.; Liu, C.; Zheng, L. Effects of high hydrostatic pressure and superfine grinding treatment on physicochemical/functional properties of pear pomace and chemical composition of its soluble dietary fibre. LWT 2019, 107, 171–177. [Google Scholar] [CrossRef]
- Perumal, A.B.; Nambiar, R.B.; Moses, J.A.; Anandharamakrishnan, C. Nanocellulose: Recent trends and applications in the food industry. Food Hydrocoll. 2022, 127, 107484. [Google Scholar] [CrossRef]
- Ahmad, K.; Din, Z.-U.; Ullah, H.; Ouyang, Q.; Rani, S.; Jan, I.; Alam, M.; Rahman, Z.; Kamal, T.; Ali, S.; et al. Preparation and Characterization of Bio-based Nanocomposites Packaging Films Reinforced with Cellulose Nanofibers from Unripe Banana Peels. Starch-Starke 2022, 74, 5–6. [Google Scholar] [CrossRef]
- Ghaderi, M.; Mousavi, M.; Yousefi, H.; Labbafi, M. All-cellulose nanocomposite film made from bagasse cellulose nanofibers for food packaging application. Carbohydr. Polym. 2014, 104, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Stijnman, A.C.; Bodnar, I.; Hans Tromp, R. Electrospinning of food-grade polysaccharides. Food Hydrocoll. 2011, 25, 1393–1398. [Google Scholar] [CrossRef]
- Shishir, M.R.I.; Saifullah, M.; Hashim, S.B.H.; Aalim, H.; Bilal, M.; Khan, S.; Marappan, G.; Tahir, H.E.; Zhihua, L.; Zhai, X.; et al. Micro and nano-encapsulated natural products in yogurt: An emerging trend to achieve multifunctional benefits in product quality and human health. Food Hydrocoll. 2024, 154, 110124. [Google Scholar] [CrossRef]
- Lubambo, A.F.; de Freitas, R.A.; Sierakowski, M.-R.; Lucyszyn, N.; Sassaki, G.L.; Serafim, B.M.; Saul, C.K. Electrospinning of commercial guar-gum: Effects of purification and filtration. Carbohydr. Polym. 2013, 93, 484–491. [Google Scholar] [CrossRef]
- Im, W.; Oh, K.; Rajabi Abhari, A.; Youn, H.J.; Lee, H.L. Recycling of isopropanol for cost-effective, environmentally friendly production of carboxymethylated cellulose nanofibrils. Carbohydr. Polym. 2019, 208, 365–371. [Google Scholar] [CrossRef]
- Yang, G.; Xia, Y.; Lin, Z.; Zhang, K.; Fatehi, P.; Chen, J. Physicochemical impact of cellulose nanocrystal on oxidation of starch and starch based composite films. Int. J. Biol. Macromol. 2021, 184, 42–49. [Google Scholar] [CrossRef]
- Quan, W.; Wang, J.; Huang, J.; Zhang, D. Structure Characterization and Dye Adsorption Properties of Modified Fiber from Wheat Bran. Molecules 2024, 29, 2581. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.J.; Wang, L.; Dong, L.; Yin, M.Y.; Wei, S.F.; Luo, P. Agrocybe cylindracea Dietary Fiber Modification: Sodium Hydroxide Treatment Outperforms High-Temperature, Cellulase, and Lactobacillus Fermentation. Molecules 2024, 29, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liao, J.; Qi, J. Functional and structural properties of dietary fiber from citrus peel affected by the alkali combined with high-speed homogenization treatment. LWT 2020, 128, 109397. [Google Scholar] [CrossRef]
- Liao, Y.; Alam, N.; Fatehi, P. Semitransparent films from low-substituted carboxymethylated cellulose fibers. J. Mater. Sci. 2022, 57, 10407–10424. [Google Scholar] [CrossRef]
- Zhang, J.-G.; Yang, G.; Zhang, W.-W.; Thakur, K.; Hu, F.; Khan, M.R.; Ni, Z.-J.; Wei, Z.-J. Physicochemical and functional properties of carboxymethylated insoluble dietary fiber of Lycium barbarum seed dreg. Food Chem. X 2024, 22, 101270. [Google Scholar] [CrossRef] [PubMed]
- Rani, K.; Gomathi, T.; Vijayalakshmi, K.; Saranya, M.; Sudha, P.N. Banana fiber Cellulose Nano Crystals grafted with butyl acrylate for heavy metal lead (II) removal. Int. J. Biol. Macromol. 2019, 131, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, X.; Tian, H.; Li, Y.; Shi, P.; Guo, W.; Zhu, Q. Effect of four modification methods on adsorption capacities and in vitro hypoglycemic properties of millet bran dietary fibre. Food Res. Int. 2021, 147, 110565. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Kotera, M.; Suetsugu, M.; Murakami, H.; Urushihara, Y. Acetylation of plant cellulose fiber in supercritical carbon dioxide. Polymer 2011, 52, 830–836. [Google Scholar] [CrossRef]
- Tserki, V.; Zafeiropoulos, N.E.; Simon, F.; Panayiotou, C. A study of the effect of acetylation and propionylation surface treatments on natural fibres. Compos. Part A Appl. Sci. Manuf. 2005, 36, 1110–1118. [Google Scholar] [CrossRef]
- Su, J.; Zhang, C.; Fan, F. Sulfation Modification and Properties Analysis of Soluble Dietary Fiber from Rosa sterilis Pomace. Sci. Technol. Food Ind. 2023, 44, 255–261. [Google Scholar] [CrossRef]
- Mo, Q.; Dai, L.H.; Ma, J.J.; Zhao, X.J.; Zhu, L. Preparation and Physiological activities of sulphated derivative extracted from corn bran. IOP Conf. Ser. Earth Environ. Sci. 2017, 64, 012120. [Google Scholar] [CrossRef]
- Xu, Y.F.; Song, S.; Wei, Y.X.; Wang, F.X.; Zhao, M.; Guo, J.; Zhang, J. Sulfated modification of the polysaccharide from Sphallerocarpus gracilis and its antioxidant activities. Int. J. Biol. Macromol. 2016, 87, 180–190. [Google Scholar] [CrossRef]
- Tuly, J.A.; Ma, H. Bioconversion of food industrial waste okara by microbial fermentation: Scope of omics study and possibility. Trends Food Sci. Technol. 2024, 146, 10439. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Yi, C.; Quan, K.; Lin, B. Chemical composition, structure, physicochemical and functional properties of rice bran dietary fiber modified by cellulase treatment. Food Chem. 2021, 342, 128352. [Google Scholar] [CrossRef]
- Xie, F.; Zhao, T.; Wan, H.; Li, M.; Sun, L.; Wang, Z.; Zhang, S. Structural and Physicochemical Characteristics of Rice Bran Dietary Fiber by Cellulase and High-Pressure Homogenization. Appl. Sci. 2019, 9, 1270. [Google Scholar] [CrossRef]
- Ma, R.; Chen, J.-N.; Zhou, X.-J.; Lin, H.; Gao, Q.; Peng, X.; Tanokura, M.; Xue, Y. Effect of chemical and enzymatic modifications on the structural and physicochemical properties of dietary fiber from purple turnip (Brassica rapa L.). LWT 2021, 145, 111313. [Google Scholar] [CrossRef]
- Ye, G.; Wu, Y.; Wang, L.; Tan, B.; Shen, W.; Li, X.; Liu, Y.; Tian, X.; Zhang, D. Comparison of six modification methods on the chemical composition, functional properties and antioxidant capacity of wheat bran. LWT 2021, 149, 111996. [Google Scholar] [CrossRef]
- Xiao, X.; Li, X.; Bai, J.; Fan, S.; Daglia, M.; Li, J.; Ding, Y.; Zhang, Y.; Zhao, Y. Changes in the structural, physicochemical and functional properties and in vitro fecal fermentation characteristics of barley dietary fiber fermented by Lactiplantibacillus plantarum dy-1. Food Funct. 2024, 15, 4276–4291. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Bai, J.; Zhang, J.; Wu, J.; Dong, Y. Inhibitory effect of fermented selected barley extracts with Lactobacillus plantarum dy-1 on the proliferation of human HT-29 Cells. J. Food Biochem. 2019, 43, e12989. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, Z.; Nie, Q.; Ackah, M.; Nie, S. Rethinking the classification of non-digestible carbohydrates: Perspectives from the gut microbiome. Crit. Rev. Food Sci. Nutr. 2024, 23, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ba, K.; Li, X.; He, Y.; Zhang, Y.; Ai, L.; Zhang, J.; Zhao, Y.; Xiao, X. Comparative analysis of barley dietary fiber fermented with and without Lactiplantibacillus plantarum dy-1 in promoting gut health and regulating hepatic energy metabolism in high-fat diet-induced obese mice. Food Funct. 2025, 16, 219–231. [Google Scholar] [CrossRef]
- Meng, W.; Hu, M.; Zhang, P.; Wang, J.; Yuan, Z.; Wang, F.; Li, S. Efficient conversion of insoluble dietary fiber to soluble dietary fiber by Bacillus subtilis BSNK-5 fermentation of okara and improvement of their structural and functional properties. Food Chem. 2025, 474, 143188. [Google Scholar] [CrossRef]
- Jung, J.T.; Lee, J.K.; Choi, Y.S.; Lee, J.H.; Choi, J.S.; Choi, Y.I.; Chung, Y.K. Effect of Rice Bran and Wheat Fibers on Microbiological and Physicochemical Properties of Fermented Sausages during Ripening and Storage. Korean J. Food Sci. Anim. Resour. 2018, 38, 302–314. [Google Scholar] [CrossRef]
- Jia, M.; Chen, J.; Liu, X.; Xie, M.; Nie, S.; Chen, Y.; Xie, J.; Yu, Q. Structural characteristics and functional properties of soluble dietary fiber from defatted rice bran obtained through Trichoderma viride fermentation. Food Hydrocoll. 2019, 94, 468–474. [Google Scholar] [CrossRef]
- Niu, Y.; Li, N.; Xia, Q.; Hou, Y.; Xu, G. Comparisons of three modifications on structural, rheological and functional properties of soluble dietary fibers from tomato peels. LWT 2018, 88, 56–63. [Google Scholar] [CrossRef]
- Chu, J.; Zhao, H.; Lu, Z.; Lu, F.; Bie, X.; Zhang, C. Improved physicochemical and functional properties of dietary fiber from millet bran fermented by Bacillus natto. Food Chem. 2019, 294, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Gong, H.; Tang, H.; Meng, Z.; Wang, Z.; Cui, W.; Zhang, K.; Chen, Y.; Yang, Y. Enhanced lignocellulose degradation in Bacillus subtilis RLI2019 through CRISPR/Cas9-mediated chromosomal integration of ternary cellulase genes. Int. J. Biol. Macromol. 2025, 306, 141727. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.D.; Liu, S.Z.; Duan, Y.Z.; Bao, C.L.; Wang, J.; Dong, B.; Cao, Y.H. Complete genome sequencing and investigation on the fiber-degrading potential of Bacillus amyloliquefaciens strain TL106 from the tibetan pig. BMC Microbiol. 2022, 22, 186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Luo, L.; Duan, X.; Li, H.; Zhao, Y.; Yang, H.; Ma, X. Preparation of Soluble Dietary Fiber from Mung Bean Hull by High-Temperature Cooking and Compound Enzyme Method and Its in vitro Hypoglycemic Effect. J. Chin. Cereals Oils Assoc. 2022, 37, 59–66. [Google Scholar] [CrossRef]
- Wang, N.; Huang, S.; Zhang, Y.; Zhang, F.; Zheng, J. Modification of bamboo shoot dietary fiber by high temperature cooking combined with cellulase. Food Ferment. Ind. 2020, 46, 13–18. [Google Scholar] [CrossRef]
- Guo, T.; He, Z.; Wang, F.; Yuen, M.; Yuen, H.; Peng, Q. Effects of enzymes combined with Lactobacillus plantarum and Bifidobacterium on the properties and in vitro hypoglycemic activity of sea buckthorn insoluble dietary fiber. Food Biosc. 2025, 68, 106472. [Google Scholar] [CrossRef]
- Si, J.; Yang, C.; Chen, Y.; Xie, J.; Tian, S.; Cheng, Y.; Hu, X.; Yu, Q. Structural properties and adsorption capacities of Mesona chinensis Benth residues dietary fiber prepared by cellulase treatment assisted by Aspergillus niger or Trichoderma reesei. Food Chem. 2023, 407, 135149. [Google Scholar] [CrossRef]
- Cornfine, C.; Hasenkopf, K.; Eisner, P.; Schweiggert, U. Influence of chemical and physical modification on the bile acid binding capacity of dietary fibre from lupins (Lupinus angustifolius L.). Food Chem. 2010, 122, 638–644. [Google Scholar] [CrossRef]
- Tang, W.; Lin, X.; Walayat, N.; Liu, J.; Zhao, P. Dietary fiber modification: Structure, physicochemical properties, bioactivities, and application—A review. Crit. Rev. Food Sci. Nutr. 2024, 64, 7895–7915. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, F.; Yang, J.; Wu, L.; Zheng, J. Application of ultrasonic technology in extraction and modification of dietary fiber. Food Ferment. Ind. 2023, 49, 330–336. [Google Scholar] [CrossRef]
- Li, Z.; Dong, Y.; Xiao, X.; Zhou, X.-H. Mechanism by which β-glucanase improves the quality of fermented barley flour-based food products. Food Chem. 2020, 311, 126026. [Google Scholar] [CrossRef]
- Li, X.L.; Wu, Y.C.; Duan, R.L.; Yu, H.R.; Liu, S.Y.; Bao, Y.L. Research Progress in the Extraction, Structural Characteristics, Bioactivity, and Commercial Applications of Oat β-Glucan: A Review. Foods 2024, 13, 4160. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Q.; Tan, B.; Wang, L. Individual and combined effects of cold plasma and enzymatic hydrolysis modification on soluble dietary fiber in wheat bran: Structural, physicochemical and functional properties. J. Cereal Sci. 2025, 123, 104196. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Y.; Wu, J.; Xu, Y.; Xiao, G.; Li, L.; Liu, H. Comparison the Structural, Physicochemical, and Prebiotic Properties of Litchi Pomace Dietary Fibers before and after Modification. Foods 2022, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, H.N.; Ma, A.M.; Zhou, J.Z.; Xia, X.D. Synergetic effects of Lactobacillus plantarum and Rhizopus oryzae on physicochemical, nutritional and antioxidant properties of whole-grain oats (Avena sativa L.) during solid-state fermentation. LWT Food Sci. Technol. 2022, 154, 112687. [Google Scholar] [CrossRef]
- Meng, K.; Wang, Y.; Liu, F.; Zhan, Q.; Zhao, L. Effect of modifications on structure, physicochemical properties and lead ions adsorption behavior of dietary fiber of Flammulina velutipes. Food Chem. 2025, 464, 141597. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Li, Y.; Yu, H.; Wang, Y.; Piao, C. Insoluble dietary fibre from okara (soybean residue) modified by yeast Kluyveromyces marxianus. LWT 2020, 134, 110252. [Google Scholar] [CrossRef]
- Huang, Y.-L.; Ma, Y.-S. The effect of extrusion processing on the physiochemical properties of extruded orange pomace. Food Chem. 2016, 192, 363–369. [Google Scholar] [CrossRef]
- Wei, X.; Jiang, C.; Liu, X.; Liu, H.; Wang, J.; Zheng, X.; Zhang, Z.; Hu, H. Effect of γ-irradiation combined with enzymatic modification on the physicochemical properties of defatted rice bran dietary fiber. Food Chem. X 2024, 24, 101975. [Google Scholar] [CrossRef]
- Ferreira-Lazarte, A.; Kachrimanidou, V.; Villamiel, M.; Rastall, R.A.; Moreno, F.J. In vitro fermentation properties of pectins and enzymatic-modified pectins obtained from different renewable bioresources. Carbohydr. Polym. 2018, 199, 482–491. [Google Scholar] [CrossRef]
- Lai, F.; Wen, Q.; Li, L.; Wu, H.; Li, X. Antioxidant activities of water-soluble polysaccharide extracted from mung bean (Vigna radiata L.) hull with ultrasonic assisted treatment. Carbohydr. Polym. 2010, 81, 323–329. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Luo, G.; Lei, A.; Chen, H.; Zhang, F. Effects of high-pressure homogenization on rheological properties of orange juice. Food Ferment. Ind. 2021, 47, 22–29. [Google Scholar] [CrossRef]
- Juodeikiene, G.; Trakselyte-Rupsiene, K.; Navickaite, B.; Zadeike, D.; Bendoraitiene, J.; Bartkiene, E.; Lele, V.; Rueller, L.; Robert, J.; Arnoldi, A. Functionalization of soya press cake (okara) by ultrasonication for enhancement of submerged fermentation with Lactobacillus paracasei LUHS244 for wheat bread production. LWT 2021, 152, 112337. [Google Scholar] [CrossRef]
- Wu, X.; Li, M.X.; Xiao, Z.G.; Daglia, M.; Dragan, S.; Delmas, D.; Vong, C.T.; Wang, Y.T.; Zhao, Y.S.; Shen, J.; et al. Dietary polyphenols for managing cancers: What have we ignored? Trends Food Sci. Technol. 2020, 101, 150–164. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Currenti, W.; Micek, A.; Falzone, L.; Libra, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Quiles, J.L.; Battino, M.; et al. The Effect of Dietary Polyphenols on Vascular Health and Hypertension: Current Evidence and Mechanisms of Action. Nutrients 2022, 14, 545. [Google Scholar] [CrossRef]
- Lei, D.; Li, J.; Li, S.; Zhu, Z.; He, J. Stability and Fat Adsorption Activity of the Complex of Soluble Dietary Fiber and Polyphenols from Lotus Root. J. Chin. Inst. Food Sci. Technol. 2022, 22, 31–39. [Google Scholar] [CrossRef]
- Liu, S.; Yu, Q.; Huang, H.; Hou, K.; Dong, R.; Chen, Y.; Xie, J.; Nie, S.; Xie, M. The effect of bound polyphenols on the fermentation and antioxidant properties of carrot dietary fiber in vivo and in vitro. Food Funct. 2020, 11, 748–758. [Google Scholar] [CrossRef]
- Zhang, H.; Troise, A.D.; Qi, Y.; Wu, G.; Zhang, H.; Fogliano, V. Insoluble dietary fibre scavenges reactive carbonyl species under simulated physiological conditions: The key role of fibre-bound polyphenols. Food Chem. 2021, 349, 129018. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Blancas-Benítez, F.J.; Sáyago-Ayerdi, S.G. Polyphenols associated with dietary fibers in plant foods: Molecular interactions and bioaccessibility. Curr. Opin. Food Sci. 2017, 13, 84–88. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Yu, Y.; Chen, W.; Liu, Y.; Yu, H. Nanoformulations of quercetin and cellulose nanofibers as healthcare supplements with sustained antioxidant activity. Carbohydr. Polym. 2019, 207, 160–168. [Google Scholar] [CrossRef]
- Li, S.; Chen, G.; Shi, X.; Ma, C.; Liu, F. Comparative Study of Heat- and Enzyme-Induced Emulsion Gels Formed by Gelatin and Whey Protein Isolate: Physical Properties and Formation Mechanism. Gels 2022, 8, 212. [Google Scholar] [CrossRef]
- Wu, D.; Wang, H.; Guo, X.; Zhang, Z.; Gao, Z.; Gao, S.; Liu, Z.; Rao, S.; Meng, X. Insight into the mechanism of enhancing myofibrillar protein gel hardness by ultrasonic treatment combined with insoluble dietary fiber from oat. LWT Food Sci. Technol. 2023, 178, 114539. [Google Scholar] [CrossRef]
- Ye, Z.; Gan, S.; Wang, J.; Yang, F.; Dong, G. Conjugation of flaxseed protein and plant polysaccharides: Process optimization, structural characterization and technical-function evaluation. Food Hydrocoll. 2025, 166, 111371. [Google Scholar] [CrossRef]
- Fu, G.-M.; Xu, Z.-W.; Luo, C.; Xu, L.-Y.; Chen, Y.-R.; Guo, S.-L.; Wu, X.-D.; Wan, Y. Modification of soy protein isolate by Maillard reaction and its application in microencapsulation of Limosilactobacillus reuteri. J. Biosci. Bioeng. 2021, 132, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Zhang, Q.; Li, X.; Xue, B.; Sun, T.; Xie, J. Effect of Oat β-Glucan on the Structure and Properties of Soybean Protein Isolate During Maillard Reaction. Plant Foods Hum. Nutr. 2023, 78, 552–556. [Google Scholar] [CrossRef]
- Lin, L.; Wu, J.J.; Li, C.Z.; Chen, X.C.; Cui, H.Y. Fabrication of a dual-response intelligent antibacterial nanofiber and its application in beef preservation. LWT Food Sci. Technol. 2022, 154, 112606. [Google Scholar] [CrossRef]
- Zhang, J.J.; Huang, X.W.; Zhang, J.N.; Liu, L.; Shi, J.Y.; Muhammad, A.; Zhai, X.D.; Zou, X.B.; Xiao, J.B.; Li, Z.H.; et al. Development of nanofiber indicator with high sensitivity for pork preservation and freshness monitoring. Food Chem. 2022, 381, 132224. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.M.; Hu, W.; Yang, H.Y.; Li, C.Z.; Cui, H.Y.; Li, X.Z.; Lin, L. Controlled release and antibacterial properties of PEO/casein nanofibers loaded with Thymol/β-cyclodextrin inclusion complexes in beef preservation. Food Chem. 2022, 382, 132369. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Z.; Bai, M.; Chen, X.C.; Hu, W.; Cui, H.Y.; Lin, L. Controlled release and antibacterial activity of nanofibers loaded with basil essential oil-encapsulated cationic liposomes against Listeria monocytogenes. Food Biosci. 2022, 46, 101578. [Google Scholar] [CrossRef]
- Lin, L.; Luo, C.C.; Li, C.Z.; Abdel-Samie, M.A.; Cui, H.Y. Eugenol/silk fibroin nanoparticles embedded Lycium barbarum polysaccharide nanofibers for active food packaging. Food Packag. Shelf Life 2022, 32, 100841. [Google Scholar] [CrossRef]
- Karimi, T.; Mottaghitalab, F.; Keshvari, H.; Farokhi, M. Carboxymethyl chitosan/sodium carboxymethyl cellulose/agarose hydrogel dressings containing silk fibroin/polydopamine nanoparticles for antibiotic delivery. J. Drug Deliv. Sci. Technol. 2023, 80, 104134. [Google Scholar] [CrossRef]
- Lohani, A.; Saxena, R.; Khan, S.; Mascarenhas-Melo, F. pH-responsive IPN beads of carboxymethyl konjac glucomannan and sodium carboxymethyl cellulose as a controlled release carrier for ibuprofen. Int. J. Biol. Macromol. 2024, 278, 134676. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, J.; Zhang, R.; Yan, C.; Cui, L.; Zhu, J. A pH-responsive carboxymethyl cellulose/chitosan hydrogel for adsorption and desorption of anionic and cationic dyes. Cellulose 2021, 28, 897–909. [Google Scholar] [CrossRef]
- Mu, R.J.; Hong, X.; Ni, Y.S.; Li, Y.Z.; Pang, J.; Wang, Q.; Xiao, J.B.; Zheng, Y.F. Recent trends and applications of cellulose nanocrystals in food industry. Trends Food Sci. Technol. 2019, 93, 136–144. [Google Scholar] [CrossRef]
- Berg, B. Cellulose degradation and cellulase formation by Phialophora malorum. Arch. Microbiol. 1978, 118, 61–65. [Google Scholar] [CrossRef]
- Fan, S.T.; Zhang, Z.H.; Zhao, Y.S.; Daglia, M.; Zhang, J.Y.; Zhu, Y.; Bai, J.; Zhu, L.; Xiao, X. Recent advances in targeted manipulation of the gut microbiome by prebiotics: From taxonomic composition to metabolic function. Curr. Opin. Food Sci. 2023, 49, 100959. [Google Scholar] [CrossRef]
- Delannoy-Bruno, O.; Desai, C.; Raman, A.S.; Chen, R.Y.; Hibberd, M.C.; Cheng, J.; Han, N.; Castillo, J.J.; Couture, G.; Lebrilla, C.B.; et al. Evaluating microbiome-directed fibre snacks in gnotobiotic mice and humans. Nature 2021, 595, 91–95. [Google Scholar] [CrossRef]
- Quraishi, U.M.; Murat, F.; Abrouk, M.; Pont, C.; Confolent, C.; Oury, F.X.; Ward, J.; Boros, D.; Gebruers, K.; Delcour, J.A.; et al. Combined meta-genomics analyses unravel candidate genes for the grain dietary fiber content in bread wheat (Triticum aestivum L.). Funct. Integr. Genom. 2011, 11, 71–83. [Google Scholar] [CrossRef]
- Maldonado-Gomez, M.X.; Ng, K.M.; Drexler, R.A.; Conner, A.M.S.; Vierra, C.G.; Krishnakumar, N.; Gerber, H.M.; Taylor, Z.R.; Treon, J.L.; Ellis, M.; et al. A diverse set of solubilized natural fibers drives structure-dependent metabolism and modulation of the human gut microbiota. mBio 2025, 16, e00425–e00470. [Google Scholar] [CrossRef]
- Baroni, L.; Rizzo, G.; Zavoli, M.; Battino, M. A Plant-Based Food Guide Adapted for Low-Fat Diets: The VegPlate Low-Fat (VP_LF). Foods 2024, 13, 4050. [Google Scholar] [CrossRef] [PubMed]
- Perrigue, M.; Carter, B.; Roberts, S.A.; Drewnowski, A. A Low-Calorie Beverage Supplemented with Low-Viscosity Pectin Reduces Energy Intake at a Subsequent Meal. J. Food Sci. 2010, 75, 300–305. [Google Scholar] [CrossRef]
- Singla, A.; Gupta, O.P.; Sagwal, V.; Kumar, A.; Patwa, N.; Mohan, N.; Ankush; Kumar, D.; Vir, O.; Singh, J.; et al. Beta-Glucan as a Soluble Dietary Fiber Source: Origins, Biosynthesis, Extraction, Purification, Structural Characteristics, Bioavailability, Biofunctional Attributes, Industrial Utilization, and Global Trade. Nutrients 2024, 16, 900. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, L.; Rioux, L.-E.; Britten, M.; Turgeon, S.L. In vitro bioaccessibility of peptides and amino acids from yogurt made with starch, pectin, or β-glucan. Int. Dairy J. 2015, 46, 39–45. [Google Scholar] [CrossRef]
- Usman, M.; Ahmed, S.; Mehmood, A.; Bilal, M.; Patil, P.J.; Akram, K.; Farooq, U. Effect of apple pomace on nutrition, rheology of dough and cookies quality. J. Food Sci. Technol. 2020, 57, 3244–3251. [Google Scholar] [CrossRef]
- Aggarwal, D.; Sabikhi, L.; Kumar, M.H.S.; Panjagari, N.R. Investigating the effect of resistant starch, polydextrose and biscuit improver on the textural and sensory characteristics of dairy-multigrain composite biscuits using response surface methodology. J. Food Meas. Charact. 2018, 12, 1167–1176. [Google Scholar] [CrossRef]
- Ghaedrahmati, S.; Shahidi, F.; Roshanak, S.; Mahallati, M.N. Application of jaban watermelon exocarp powder in low-calorie ice cream formulation and evaluation of its physicochemical, rheological, and sensory properties. J. Food Process. Pres. 2021, 45, 15768. [Google Scholar] [CrossRef]
- Wu, D.-T.; Wang, J.; Li, J.; Hu, J.-L.; Yan, H.; Zhao, J.; Zou, L.; Hu, Y.-C. Physicochemical properties and biological functions of soluble dietary fibers isolated from common and Tartary buckwheat sprouts. LWT Food Sci. Technol. 2023, 183, 114944. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, X.; Cao, X.; Liu, T. The structural characteristics of dietary fibers from Tremella fuciformis and their hypolipidemic effects in mice. Food Sci. Hum. Wellness 2023, 12, 503–511. [Google Scholar] [CrossRef]
- Teparak, C.; Uriyapongson, J.; Phoemsapthawee, J.; Tunkamnerdthai, O.; Aneknan, P.; Tong-un, T.; Panthongviriyakul, C.; Leelayuwat, N.; Alkhatib, A. Diabetes Therapeutics of Prebiotic Soluble Dietary Fibre and Antioxidant Anthocyanin Supplement in Patients with Type 2 Diabetes: Randomised Placebo-Controlled Clinical Trial. Nutrients 2025, 17, 1098. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, C.; Du, M.; Hu, X.; Yi, J. Effects of gel spatial structure and water state on 3D printability: A case study of honey bee pupa protein and soybean dietary fiber. Food Hydrocoll. 2024, 146, 109224. [Google Scholar] [CrossRef]
- Xie, Y.; Yu, X.; Wang, Y.; Yu, C.; Prakash, S.; Zhu, B.; Dong, X. Role of dietary fiber and flaxseed oil in altering the physicochemical properties and 3D printability of cod protein composite gel. J. Food Eng. 2022, 327, 111053. [Google Scholar] [CrossRef]
- Liu, Y.; Yi, S.; Ye, T.; Leng, Y.; Hossen, M.A.; Sameen, D.E.; Dai, J.; Li, S.; Qin, W. Effects of ultrasonic treatment and homogenization on physicochemical properties of okara dietary fibers for 3D printing cookies. Ultrason. Sonochem. 2021, 77, 105693. [Google Scholar] [CrossRef]
- Williams, G.; Noakes, M.; Keogh, J.; Foster, P.; Clifton, P. High protein high fibre snack bars reduce food intake and improve short term glucose and insulin profiles compared with high fat snack bars. Asia Pac. J. Clin. Nutr. 2006, 15, 443–450. [Google Scholar] [CrossRef]
- Urita, Y.; Noda, T.; Watanabe, D.; Iwashita, S.; Hamada, K.; Sugimoto, M. Effects of a soybean nutrition bar on the postprandial blood glucose and lipid levels in patients with diabetes mellitus. Int. J. Food Sci. Nutr. 2012, 63, 921–929. [Google Scholar] [CrossRef]
- Kowalska, H.; Kowalska, J.; Ignaczak, A.; Masiarz, E.; Domian, E.; Galus, S.; Ciurzyńska, A.; Salamon, A.; Zając, A.; Marzec, A. Development of a High-Fibre Multigrain Bar Technology with the Addition of Curly Kale. Molecules 2021, 26, 3939. [Google Scholar] [CrossRef]
- Kaviani, M.; Chilibeck, P.D.; Gall, S.; Jochim, J.; Zello, G.A. The Effects of Low- and High-Glycemic Index Sport Nutrition Bars on Metabolism and Performance in Recreational Soccer Players. Nutrients 2020, 12, 982. [Google Scholar] [CrossRef]
- Cui, H.Y.; Bai, M.; Rashed, M.M.A.; Lin, L. The antibacterial activity of clove oil/chitosan nanoparticles embedded gelatin nanofibers against Escherichia coli O157:H7 biofilms on cucumber. Int. J. Food Microbiol. 2018, 266, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Kausar, T.; Aslam, J.; Quddoos, M.Y.; Ali, A.; Kauser, S.; Zerlasht, M.; Rafique, A.; Noreen, S.; Iftikhar, K.; et al. Physical and Rheological Studies of Biscuits Developed with Different Replacement Levels of Pumpkin (Cucurbita maxima) Peel, Flesh, and Seed Powders. J. Food Qual. 2023, 2023, 4362094. [Google Scholar] [CrossRef]
- Cappa, C.; Castorina, G.; Fiorillo, G.; Casiraghi, M.C.; Rollini, M.; Consonni, G.; Erba, D.; Negrini, N.; Marti, A. Gluten-free breadsticks with Ganoderma-fermented corncobs: Technological and nutritional features. J. Sci. Food Agric. 2025, 105, 4916–4926. [Google Scholar] [CrossRef] [PubMed]
- Arslan, M.; Rakha, A.; Khan, M.R.; Zou, X.B. Complementing the dietary fiber and antioxidant potential of gluten free bread with guava pulp powder. J. Food Meas. Charact. 2017, 11, 1959–1968. [Google Scholar] [CrossRef]
- Alam, M.; Majid, I.; Kaur, S.; Dar, B.N.; Nanda, V. An Updated Review on Exploring Hydrocolloids Application in Food Matrix: Current Insights Into Fruit, Bakery, Meat, and Dairy Based Products. J. Texture Stud. 2025, 56, 70020. [Google Scholar] [CrossRef] [PubMed]
- Lomer, B.B.; Ghannadiasl, F. Enrichment of Oil Cake with Cinnamon Extract Positively Effects Antioxidant Activity and Textural Profile. Food Sci. Nutr. 2025, 13, 4714. [Google Scholar] [CrossRef]
- Nurko, E.; Nakilcioglu, E. Optimization of Spirulina-Enriched Vegan Cake Formulation Using Response Surface Methodology. Food Sci. Nutr. 2025, 13, 70116. [Google Scholar] [CrossRef]
- Miele, N.A.; Di Monaco, R.; Masi, P.; Cavella, S. Reduced-Calorie Filling Cream: Formula Optimization and Mechanical Characterization. Chem. Eng. Trans. 2015, 43, 67–72. [Google Scholar] [CrossRef]
- Shen, C.; Chen, W.Q.; Li, C.Z.; Ye, Y.; Cui, H.Y.; Lin, L. Preparation and physicochemical effects of zein nanofiber membrane encapsulated with citral/HP-β-CD inclusion complex and its application on cheese. Food Biosci. 2022, 50, 101990. [Google Scholar] [CrossRef]
- Ozmen, D.; Toker, O.S.; Ozturk, S.A. Lemon Fiber as a New Source of Fat Replacement in Filling Creams. J. Food Process. Eng. 2025, 48, 70056. [Google Scholar] [CrossRef]
- Shoaib, M.; Shehzad, A.; Omar, M.; Rakha, A.; Raza, H.; Sharif, H.R.; Shakeel, A.; Ansari, A.; Niazi, S. GRAS, plant- and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Carbohydr. Polym. 2016, 147, 444–454. [Google Scholar] [CrossRef]
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant- and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharvest Biol. Technol. 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Ding, H.; Zheng, J.; Wang, X.; Wang, H. Study on the comparison of key issues of Codex Commission on Food Additives (CCFA) and corresponding food additive management in China. Chin. J. Food Hyg. 2021, 33, 228–233. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Plant Sources | Total Dietary Fiber (%) | Soluble Dietary Fiber (%) | Insoluble Dietary Fiber (%) | Refs. |
|---|---|---|---|---|
| Wheat bran | 59.10 ± 0.2 | 15.50 ± 0.30 | 43.50 ± 0.08 | [17] |
| Maize bran | 68.80 ± 0.20 | 21.40 ± 0.04 | 47.20 ± 0.09 | |
| Oat bran | 61.20 ± 0.30 | 16.20 ± 0.05 | 47.20 ± 0.09 | |
| Pea | 10.40 ± 2.33 | 1.73 ± 0.26 | 20.30 ± 0.40 | [18] |
| Common bean | 8.55 ± 3.31 | 2.42 ± 0.74 | 19.90 ± 0.19 | |
| Chickpea | 9.88 ± 2.11 | 0.00 ± 0.00 | 13.90 ± 0.09 | |
| Lentil | 6.83 ± 2.42 | 1.44 ± 0.11 | 19.00 ± 1.27 | |
| Lyophilized plum skin | 38.98 ± 0.52 | 14.19 ± 0.66 | 24.81 ± 1.16 | [19] |
| Pomegranate (Acide) | 28.27 ± 0.90 | 1.16 ± 0.11 | 27.11 ± 0.65 | [20] |
| Actinidia deliciosa | 92.88 ± 0.75 | 32.85 ± 0.24 | 60.03 ± 0.50 | [21] |
| Kiwifruit pomace | 56.44 ± 1.02 | 6.90 ± 0.20 | 49.54 ± 0.86 | [22] |
| Okara | 65.96 ± 0.05 | 2.24 ± 0.10 | 63.72 ± 0.18 | [23,24] |
| Rose pomace | 46.30 ± 0.40 | 4.45 ± 0.81 | 41.12 ± 1.15 | [25] |
| Source of PDF | Method | Enhancement Activity | Improvement of Beneficial Bioactivity |
|---|---|---|---|
| Barley β-glucan [102,103] | β-glucanase hydrolysis | Improving viscosity | Increased yeast gas production and indirectly improve gluten network structure. |
| Wheat bran [104] | Cold plasma and enzymatic hydrolysis | Enhancing lipid-lowering ability | The combined modified SDF showed the best performance in glucose adsorption capacity, cholesterol adsorption capacity and antioxidant capacity. |
| Litchi Pomace [105,106] | Ultrasonic-enzymatic co-modification | Enriching beneficial bacteria | The modified SDF significantly stimulated the growth of probiotic bacterial species. |
| [107] | Extrusion and cellulase modification | Exposing more surface area and functional groups | The physicochemical properties and in vitro functional activity were enhanced, while the Pb2+ adsorption capacity was strengthened. |
| Rose pomace [25] | Enzymatic hydrolysis (EH) and ultrasound-assisted enzymatic hydrolysis (UEH) | Enhancing the looseness of the surface structure, resulting in a shale-like blocky appearance | Enhanced the capacity of oil-holding, swelling, cation-exchange, and cholesterol adsorption capacities. |
| Okara [108] | K. marxianu fermentation and β-glucosidase hydrolysis | Increasing porosity and reducing bulk density | Enhanced and prolonged the adsorption capacity for both glucose and cholesterol. |
| Orange pomace [109] | Extrusion processing | Increasing the content of soluble dietary fiber | Increased the uronic acid content in the SDF fraction. |
| Defatted rice bran [110] | γ-irradiation combined with enzymatic modification | Reducing particle size and increasing specific surface area | Enhanced the anti-digestive properties and probiotic activity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Shao, Y.; Fan, S.; Bai, J.; Zhu, L.; Zhu, Y.; Xiao, X. Advanced Modification Strategies of Plant-Sourced Dietary Fibers and Their Applications in Functional Foods. Foods 2025, 14, 2710. https://doi.org/10.3390/foods14152710
Zhao Y, Shao Y, Fan S, Bai J, Zhu L, Zhu Y, Xiao X. Advanced Modification Strategies of Plant-Sourced Dietary Fibers and Their Applications in Functional Foods. Foods. 2025; 14(15):2710. https://doi.org/10.3390/foods14152710
Chicago/Turabian StyleZhao, Yansheng, Ying Shao, Songtao Fan, Juan Bai, Lin Zhu, Ying Zhu, and Xiang Xiao. 2025. "Advanced Modification Strategies of Plant-Sourced Dietary Fibers and Their Applications in Functional Foods" Foods 14, no. 15: 2710. https://doi.org/10.3390/foods14152710
APA StyleZhao, Y., Shao, Y., Fan, S., Bai, J., Zhu, L., Zhu, Y., & Xiao, X. (2025). Advanced Modification Strategies of Plant-Sourced Dietary Fibers and Their Applications in Functional Foods. Foods, 14(15), 2710. https://doi.org/10.3390/foods14152710