
Isolation and Characterization of a Cold-Adapted Bacteriophage for Biocontrol of Vibrio parahaemolyticus in Seafood

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Isolation, Purification, and Propagation of Bacteriophages
2.3. Transmission Electron Microscopy (TEM)
2.4. Phage Lytic Range
2.5. Multiplicity of Infection (MOI)
2.6. One-Step Growth Assay
2.7. Stability Tests of Bacteriophage XY75
2.8. Extraction of DNA
2.9. Genome-Wide Analysis and Phylogenetic Analysis
2.10. Prevention and Control Applications for Salmon
3. Results
3.1. Isolation and Morphological Analysis of Phage XY75
3.2. Host Range of XY75
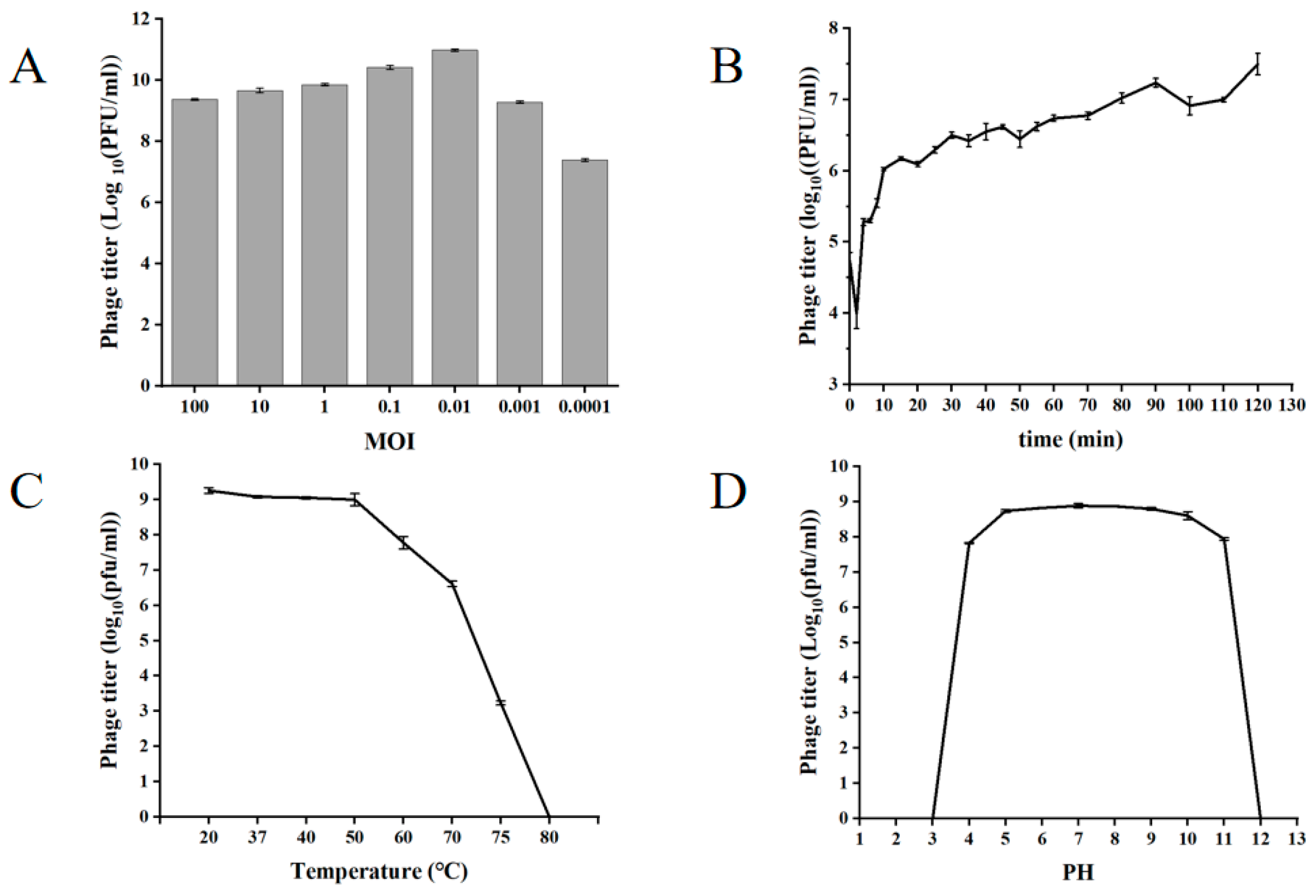
3.3. Biological Characteristics of XY75
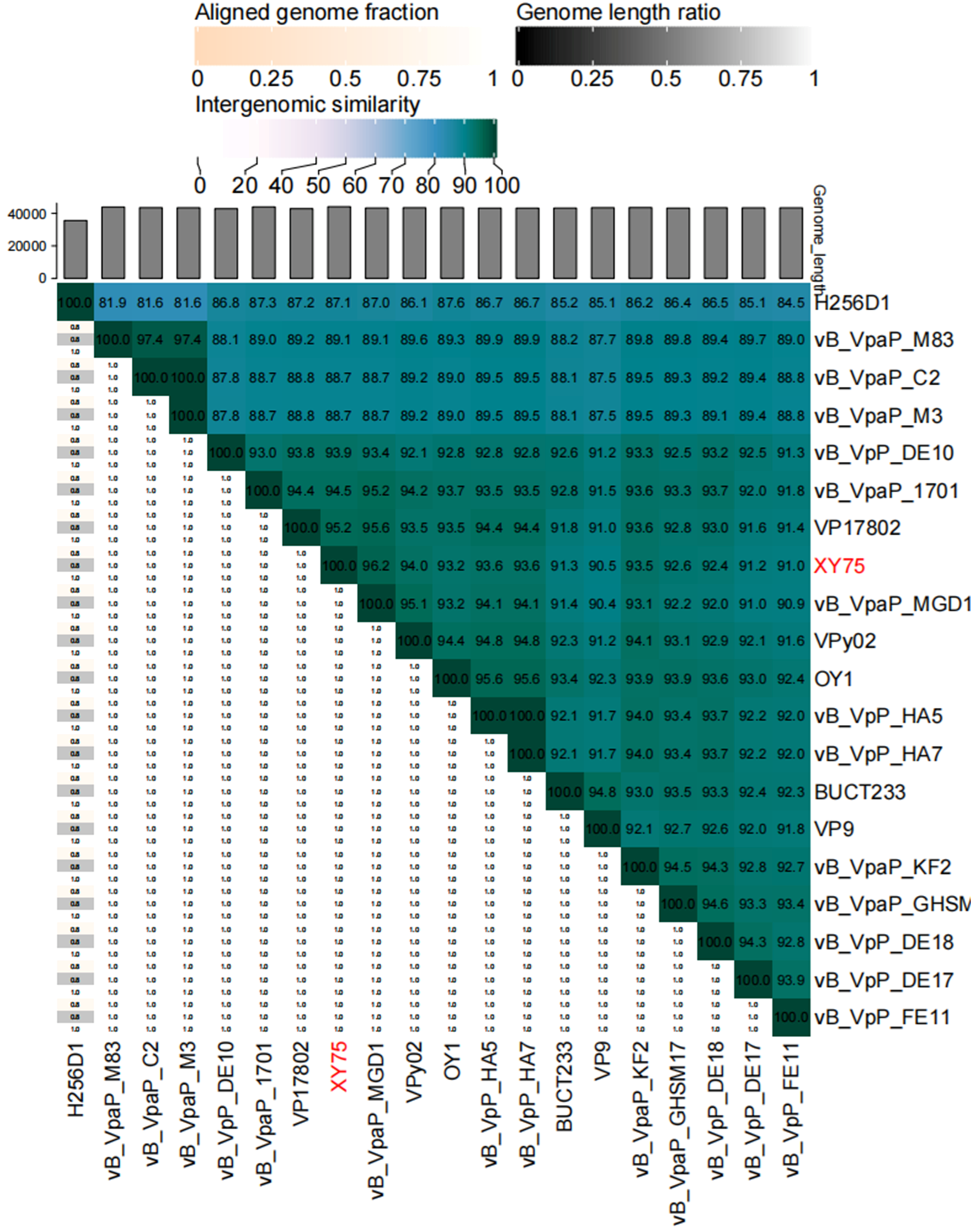
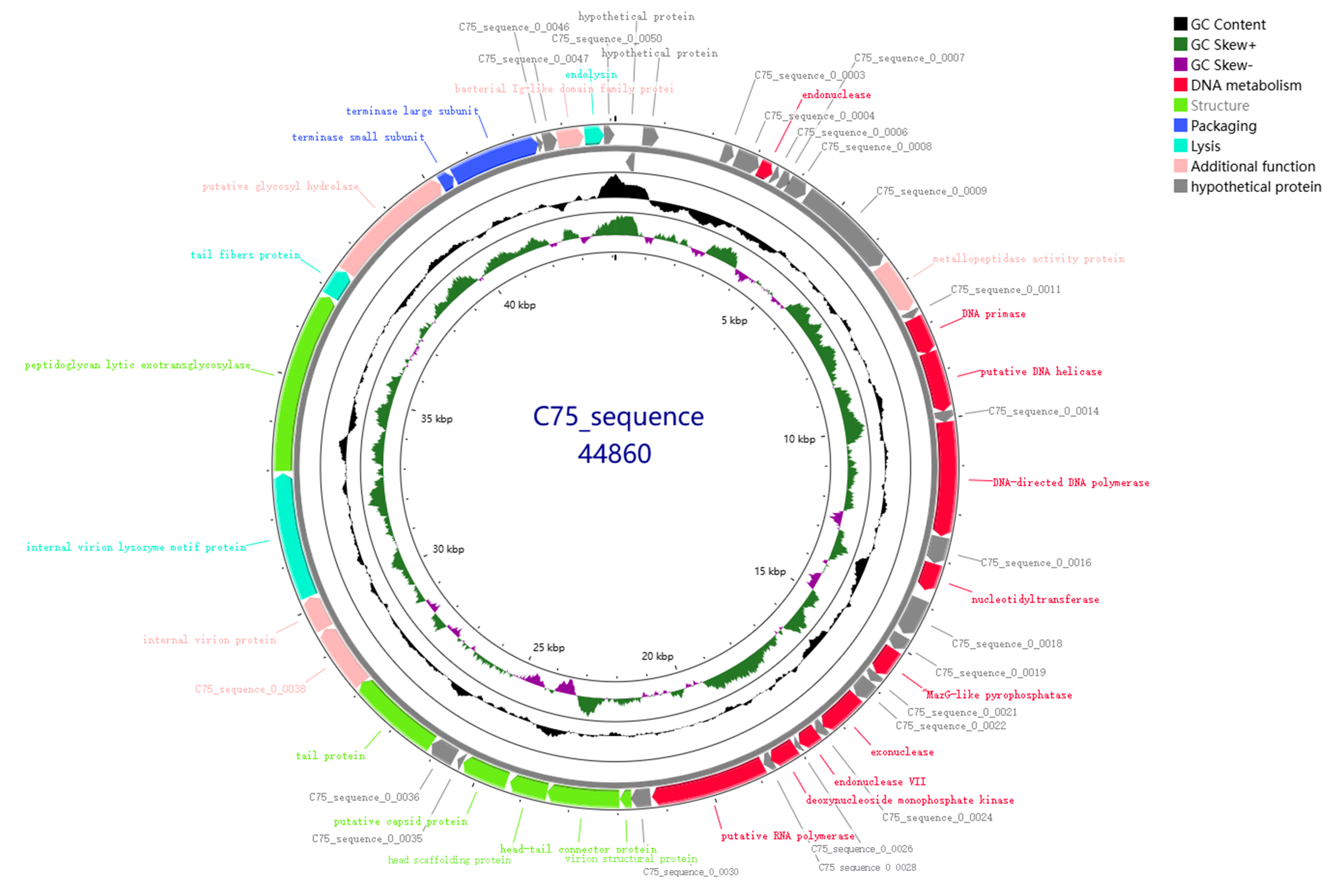
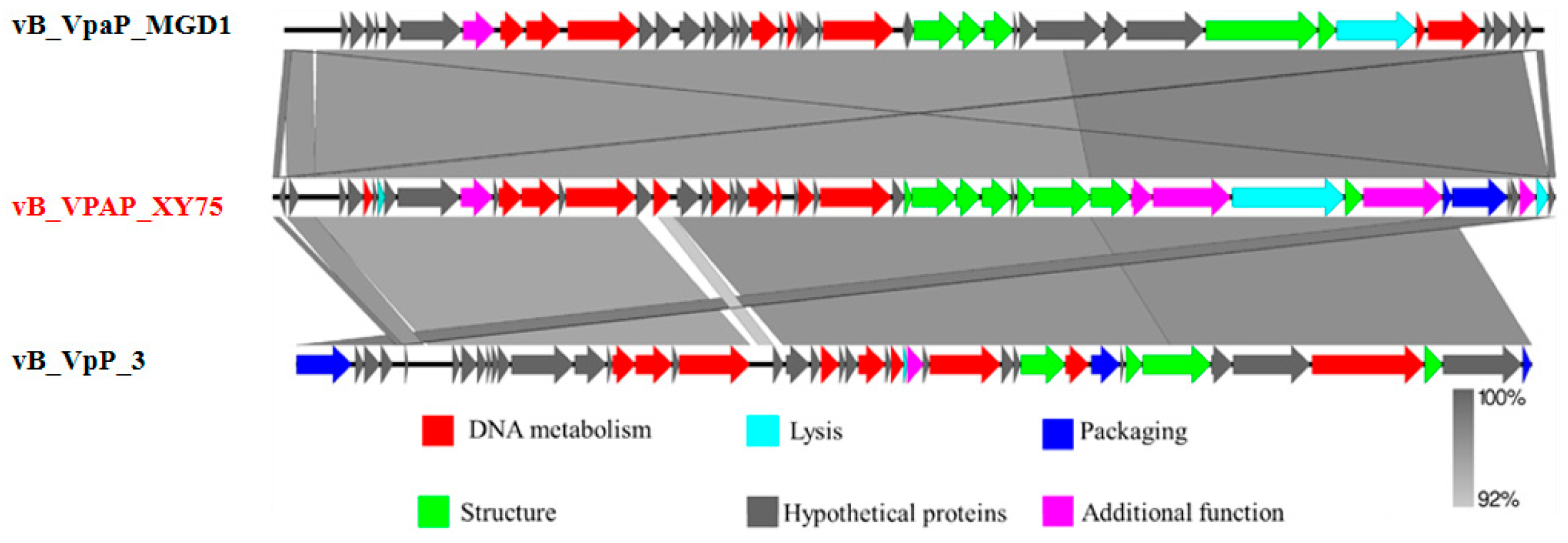
3.4. Genome Sequence Analysis
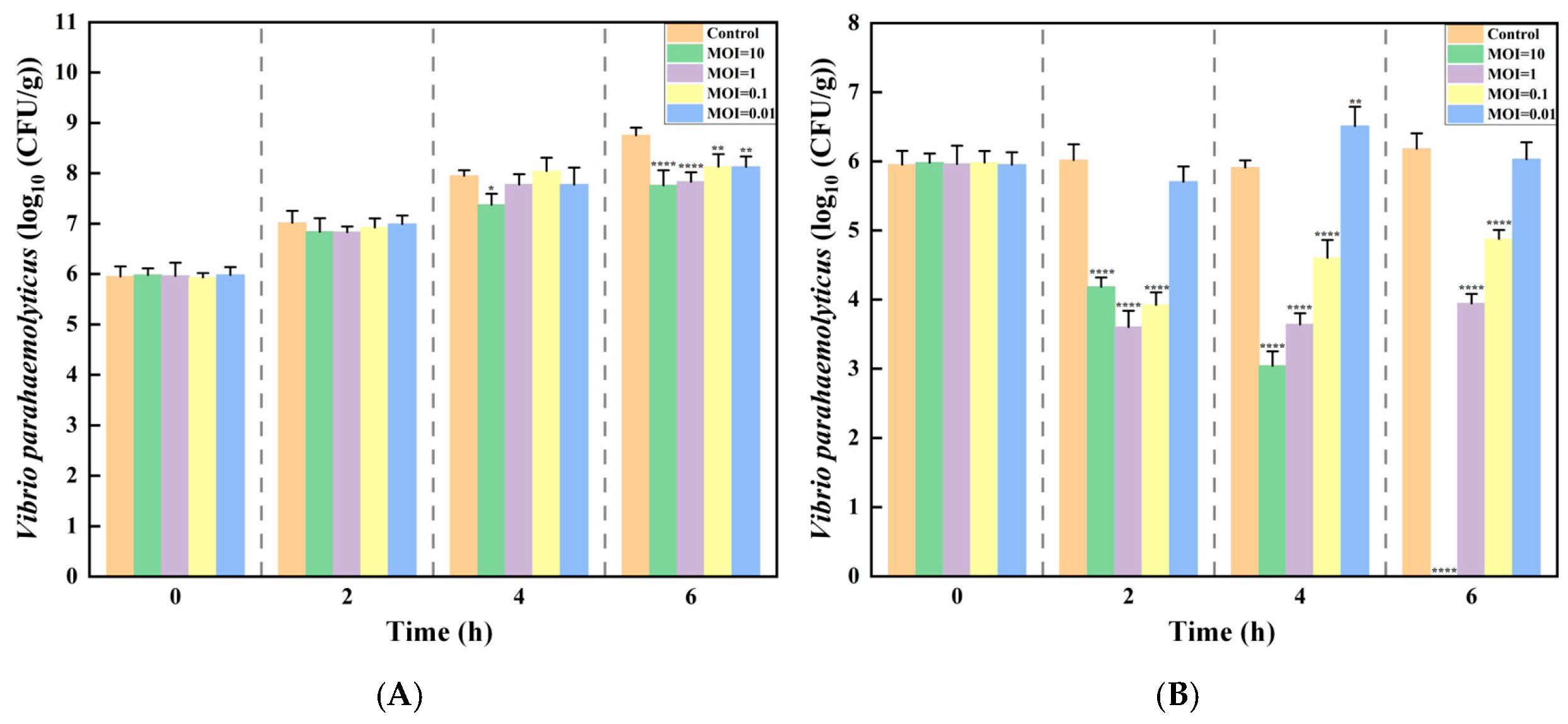
3.5. Food Applications of Phage XY75
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lian, X.Q.; Liu, G.D.; Huang, M.F.; Fan, Q.H.; Lin, Z.D. Antimicrobial Resistance, Virulence Factors and Phylogenetic Profiles of Vibrio parahaemolyticus in the Eastern Coast of Shenzhen. Front. Microbiol. 2024, 15, 1452942. [Google Scholar] [CrossRef]
- Shinoda, S. Sixty Years from the Discovery of Vibrio parahaemolyticus and Some Recollections. Biocontrol Sci. 2011, 16, 129–137. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Li, Y.; Chen, M.; Zeng, H.; Lei, T.; Zhang, J.; Ding, Y.; Wang, J.; Wu, S.; Ye, Q.; et al. A Database for Risk Assessment and Comparative Genomic Analysis of Foodborne Vibrio parahaemolyticus in China. Sci. Data 2020, 7, 321. [Google Scholar] [CrossRef] [PubMed]
- Chimalapati, S.; Lafrance, A.E.; Chen, L.; Orth, K. Vibrio parahaemolyticus: Basic Techniques for Growth, Genetic Manipulation, and Analysis of Virulence Factors. Curr. Protoc. Microbiol. 2020, 59, e131. [Google Scholar] [CrossRef] [PubMed]
- Sorée, M.; Lozach, S.; Kéomurdjian, N.; Richard, D.; Hughes, A.; Delbarre-Ladrat, C.; Verrez-Bagnis, V.; Rincé, A.; Passerini, D.; Ritchie, J.M.; et al. Virulence Phenotypes Differ between Toxigenic Vibrio parahaemolyticus Isolated from Western Coasts of Europe. Microbiol. Res. 2024, 285, 127744. [Google Scholar] [CrossRef]
- Hong, X.; Lu, L.; Xu, D. Progress in Research on Acute Hepatopancreatic Necrosis Disease (AHPND). Aquacult. Int. 2016, 24, 577–593. [Google Scholar] [CrossRef]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial Use in Aquaculture Re-Examined: Its Relevance to Antimicrobial Resistance and to Animal and Human Health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef]
- Yano, Y.; Hamano, K.; Satomi, M.; Tsutsui, I.; Ban, M.; Aue-umneoy, D. Prevalence and Antimicrobial Susceptibility of Vibrio Species Related to Food Safety Isolated from Shrimp Cultured at Inland Ponds in Thailand. Food Control 2014, 38, 30–36. [Google Scholar] [CrossRef]
- Oniciuc, E.-A.; Likotrafiti, E.; Alvarez-Molina, A.; Prieto, M.; López, M.; Alvarez-Ordóñez, A. Food Processing as a Risk Factor for Antimicrobial Resistance Spread along the Food Chain. Curr. Opin. Food Sci. 2019, 30, 21–26. [Google Scholar] [CrossRef]
- Zaafrane, S.; Maatouk, K.; Alibi, S.; Ben Mansour, H. Occurrence and Antibiotic Resistance of Vibrio parahaemolyticus Isolated from the Tunisian Coastal Seawater. J. Water Health 2022, 20, 369–384. [Google Scholar] [CrossRef]
- Voigt, A.M.; Zacharias, N.; Timm, C.; Wasser, F.; Sib, E.; Skutlarek, D.; Parcina, M.; Schmithausen, R.M.; Schwartz, T.; Hembach, N.; et al. Association between Antibiotic Residues, Antibiotic Resistant Bacteria and Antibiotic Resistance Genes in Anthropogenic Wastewater—An Evaluation of Clinical Influences. Chemosphere 2020, 241, 125032. [Google Scholar] [CrossRef]
- Brüssow, H. Phage Therapy: The Escherichia coli Experience. Microbiology 2005, 151, 2133–2140. [Google Scholar] [CrossRef]
- Gibb, B.; Hyman, P.; Schneider, C.L. The Many Applications of Engineered Bacteriophages—An Overview. Pharmaceuticals 2021, 14, 634. [Google Scholar] [CrossRef]
- Myelnikov, D. An Alternative Cure: The Adoption and Survival of Bacteriophage Therapy in the USSR, 1922–1955. J. Hist. Med. Allied Sci. 2018, 73, 385–411. [Google Scholar] [CrossRef]
- Chen, C.; Yuan, L.; Zhou, W.; Mgomi, F.C.; Zhang, Y.; Wang, Y.; Zheng, X.; Hu, Q.; Gao, L.; Rao, S.; et al. Isolation and Genomic Characterization of P.A-5, a Novel Virulent Bacteriophage against Enterobacter hormaechei. Microb. Pathog. 2021, 152, 104767. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.D.; Parks, A.; Abuladze, T.; Li, M.; Woolston, J.; Magnone, J.; Senecal, A.; Kropinski, A.M.; Sulakvelidze, A. Bacteriophage Cocktail Significantly Reduces Escherichia Coli O157: H7 Contamination of Lettuce and Beef, but Does Not Protect against Recontamination. Bacteriophage 2012, 2, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Sadekuzzaman, M.; Ha, S.-D. Reduction of Listeria monocytogenes on Chicken Breasts by Combined Treatment with UV-C Light and Bacteriophage ListShield. LWT 2017, 86, 193–200. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, Y.D.; Nan, Y.; Stanford, K.; Holley, R.; McAllister, T.; Narváez-Bravo, C. SalmoFreshTM Effectiveness in Controlling Salmonella on Romaine Lettuce, Mung Bean Sprouts and Seeds. Int. J. Food Microbiol. 2019, 305, 108250. [Google Scholar] [CrossRef]
- Cheng, B.; Zhang, D.; Wang, T.; Chen, H.; Wang, Y.; Wang, Y.; Li, Z.; Ling, N.; Ye, Y. Characterization of the Broad-Spectrum Phage vB_CsaM_CBT2 and Its Inhibition of Multidrug-Resistant Cronobacter sakazakii in Powered Infant Formula. Food Control 2024, 158, 110262. [Google Scholar] [CrossRef]
- Vo Thi Tu, A.; Pham-Khanh, N.H.; Nguyen, S.H.; Sunahara, H.; Dai Thi Xuan, T.; Kamei, K. Isolation, Characterization, and Complete Genome Sequence of Vibrio Phage KIT04, a Novel Lytic Phage of the Subfamily Ermolyevavirinae. Virology 2023, 579, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Peña-Rodríguez, A.; Ramírez-Sánchez, I.; Leptihn, S.; Quiroz-Guzmán, E. A Novel Vibrio phage vB_Vp_PvVp04 against Pathogenic Vibrio parahaemolyticus and Its Formulation for Inclusion in Shrimp Feed. Aquac. Int. 2025, 33, 117. [Google Scholar] [CrossRef]
- Yilmaz, N.; Yildirim, Z.; Sahin, T.S.; Karaoğlan, M. Isolation and Characterization of Lytic Bacteriophages Specific to Foodborne Pathogenic Pseudomonas aeruginosa and Use as a Biopreservative in Chicken Meat. Food Bioprocess Technol. 2024, 18, 2545–2560. [Google Scholar] [CrossRef]
- Azari, R.; Yousefi, M.H.; Taghipour, Z.; Wagemans, J.; Lavigne, R.; Hosseinzadeh, S.; Mazloomi, S.M.; Vallino, M.; Khalatbari-Limaki, S.; Berizi, E. Application of the Lytic Bacteriophage Rostam to Control Salmonella enteritidis in Eggs. Int. J. Food Microbiol. 2023, 389, 110097. [Google Scholar] [CrossRef]
- Zeng, H.; Li, C.; Luo, D.; Zhang, J.; Ding, Y.; Chen, M.; Yang, X.; Lei, T.; Wu, S.; Ye, Q.; et al. Novel Phage vB_CtuP_B1 for Controlling Cronobacter malonaticus and Cronobacter turicensis in Ready-to-Eat Lettuce and Powered Infant Formula. Food Res. Int. 2021, 143, 110255. [Google Scholar] [CrossRef]
- Kot, W. Genome Sequencing of dsDNA-Containing Bacteriophages Directly from a Single Plaque. Methods Mol. Biol. 2017, 1681, 179–184. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P.; Grant, J.R.; Domselaar, G.V. Visualizing and Comparing Circular Genomes Using the CGView Family of Tools. Brief. Bioinform. 2017, 20, 1576–1582. [Google Scholar] [CrossRef]
- Lomelí-Ortega, C.O.; Martínez-Díaz, S.F. Phage Therapy against Vibrio Parahaemolyticus Infection in the Whiteleg Shrimp (Litopenaeus vannamei) Larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Stalin, N.; Srinivasan, P. Characterization of Vibrio parahaemolyticus and Its Specific Phage from Shrimp Pond in Palk Strait, South East Coast of India. Biologicals 2016, 44, 526–533. [Google Scholar] [CrossRef]
- Han, N.; Mizan, M.F.R.; Jahid, I.K.; Ha, S.-D. Biofilm Formation by Vibrio parahaemolyticus on Food and Food Contact Surfaces Increases with Rise in Temperature. Food Control 2016, 70, 161–166. [Google Scholar] [CrossRef]
- Lopatek, M.; Wieczorek, K.; Osek, J. Antimicrobial Resistance, Virulence Factors, and Genetic Profiles of Vibrio parahaemolyticus from Seafood. Appl. Environ. Microbiol. 2018, 84, e00537-18. [Google Scholar] [CrossRef]
- Lee, L.-H.; Mutalib, N.-S.A.; Law, J.W.-F.; Wong, S.H.; Letchumanan, V. Discovery on Antibiotic Resistance Patterns of Vibrio Parahaemolyticus in Selangor Reveals Carbapenemase Producing Vibrio parahaemolyticus in Marine and Freshwater Fish. Front. Microbiol. 2018, 9, 2513. [Google Scholar] [CrossRef]
- Popova, K.B.; Valsamatzi-Panagiotou, A.; Penchovsky, R. New Drug Discovery Strategies for Targeting Drug-Resistant Bacteria. Environ. Chem. Lett. 2021, 19, 1995–2004. [Google Scholar] [CrossRef]
- Pereira, C.; Costa, P.; Duarte, J.; Balcão, V.M.; Almeida, A. Phage Therapy as a Potential Approach in the Biocontrol of Pathogenic Bacteria Associated with Shellfish Consumption. Int. J. Food Microbiol. 2021, 338, 108995. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Li, J.; Nazir, A.; Tong, Y. Bacteriophage—A Promising Alternative Measure for Bacterial Biofilm Control. Infect. Drug Resist. 2021, 14, 205–217. [Google Scholar] [CrossRef]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; Fiol, F.S.D.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Yehl, K.; Lemire, S.; Yang, A.C.; Ando, H.; Mimee, M.; Torres, M.D.T.; de la Fuente-Nunez, C.; Lu, T.K. Engineering Phage Host-Range and Suppressing Bacterial Resistance through Phage Tail Fiber Mutagenesis. Cell 2019, 179, 459–469.e9. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, C.; Whyte, P.; Brunton, N.; Lyng, J.; Smyth, C.; Fagan, J.; Bolton, D. Spoilage Indicator Bacteria in Farmed Atlantic Salmon (Salmo Salar) Stored on Ice for 10 Days. Food Microbiol. 2019, 77, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Hao, L.; Xia, H.; Wang, H.; Hou, W.; Wang, H.; Zhou, M. Inhibitory Effect of Two Closely Related Phages on Vibrio Parahaemolyticus. Foodborne Pathog. Dis. 2023, 20, 149–157. [Google Scholar] [CrossRef]
- Nobrega, F.L.; Vlot, M.; de Jonge, P.A.; Dreesens, L.L.; Beaumont, H.J.E.; Lavigne, R.; Dutilh, B.E.; Brouns, S.J.J. Targeting Mechanisms of Tailed Bacteriophages. Nat. Rev. Microbiol. 2018, 16, 760–773. [Google Scholar] [CrossRef]
- Hu, M.; Zhang, H.; Gu, D.; Ma, Y.; Zhou, X. Identification of a Novel Bacterial Receptor That Binds Tail Tubular Proteins and Mediates Phage Infection of Vibrio parahaemolyticus. Emerg. Microbes Infect. 2020, 9, 855–867. [Google Scholar] [CrossRef]
- Grabowski, Ł.; Łepek, K.; Stasiłojć, M.; Kosznik-Kwaśnicka, K.; Zdrojewska, K.; Maciąg-Dorszyńska, M.; Węgrzyn, G.; Węgrzyn, A. Bacteriophage-Encoded Enzymes Destroying Bacterial Cell Membranes and Walls, and Their Potential Use as Antimicrobial Agents. Microbiol. Res. 2021, 248, 126746. [Google Scholar] [CrossRef]
- Ge, H.; Fu, S.; Guo, H.; Hu, M.; Xu, Z.; Zhou, X.; Chen, X.; Jiao, X. Application and Challenge of Bacteriophage in the Food Protection. Int. J. Food Microbiol. 2022, 380, 109872. [Google Scholar] [CrossRef] [PubMed]
- Mudiyanselage, C.S.S.S.; Farouk, M.M.; Realini, C.E.; Kantono, K.; Hamid, N. Determinants of Purchase Intention for Meat-Based Chilled Ready Meals in New Zealand: A Consumer Behaviour Perspective. Foods 2025, 14, 1038. [Google Scholar] [CrossRef] [PubMed]
- Altintzoglou, T.; Heide, M.; Wien, A.H.; Honkanen, P. Traditional Sushi for Modern Consumers: A Comparison Between Sushi Consumption Behavior in Japan and Norway. J. Food Prod. Mark. 2016, 22, 717–732. [Google Scholar] [CrossRef]
- Kim, S.; Kim, S. Recent Surge of Ready Meals in South Korea: Can They Be Healthy Alternatives? Public Health Nutr. 2019, 23, 711–720. [Google Scholar] [CrossRef]
- Kulawik, P.; Dordević, D. Sushi Processing: Microbiological Hazards and the Use of Emerging Technologies. Crit. Rev. Food Sci. Nutr. 2020, 62, 1270–1283. [Google Scholar] [CrossRef] [PubMed]
- Giannoglou, M.; Evangelopoulou, A.-M.; Perikleous, N.; Baclori, C.; Tsironi, T.; Taoukis, P. Time Temperature Integrators for Monitoring the Shelf Life of Ready-to-Eat Chilled Smoked Fish Products. Food Packag. Shelf Life 2019, 22, 100403. [Google Scholar] [CrossRef]
- Tao, Z.; Sun, Y.; Zhou, M.; Li, T.; Lan, W.; Zhao, Y.; Sun, X. Two novel Vibrio parahaemolyticus phages (vB_VpaS_1601 and vB_VpaP_1701) can serve as potential biocontrol agents against Vibrio parahaemolyticus infection in seafood. Int. J. Food Microbiol. 2025, 441, 111303. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Zhou, Y.; Bao, H.; Wang, R.; Li, T.; Pang, M.; Sun, L.; Zhou, X. Application of a phage in decontaminating Vibrio parahaemolyticus in oysters. Int. J. Food Microbiol. 2018, 275, 24–31. [Google Scholar] [CrossRef]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef]
- Malik, S.; Nehra, K.; Rana, J.S. Bacteriophage cocktail and phage antibiotic synergism as promising alternatives to conventional antibiotics for the control of multi-drug-resistant uropathogenic Escherichia coli. Virus Res. 2021, 302, 198496. [Google Scholar] [CrossRef] [PubMed]
- You, H.J.; Lee, J.H.; Oh, M.; Hong, S.Y.; Kim, D.; Noh, J.; Kim, M.; Kim, B.S. Tackling Vibrio parahaemolyticus in Ready-to-Eat Raw Fish Flesh Slices Using Lytic Phage VPT02 Isolated from Market Oyster. Food Res. Int. 2021, 150, 110779. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Plaque Formation a | Source b |
|---|---|---|---|
| ATCC 17802 | Vibrio parahaemolyticus | − | ATCC |
| GD91 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD103 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD71 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD78 | Vibrio parahaemolyticus | − | Laboratory collection |
| GD81 | Vibrio parahaemolyticus | − | Laboratory collection |
| GD62 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD75 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD74 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD89 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD80 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD83 | Vibrio parahaemolyticus | + | Laboratory collection |
| GD73 | Vibrio parahaemolyticus | − | Laboratory collection |
| 99-1 | Vibrio parahaemolyticus | − | Food |
| 66-1 | Vibrio parahaemolyticus | − | Seawater |
| 124-1 | Vibrio parahaemolyticus | − | Food |
| 84-1 | Vibrio parahaemolyticus | + | Food |
| 163-1 | Vibrio parahaemolyticus | − | Freshwater |
| 240-1 | Vibrio parahaemolyticus | − | Food |
| 79-1 | Vibrio parahaemolyticus | − | Food |
| 87-2 | Vibrio parahaemolyticus | − | Freshwater |
| 84-2 | Vibrio parahaemolyticus | − | Food |
| 139-1 | Vibrio parahaemolyticus | − | Food |
| 227-2 | Vibrio parahaemolyticus | − | Food |
| 51-1 | Vibrio parahaemolyticus | − | Food |
| 75-2 | Vibrio parahaemolyticus | − | Food |
| ATCC 13076 | Salmonella enterica subsp. enterica | − | ATCC |
| ATCC 19111 | Listeria monocytogenes | − | ATCC |
| ATCC 51329 | Cronobacter muytjensii | − | ATCC |
| ATCC 19433 | Enterococcus faecalis | − | ATCC |
| ATCC 14028 | Salmonella typhimurium | − | ATCC |
| ATCC 49128 | Pseudomonas putida | − | ATCC |
| ATCC 27583 | Pseudomonas aeruginosa | − | ATCC |
| ATCC 6538 | Staphylococcus aureus | − | ATCC |
| ORF | Predicted Function | Best BLAST Hit | Functional Module |
|---|---|---|---|
| 5 | endonuclease | Vibrio phage VP46 | DNA metabolism |
| 10 | metallopeptidase activity protein | Vibrio phage VP46 | Additional function |
| 12 | DNA primase | Vibrio phage vB_VpaP_GHSM17 | DNA metabolism |
| 13 | putative DNA helicase | Vibrio phage vB_VpaP_KF1 | DNA metabolism |
| 15 | DNA-directed DNA polymerase | Vibrio phage vB_VpP_HA1 | DNA metabolism |
| 17 | nucleotidyltransferase | Vibrio phage VP9 | DNA metabolism |
| 20 | MazG-like pyrophosphatase | Vibrio phage F23s2 | DNA metabolism |
| 23 | exonuclease | Vibrio phage vB_VpP_NS8 | DNA metabolism |
| 25 | endonuclease VII | Vibrio phage VP48 | DNA metabolism |
| 27 | putative deoxynucleoside monophosphate kinase | Vibrio phage vB_VpP_DE18 | DNA metabolism |
| 29 | putative RNA polymerase | Vibrio phage H256D1 | DNA metabolism |
| 31 | virion structural protein | Vibrio phage VP48 | Structure |
| 32 | head–tail connector protein | Vibrio phage OY1 | Structure |
| 33 | head scaffolding protein | Vibrio phage vB_VpaP_KF1 | Structure |
| 34 | putative capsid protein | Vibrio phage vB_VpP_FE11 | Structure |
| 36 | putative tail tubular A | Vibrio phage vB_VpaP_KF2 | Structure |
| 37 | putative tail tubular protein B | Vibrio phage VP46 | DNA metabolism |
| 38 | tail protein | Vibrio phage vB_VpP_HA5 | Structure |
| 39 | internal virion protein | Vibrio phage vB_VpP_DE10 | Additional function |
| 40 | internal virion lysozyme motif protein | Vibrio phage vB_VpP_DE10 | Additional function |
| 41 | peptidoglycan lytic exotransglycosylase | Vibrio phage vB_VpP_DE10 | Lysis |
| 42 | tail fiber protein | Vibrio phage VP46 | Structure |
| 43 | putative glycosyl hydrolase | Vibrio phage vB_VpaP_KF1 | Additional function |
| 44 | terminase small subunit | Vibrio phage vB_VpaP_KF1 | Additional function |
| 45 | terminase large subunit | Vibrio phage BUCT233 | Additional function |
| 48 | bacterial Ig-like domain family protein | Vibrio phage vB_VpP_DE10 | Additional function |
| 49 | endolysin | Vibrio phage VP46 | Lysis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, Z.; Cheng, X.; Jiang, S.; Zhang, Z.; Zhang, D.; Chen, H.; Ling, N.; Ye, Y. Isolation and Characterization of a Cold-Adapted Bacteriophage for Biocontrol of Vibrio parahaemolyticus in Seafood. Foods 2025, 14, 2660. https://doi.org/10.3390/foods14152660
Nie Z, Cheng X, Jiang S, Zhang Z, Zhang D, Chen H, Ling N, Ye Y. Isolation and Characterization of a Cold-Adapted Bacteriophage for Biocontrol of Vibrio parahaemolyticus in Seafood. Foods. 2025; 14(15):2660. https://doi.org/10.3390/foods14152660
Chicago/Turabian StyleNie, Zhixiang, Xiangyu Cheng, Shengshi Jiang, Zhibin Zhang, Diwei Zhang, Hanfang Chen, Na Ling, and Yingwang Ye. 2025. "Isolation and Characterization of a Cold-Adapted Bacteriophage for Biocontrol of Vibrio parahaemolyticus in Seafood" Foods 14, no. 15: 2660. https://doi.org/10.3390/foods14152660
APA StyleNie, Z., Cheng, X., Jiang, S., Zhang, Z., Zhang, D., Chen, H., Ling, N., & Ye, Y. (2025). Isolation and Characterization of a Cold-Adapted Bacteriophage for Biocontrol of Vibrio parahaemolyticus in Seafood. Foods, 14(15), 2660. https://doi.org/10.3390/foods14152660