
High-Vigor Rootstock Exacerbates Herbaceous Notes in Vitis vinifera L. cv. Cabernet Sauvignon Berries and Wines Under Humid Climates


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Site and Sampling
2.3. Berry Sampling and Winemaking
2.4. Quantification of Aromatic Volatiles by GC–MS/MS
2.5. Sensory Analysis
2.6. Total RNA Extraction and Real-Time qPCR Assay
2.7. Statistical Analyses
3. Results
3.1. Meteorological Conditions and Soil Indices
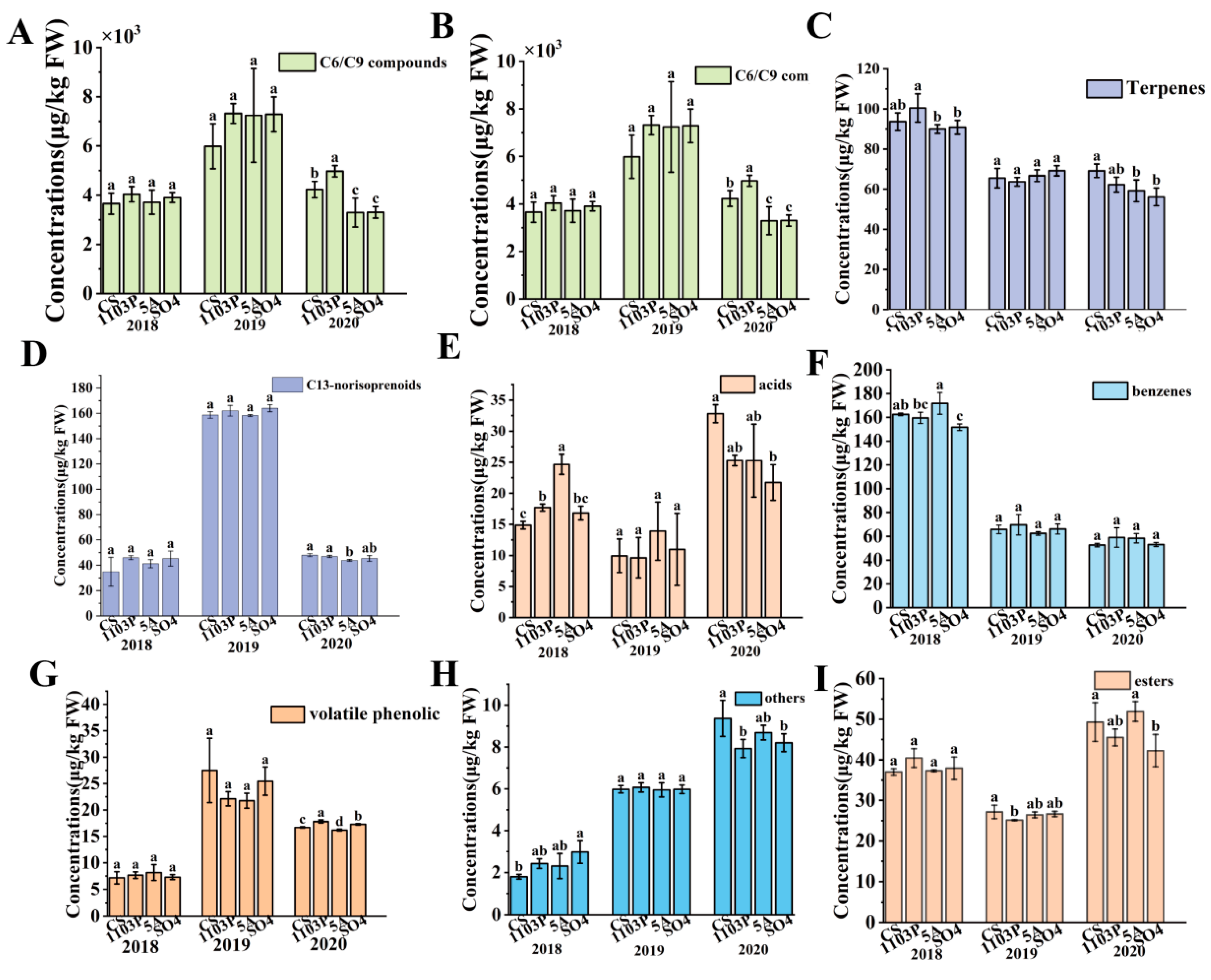
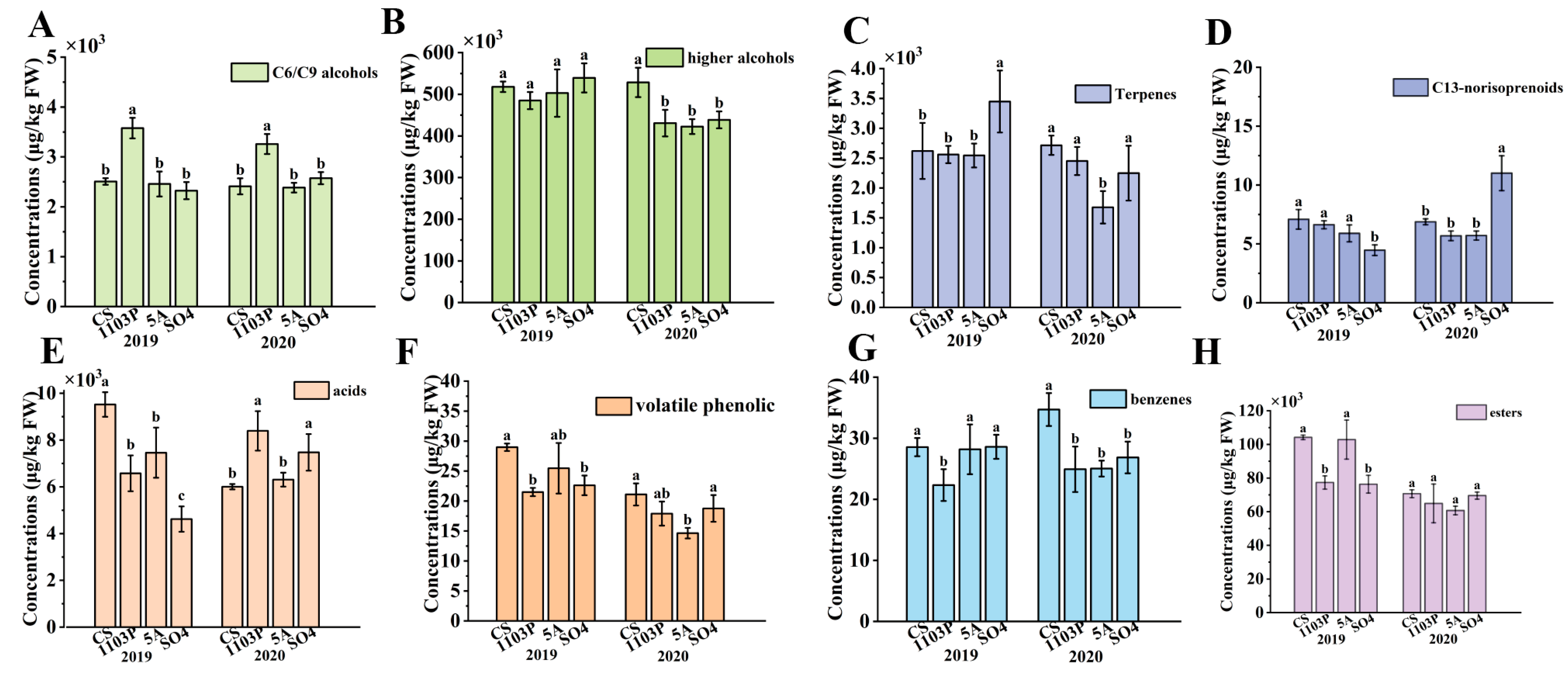
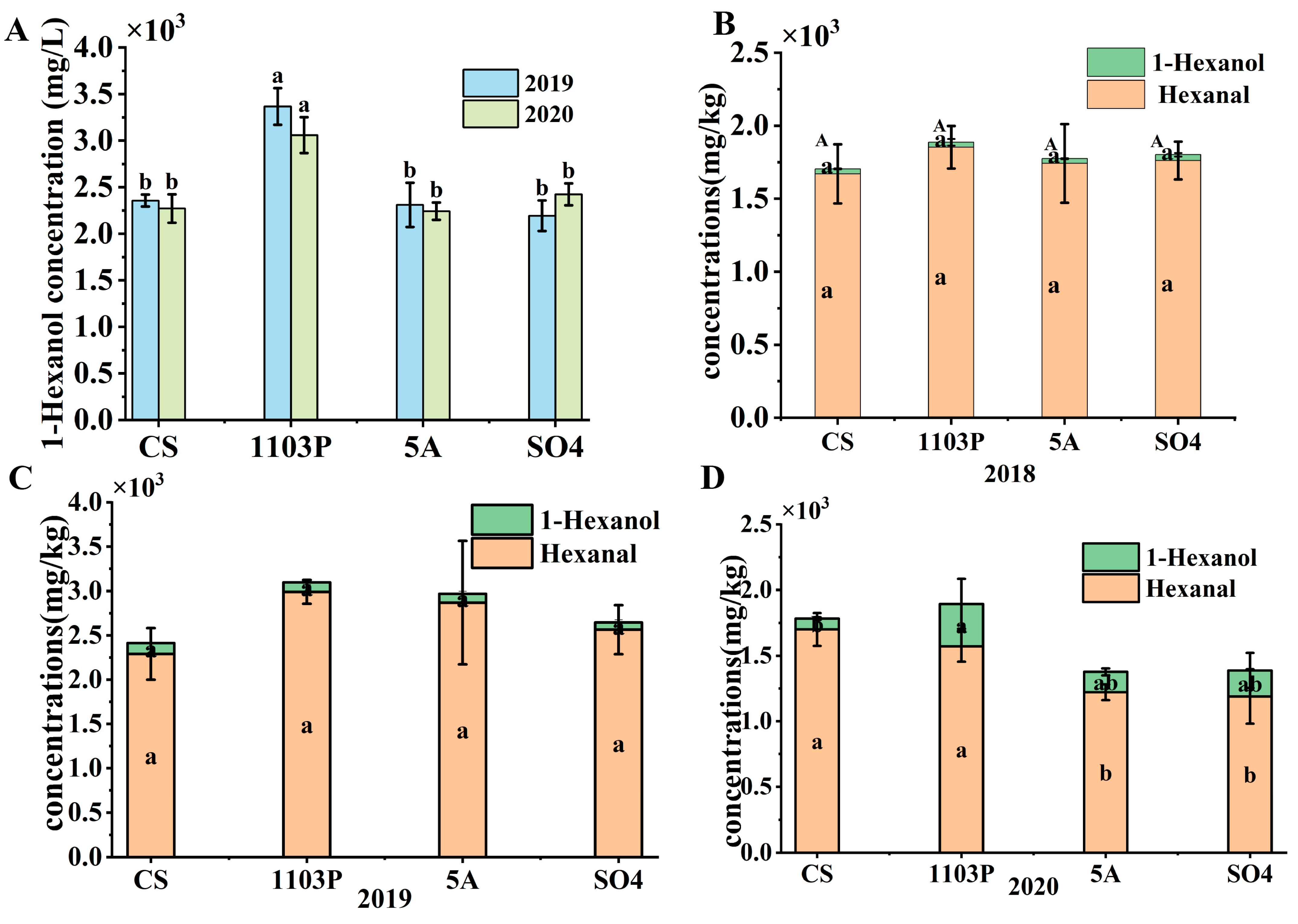
3.2. Effects of Rootstocks on Volatile Compound Profiles in Berries and in Wines
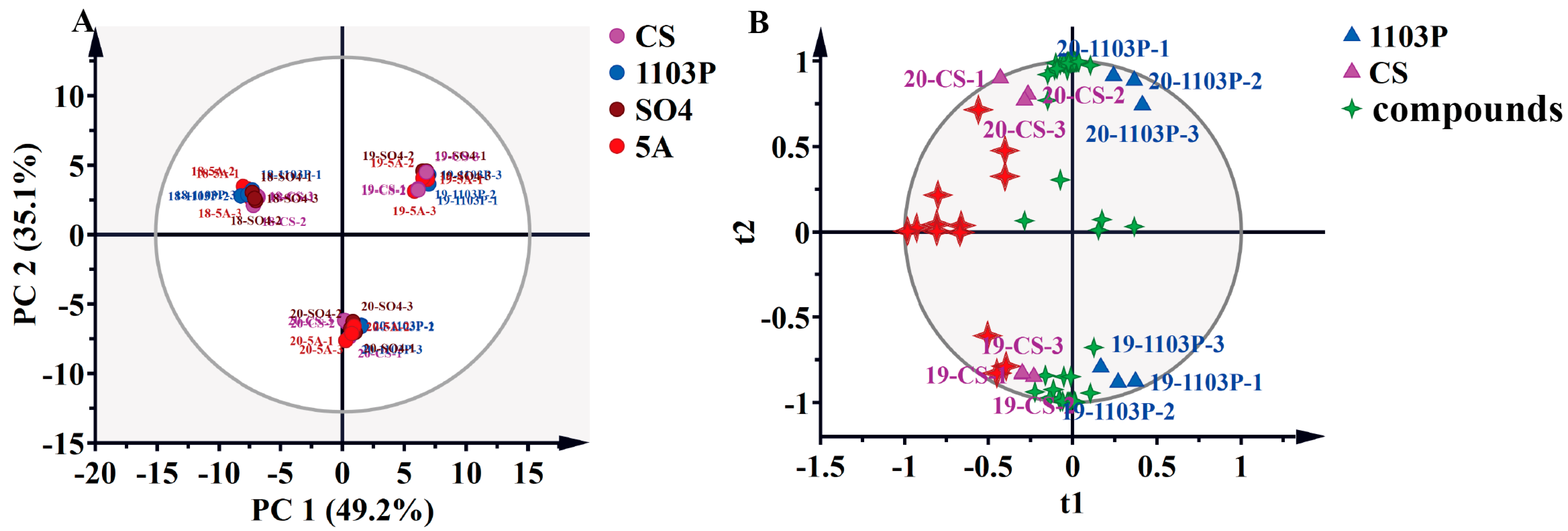
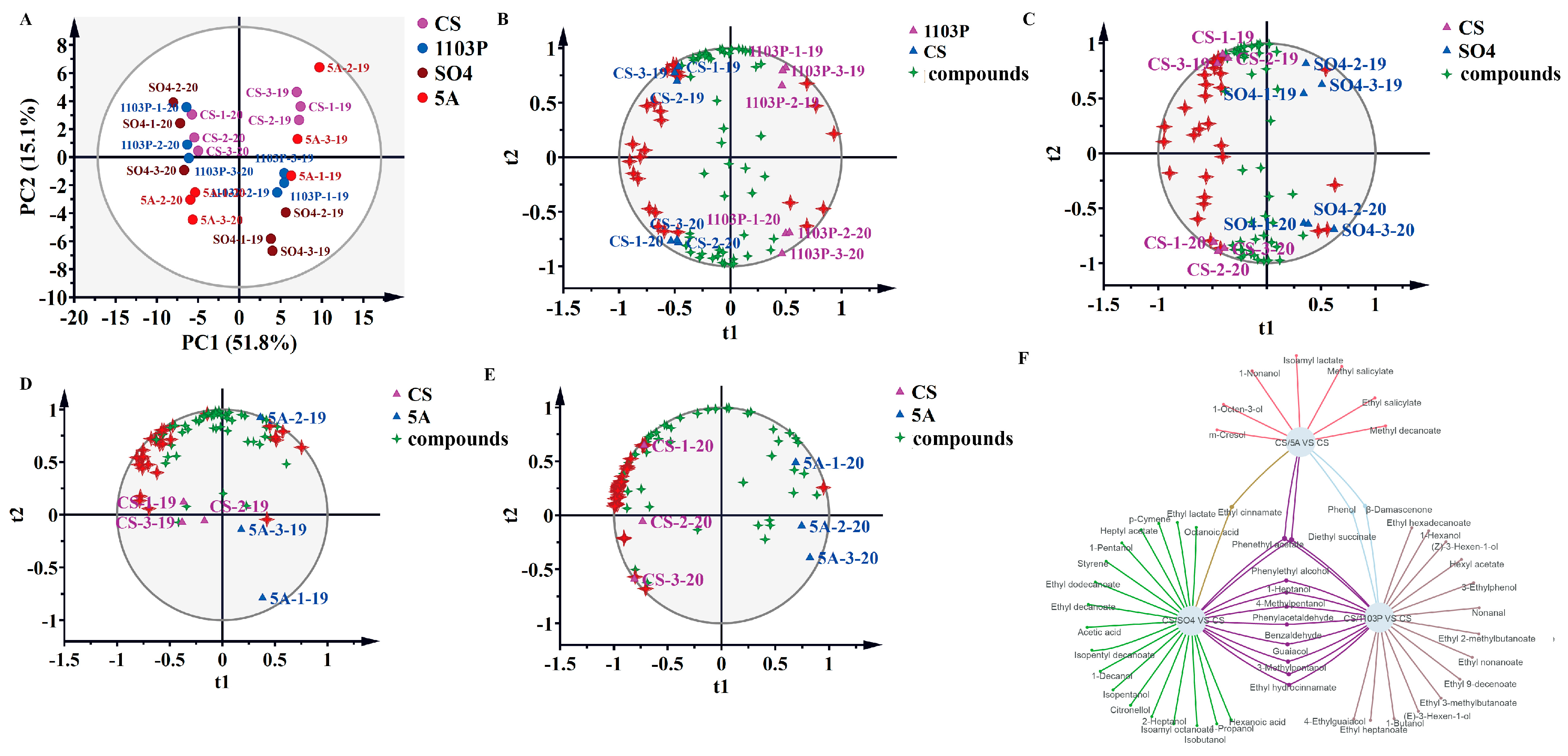
3.3. PCA and OPLS-DA of Compounds in Berries and in Wine
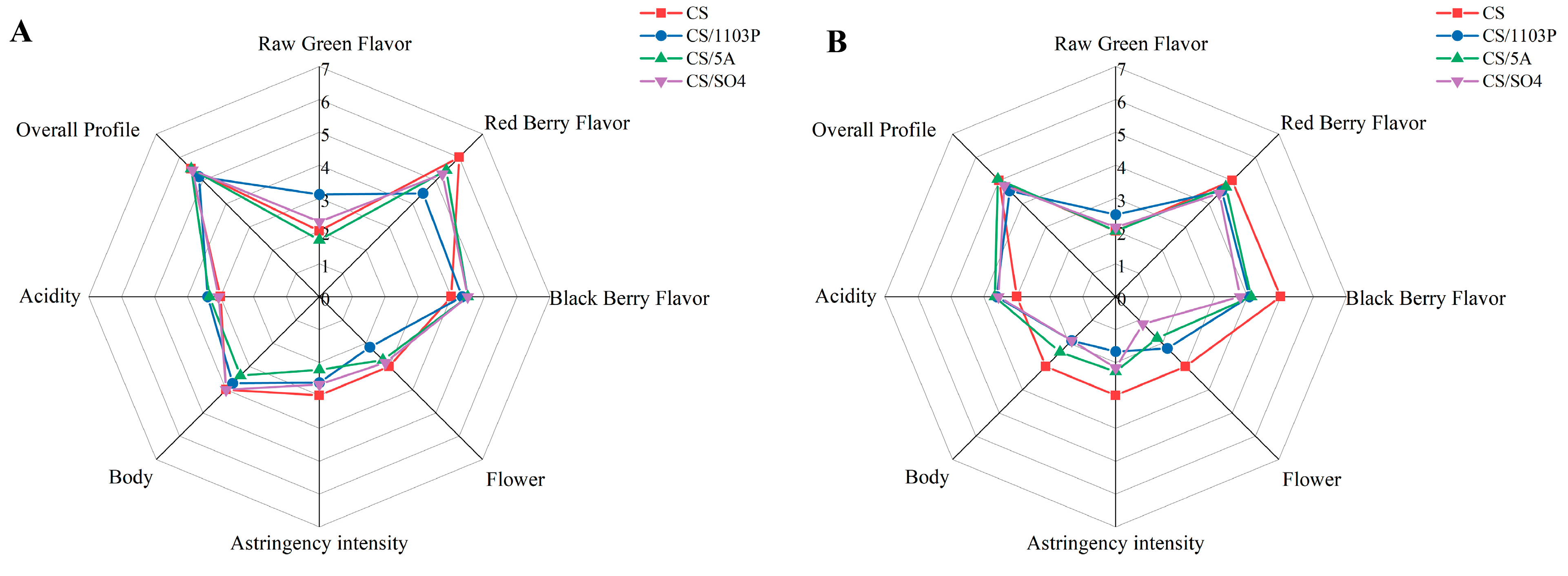
3.4. Volatile Compounds and Their OAVs in Wine and Sensory Tasting
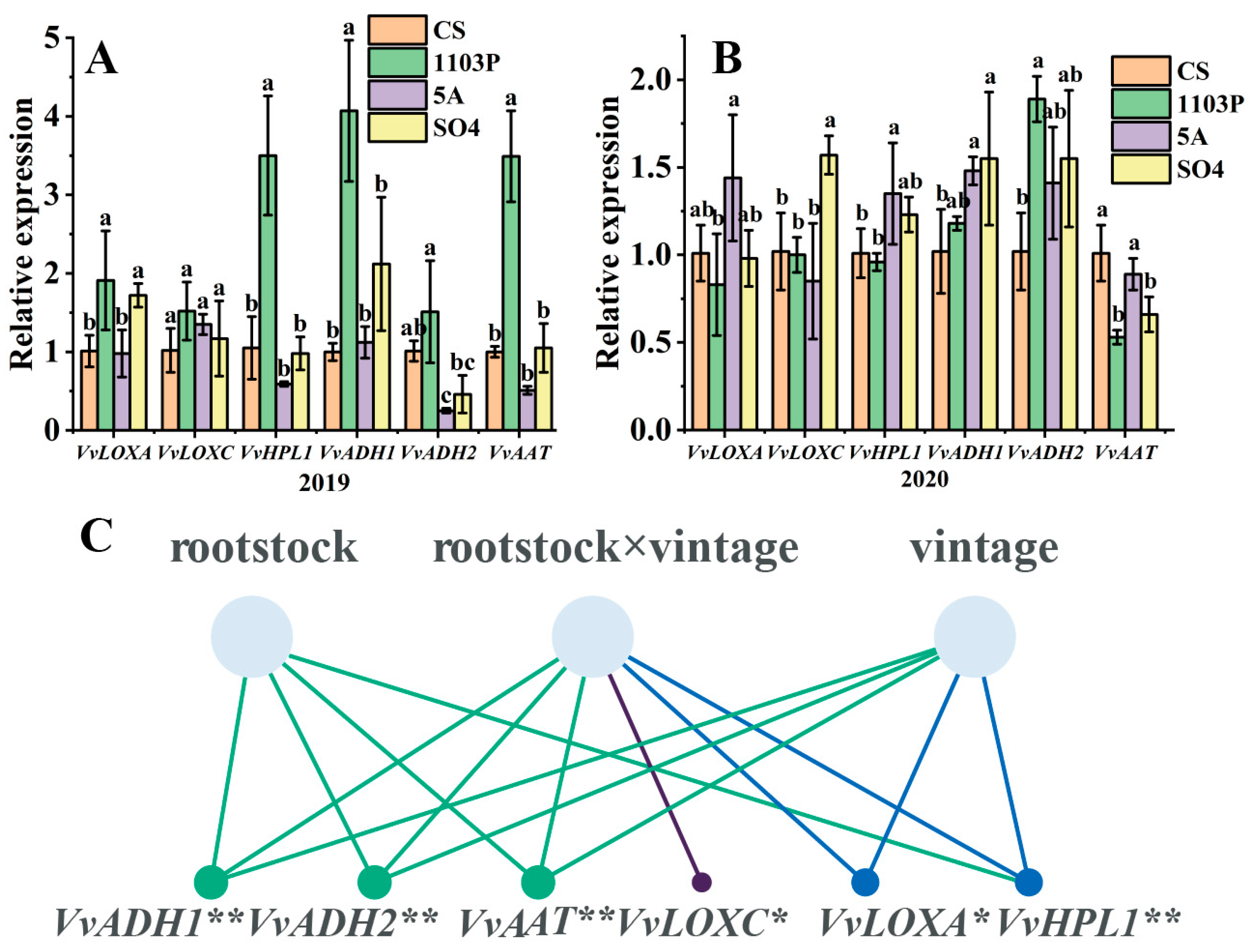
3.5. Expression of Genes in LOX-HPL Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delaunois, B.; Colby, T.; Belloy, N.; Conreux, A.; Harzen, A.; Baillieul, F.; Clément, C.; Schmidt, J.; Jeandet, P.; Cordelier, S. Large-scale proteomic analysis of the grapevine leaf apoplastic fluid reveals mainly stress-related proteins and cell wall modifying enzymes. BMC Plant Biol. 2013, 13, 24. [Google Scholar] [CrossRef]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agr. 2019, 99, 975–985. [Google Scholar] [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine aroma compounds in grapes: A critical review. Crit. Rev. Food Sci. 2015, 55, 202–218. [Google Scholar] [CrossRef]
- Parker, M.; Capone, D.L.; Francis, I.L.; Herderich, M.J. Aroma precursors in grapes and wine: Flavor release during wine production and consumption. J. Agr. Food Chem. 2017, 66, 2281–2286. [Google Scholar] [CrossRef]
- Chen, X.; Quek, S.Y. Free and glycosidically bound aroma compounds in fruit: Biosynthesis, transformation, and practical control. Crit. Rev. Food Sci. 2023, 63, 9052–9073. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.B.; Figueiredo, A. Biotic and Abiotic Stress Management in Grapevine: Recent Advances and Major Breakthroughs. Agronomy 2023, 13, 1584. [Google Scholar] [CrossRef]
- Granett, J.; Walker, M.A.; Kocsis, L.; Omer, A.D. Biology and management of grape phylloxera. Annu. Rev. Entomol. 2001, 46, 387–412. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Strever, A.; Myburgh, P.; Deloire, A. The interaction between rootstocks and cultivars (Vitis vinifera L.) to enhance drought tolerance in grapevine. Aust. J. Grape Wine R. 2014, 20, 1–14. [Google Scholar] [CrossRef]
- Koundouras, S.; Hatzidimitriou, E.; Karamolegkou, M.; Dimopoulou, E.; Kallithraka, S.; Tsialtas, J.T.; Zioziou, E.; Nikolaou, N.; Kotseridis, Y. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agr. Food Chem. 2009, 57, 7805–7813. [Google Scholar] [CrossRef]
- Ziegler, M.; Wegmann-Herr, P.; Schmarr, H.-G.; Gök, R.; Winterhalter, P.; Fischer, U. Impact of rootstock, clonal selection, and berry size of Vitis vinifera sp. Riesling on the formation of TDN, vitispiranes, and other volatile compounds. J. Agric. Food Chem. 2020, 68, 3834–3849. [Google Scholar] [CrossRef]
- Harbertson, J.F.; Keller, M. Rootstock Effects on Deficit-Irrigated Winegrapes in a Dry Climate: Grape and Wine Composition. Am. J. Enol. Viticult. 2012, 63, 40–48. [Google Scholar] [CrossRef]
- Zhang, L.; Marguerit, E.; Rossdeutsch, L.; Ollat, N.; Gambetta, G.A. The influence of grapevine rootstocks on scion growth and drought resistance. Theor. Exp. Plant Phys. 2016, 28, 143–157. [Google Scholar] [CrossRef]
- Francioli, D.; Strack, T.; Dries, L.; Voss-Fels, K.P.; Geilfus, C.M. Roots of resilience: Optimizing microbe-rootstock interactions to enhance vineyard productivity. Plants People Planet 2025, 7, 524–535. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.K.; Gao, X.T.; He, L.; Yang, X.H.; He, F.; Duan, C.Q.; Wang, J. Rootstock-Mediated Effects on Cabernet Sauvignon Performance: Vine Growth, Berry Ripening, Flavonoids, and Aromatic Profiles. Int. J. Mol. Sci. 2019, 20, 401. [Google Scholar] [CrossRef]
- Cheng, J.; Li, H.; Wang, W.; Duan, C.; Wang, J.; He, F. The influence of rootstocks on the scions’ aromatic profiles of Vitis vinifera L. cv. Chardonnay. Sci. Hortic. 2020, 272, 109517. [Google Scholar] [CrossRef]
- Vilanova, M.; Genisheva, Z.; Tubío, M.; Alvarez, K.; Lissarrague, J.R.; Oliveira, J.M. Rootstock effect on volatile composition of Albariño wines. Appl. Sci. 2021, 11, 2135. [Google Scholar] [CrossRef]
- Carrasco-Quiroz, M.; Martínez-Gil, A.M.; Gutiérrez-Gamboa, G.; Moreno-Simunovic, Y. Effect of rootstocks on volatile composition of Merlot wines. J. Sci. Food Agr. 2020, 100, 3517–3524. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Pichersky, E.; Schaub, A.; Hansel, A.; Gershenzon, J. Characterization of a BAHD acyltransferase responsible for producing the green leaf volatile (Z)-3-hexen-1-yl acetate in Arabidopsis thaliana. Plant J. 2007, 49, 194–207. [Google Scholar] [CrossRef]
- Tesnière, C.; Verries, C. Molecular cloning and expression of cDNAs encoding alcohol dehydrogenases from Vitis vinifera L. during berry development. Plant Sci. 2000, 157, 77–88. [Google Scholar] [CrossRef]
- Han, X.; Lu, H.-C.; Wang, Y.; Gao, X.-T.; Li, H.-Q.; Tian, M.-B.; Shi, N.; Li, M.-Y.; Yang, X.-L.; He, F. Region, vintage, and grape maturity co-shaped the ionomic signatures of the Cabernet Sauvignon wines. Food Res. Int. 2023, 163, 112165. [Google Scholar] [CrossRef]
- Brun, L.; Maillet, J.; Hinsinger, P.; Pepin, M. Evaluation of copper availability to plants in copper-contaminated vineyard soils. Environ. Pollut. 2001, 111, 293–302. [Google Scholar] [CrossRef]
- Sáez-Plaza, P.; Navas, M.J.; Wybraniec, S.; Michałowski, T.; Asuero, A.G. An overview of the Kjeldahl method of nitrogen determination. Part II. Sample preparation, working scale, instrumental finish, and quality control. Crit. Rev. Anal. Chem. 2013, 43, 224–272. [Google Scholar] [CrossRef]
- Buondonno, A.; Rashad, A.A.; Coppola, E. Comparing tests for soil fertility. II. The hydrogen peroxide/sulfuric acid treatment as an alternative to the copper/selenium catalyzed digestion process for routine determination of soil nitrogen-kjeldahl. Commun. Soil Sci. Plan. 1995, 26, 1607–1619. [Google Scholar] [CrossRef]
- Górecka, H.; Chojnacka, K.; Górecki, H. The application of ICP-MS and ICP-OES in determination of micronutrients in wood ashes used as soil conditioners. Talanta 2006, 70, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, H.-Q.; Gao, X.-T.; Lu, H.-C.; Peng, W.-T.; Chen, W.; Li, S.-P.; Duan, C.-Q.; Wang, J. Influence of attenuated reflected solar radiation from the vineyard floor on volatile compounds in Cabernet Sauvignon grapes and wines of the north foot of Mt. Tianshan. Food Res. Int. 2020, 137, 109688. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.; Qi, M.; Li, S.; Shi, Y.; Pan, Q.; Cheng, C.; Yang, W.; Duan, C. The influence of polyphenol supplementation on ester formation during red wine alcoholic fermentation. Food Chem. 2022, 377, 131961. [Google Scholar] [CrossRef]
- Francis, I.; Newton, J. Determining wine aroma from compositional data. Aust. J. Grape Wine R. 2005, 11, 114–126. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Chen, H.; He, C.; Sun, L.; Zhang, X.; Xu, Z.; Liu, H. Integrated omics profiles for exploring the potential mechanism underlying aroma formation in the terpenoid-rich aromatic plant Opisthopappus taihangensis and the bioactivity of its leaf essential oil. Agric. Commun. 2024, 2, 100061. [Google Scholar] [CrossRef]
- Qian, X.; Xu, X.-Q.; Yu, K.-J.; Zhu, B.-Q.; Lan, Y.-B.; Duan, C.-Q.; Pan, Q.-H. Varietal dependence of GLVs accumulation and LOX-HPL pathway gene expression in four Vitis vinifera wine grapes. Int. J. Mol. Sci. 2016, 17, 1924. [Google Scholar] [CrossRef]
- Tan, L.; Lu, L.; Sun, W.; Zhang, X.; Liu, Y.; Xiang, Y.; Yan, H. Identification and validation of qRT-PCR reference genes for analyzing grape infection with gray mold. BMC Genom. 2024, 25, 997. [Google Scholar] [CrossRef]
- Podolyan, A.; White, J.; Jordan, B.; Winefield, C. Identification of the lipoxygenase gene family from Vitis vinifera and biochemical characterisation of two 13-lipoxygenases expressed in grape berries of Sauvignon Blanc. Funct. Plant Biol. 2010, 37, 767–784. [Google Scholar] [CrossRef]
- Xu, X.-Q.; Cheng, G.; Duan, L.-L.; Jiang, R.; Pan, Q.-H.; Duan, C.-Q.; Wang, J. Effect of training systems on fatty acids and their derived volatiles in Cabernet Sauvignon grapes and wines of the north foot of Mt.Tianshan. Food Chem. 2015, 181, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Tesniere, C.; Davies, C.; Sreekantan, L.; Bogs, J.; Thomas, M.; Torregrosa, L. Analysis of the transcript levels of VvAdh1, VvAdh2 and VvGrip4, three genes highly expressed during Vitis vinifera L. berry development. Vitis 2006, 45, 75. [Google Scholar]
- Lee, K.R.; Kim, S.H.; Go, Y.S.; Jung, S.M.; Roh, K.H.; Kim, J.B.; Suh, M.C.; Lee, S.; Kim, H.U. Molecular cloning and functional analysis of two FAD2 genes from American grape (Vitis labrusca L.). Gene 2012, 509, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Weindorf, D.C.; Wang, D.; Chakraborty, S. Characterizing soils via portable X-ray fluorescence spectrometer: 4. Cation exchange capacity (CEC). Geoderma 2015, 239, 130–134. [Google Scholar] [CrossRef]
- Yu, R.; Brillante, L.; Martínez-Lüscher, J.; Kurtural, S.K. Spatial variability of soil and plant water status and their cascading effects on grapevine physiology are linked to berry and wine chemistry. Front. Plant Sci. 2020, 11, 790. [Google Scholar] [CrossRef]
- Noguerol-Pato, R.; González-Álvarez, M.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J. Aroma profile of Garnacha Tintorera-based sweet wines by chromatographic and sensorial analyses. Food Chem. 2012, 134, 2313–2325. [Google Scholar] [CrossRef]
- Gómez-Míguez, M.J.; Cacho, J.F.; Ferreira, V.; Vicario, I.M.; Heredia, F.J. Volatile components of Zalema white wines. Food Chem. 2007, 100, 1464–1473. [Google Scholar] [CrossRef]
- Escudero, A.; Campo, E.; Fariña, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agr. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef]
- Pineau, B.; Barbe, J.-C.; Van Leeuwen, C.; Dubourdieu, D. Examples of perceptive interactions involved in specific “red-” and “black-berry” aromas in red wines. J. Agr. Food Chem. 2009, 57, 3702–3708. [Google Scholar] [CrossRef]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agr. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Gomez, E.; Martinez, A.; Laencina, J. Changes in volatile compounds during maturation of some grape varieties. J. Sci. Food Agr. 1995, 67, 229–233. [Google Scholar] [CrossRef]
- Ou, C.; Du, X.; Shellie, K.; Ross, C.; Qian, M.C. Volatile compounds and sensory attributes of wine from cv. Merlot (Vitis vinifera L.) grown under differential levels of water deficit with or without a kaolin-based, foliar reflectant particle film. J. Agr. Food Chem. 2010, 58, 12890–12898. [Google Scholar] [CrossRef]
- Romero, P.; Botía, P.; del Amor, F.M.; Gil-Muñoz, R.; Flores, P.; Navarro, J.M. Interactive effects of the rootstock and the deficit irrigation technique on wine composition, nutraceutical potential, aromatic profile, and sensory attributes under semiarid and water limiting conditions. Agr. Water Manag. 2019, 225, 105733. [Google Scholar] [CrossRef]
- Romano, P.; Braschi, G.; Siesto, G.; Patrignani, F.; Lanciotti, R. Role of yeasts on the sensory component of wines. Foods 2022, 11, 1921. [Google Scholar] [CrossRef]
- Li, C.; Chen, H.; Li, Y.; Du, T.; Jia, J.; Xi, Z. The Expression of Aroma Components and Related Genes in Merlot and Marselan Scion–Rootstock Grape and Wine. Foods 2022, 11, 2777. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yu, M.; Yao, Y.; Zhai, H.; Tang, M.; Gao, Z.; Du, Y. Effect of Seawater Irrigation on the Sugars, Organic Acids, and Volatiles in ‘Reliance’ Grape. Horticulturae 2022, 8, 248. [Google Scholar] [CrossRef]
- Han, X.; Wang, Y.; Lu, H.-C.; Yang, H.-Y.; Li, H.-Q.; Gao, X.-T.; Pei, X.-X.; He, F.; Duan, C.-Q.; Wang, J. The combined influence of rootstock and vintage climate on the grape and wine flavonoids of Vitis vinifera L. cv. Cabernet Sauvignon in eastern China. Front. Plant Sci. 2022, 13, 978497. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Lu, H.; Wang, X.; Wang, Y.; Chen, W.; Pei, X.; He, F.; Duan, C.; Wang, J. High-Vigor Rootstock Exacerbates Herbaceous Notes in Vitis vinifera L. cv. Cabernet Sauvignon Berries and Wines Under Humid Climates. Foods 2025, 14, 2695. https://doi.org/10.3390/foods14152695
Han X, Lu H, Wang X, Wang Y, Chen W, Pei X, He F, Duan C, Wang J. High-Vigor Rootstock Exacerbates Herbaceous Notes in Vitis vinifera L. cv. Cabernet Sauvignon Berries and Wines Under Humid Climates. Foods. 2025; 14(15):2695. https://doi.org/10.3390/foods14152695
Chicago/Turabian StyleHan, Xiao, Haocheng Lu, Xia Wang, Yu Wang, Weikai Chen, Xuanxuan Pei, Fei He, Changqing Duan, and Jun Wang. 2025. "High-Vigor Rootstock Exacerbates Herbaceous Notes in Vitis vinifera L. cv. Cabernet Sauvignon Berries and Wines Under Humid Climates" Foods 14, no. 15: 2695. https://doi.org/10.3390/foods14152695
APA StyleHan, X., Lu, H., Wang, X., Wang, Y., Chen, W., Pei, X., He, F., Duan, C., & Wang, J. (2025). High-Vigor Rootstock Exacerbates Herbaceous Notes in Vitis vinifera L. cv. Cabernet Sauvignon Berries and Wines Under Humid Climates. Foods, 14(15), 2695. https://doi.org/10.3390/foods14152695