
Donkey-Hide Gelatin Peptide-Iron Complexes: Structural Characterization, Enhanced Iron Solubility Under Simulated Digestion, and Dual Iron Chelation-Antioxidant Functions

Abstract

1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Low Molecular Weight Donkey-Hide Gelatin Peptide (LMW DHGP) by Enzymolysis
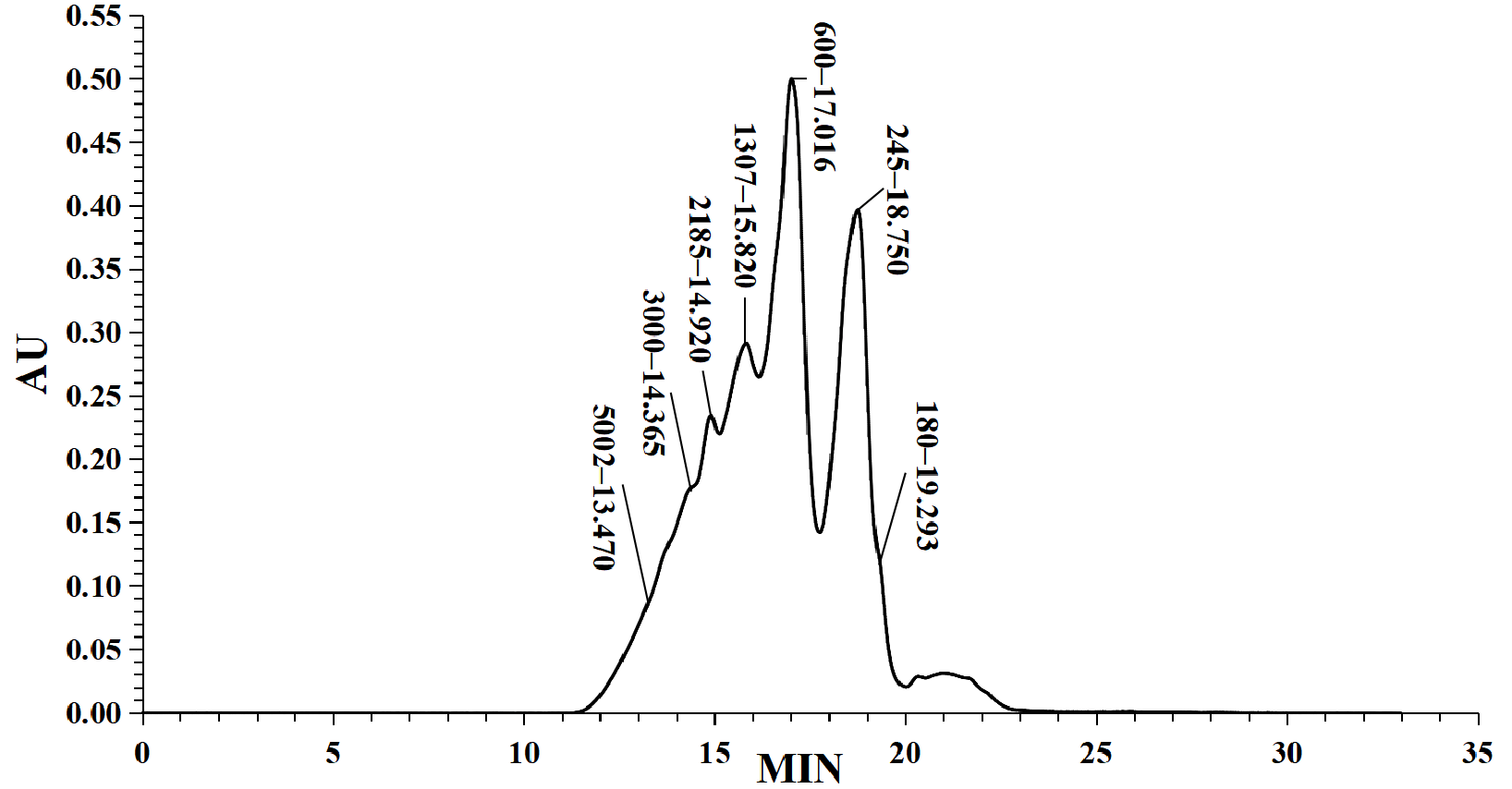
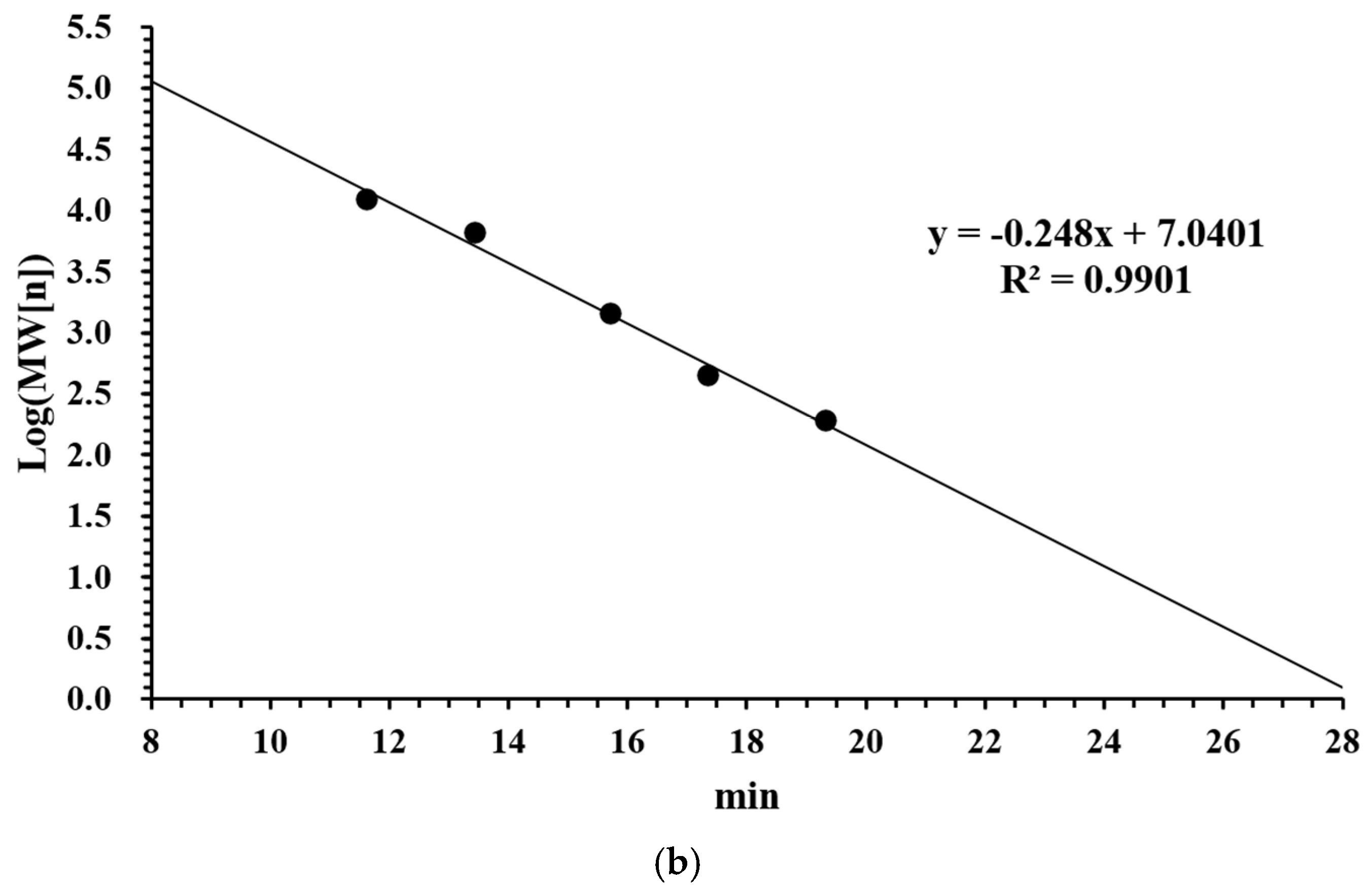
2.3. Determination of Molecular Weight Distribution of LMW DHGP
2.4. Determination of the Iron Ion Binding Capacity of LMW DHGP
2.5. Chelation of LMW DHGP with Ferrous Chloride
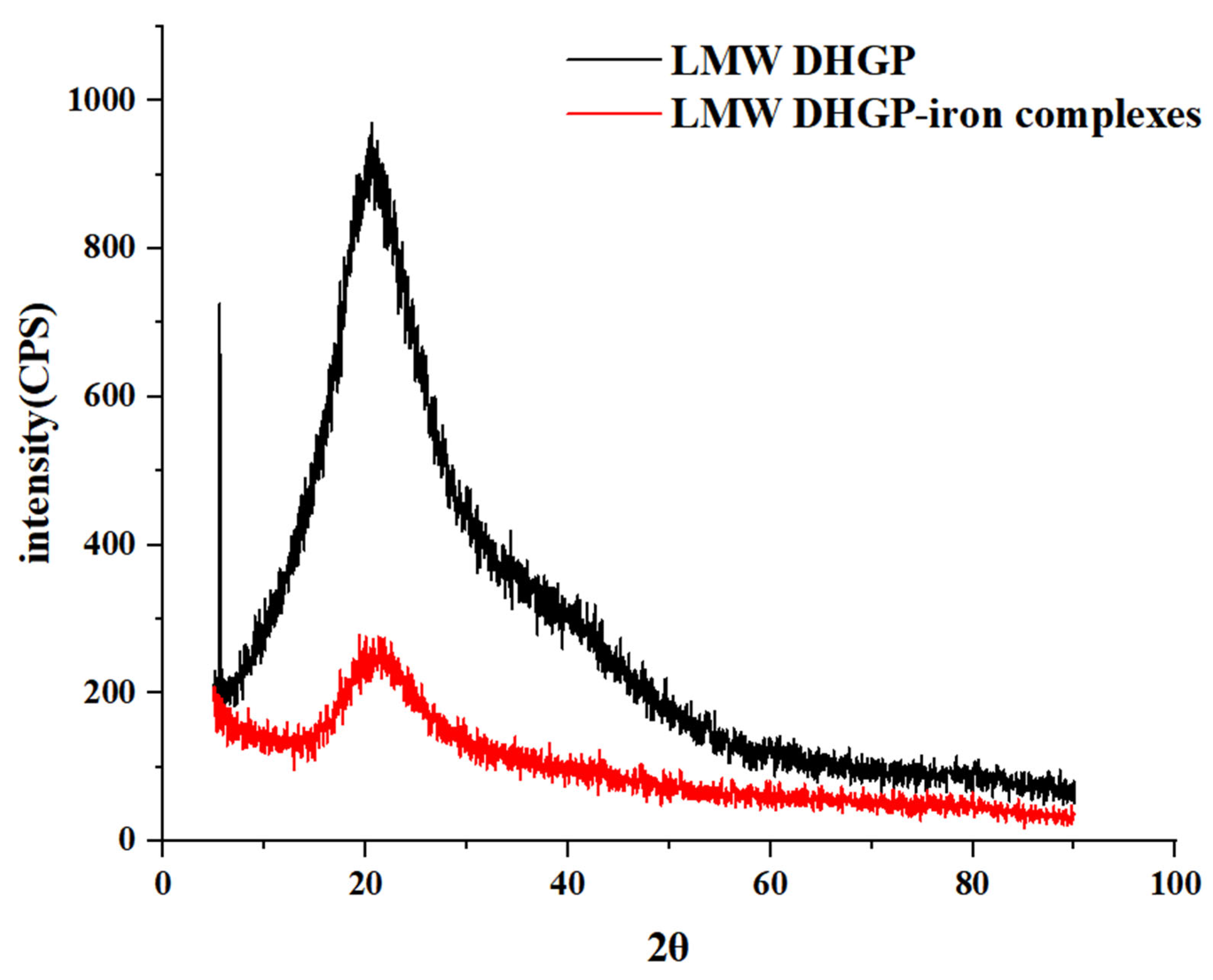
2.6. Scanning Electron Microscopy (SEM) and X-Ray Diffraction (XRD) Analysis of LMW DHGP and LMW DHGP-Iron Complexes
2.7. Analysis of the Amino Acid Composition of LMW DHGP and LMW DHGP-Iron Complexes
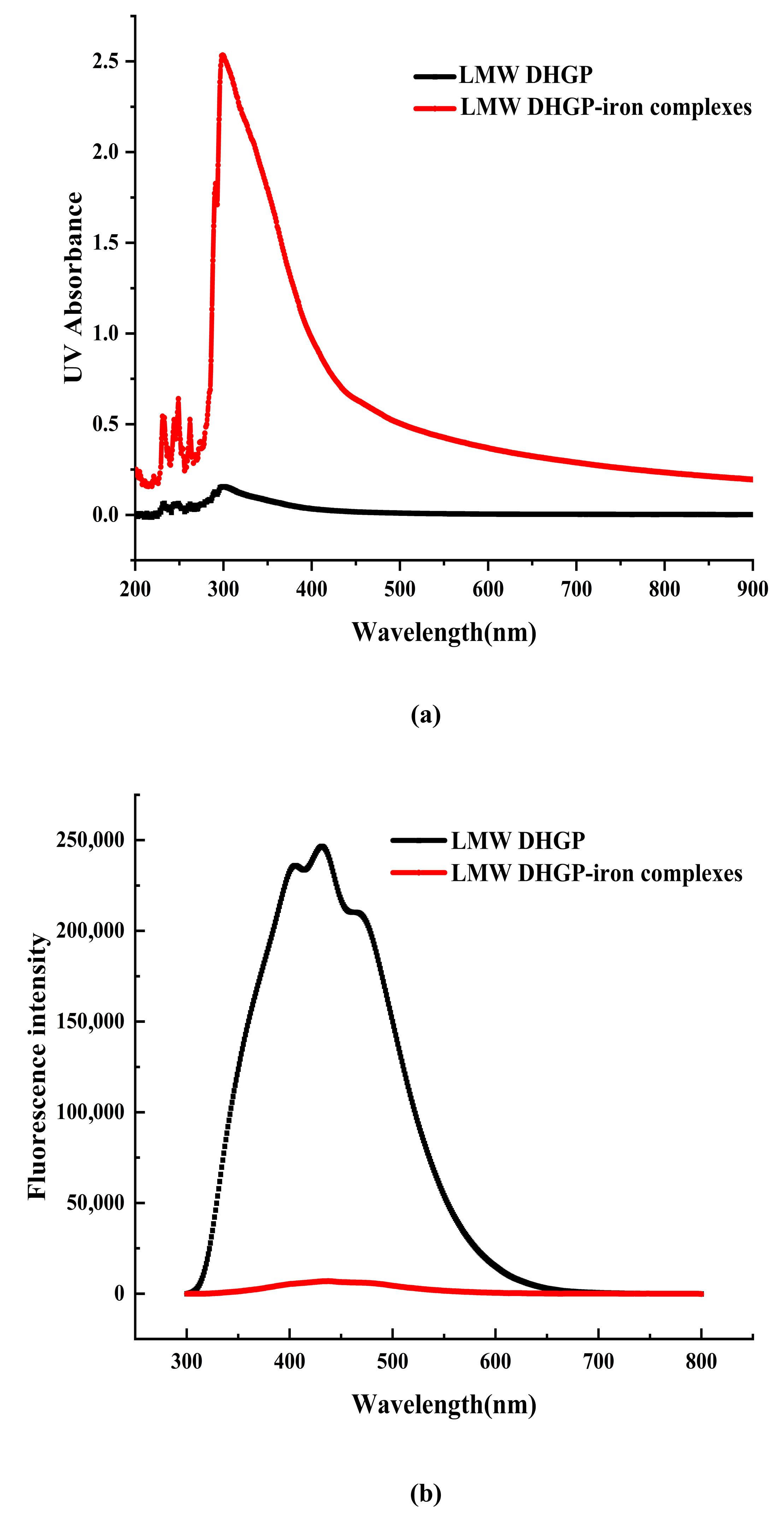
2.8. UV and FL Analysis of LMW DHGP and LMW DHGP-Iron Complexes
2.9. Fourier Transform Infrared Spectroscopy (FTIR) Analysis of LMW DHGP and LMW DHGP-Iron Complexes
2.10. Circular Dichroism Spectroscopy (CD) Analysis of LMW DHGP and LMW DHGP-Iron Complexes
2.11. In Vitro Assessment of the Solubility Stability of LMW DHGP and LMW DHGP-Iron Complexes
2.12. Determination of the Antioxidant Activity
2.13. Statistical Analysis
3. Results and Discussion
3.1. Molecular Weight Distribution and Iron Chelating Ability of LMW DHGP
3.2. Morphological Characterization and X-Ray Diffraction of LMW DHGP and LMW DHGP-Iron Complexes
3.3. Amino Acid Composition of LMW DHGP and LMW DHGP-Iron Complexes
3.4. UV and FL Analysis of LMW DHGP and LMW DHGP-Iron Complexes
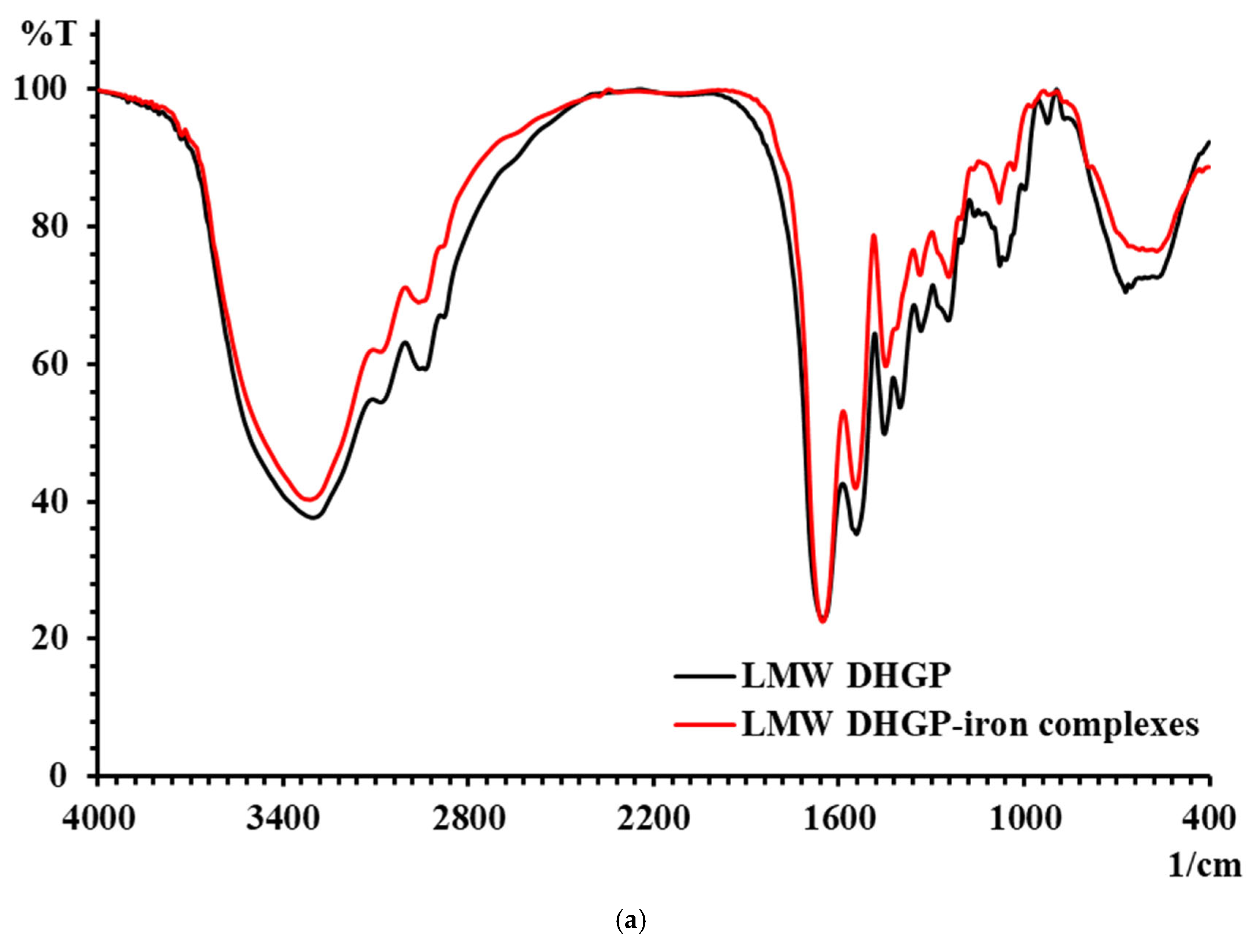
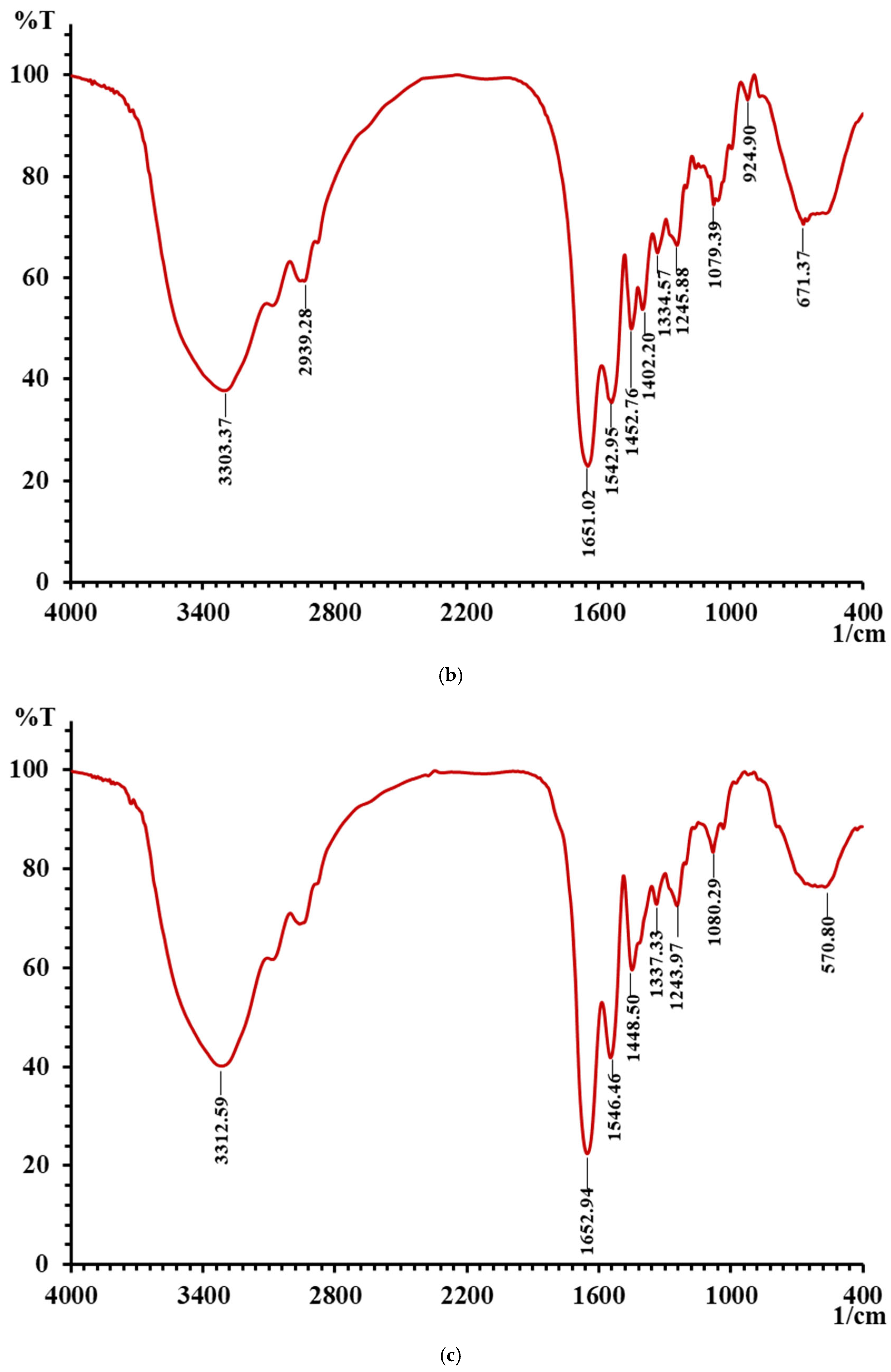
3.5. Results of FTIR of LMW DHGP and LMW DHGP-Iron Complexes
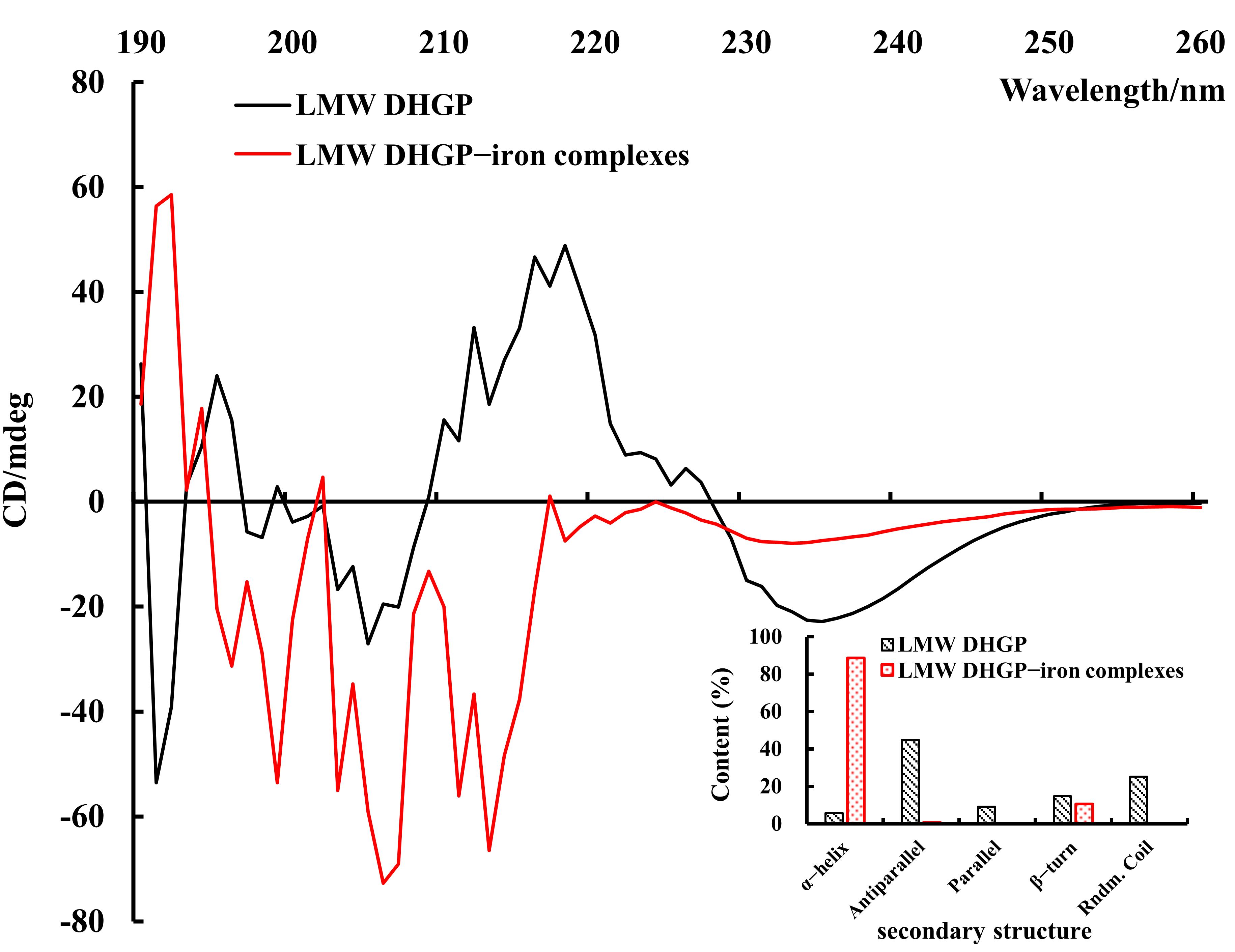
3.6. Secondary Structure of LMW DHGP and LMW DHGP-Iron Complexes
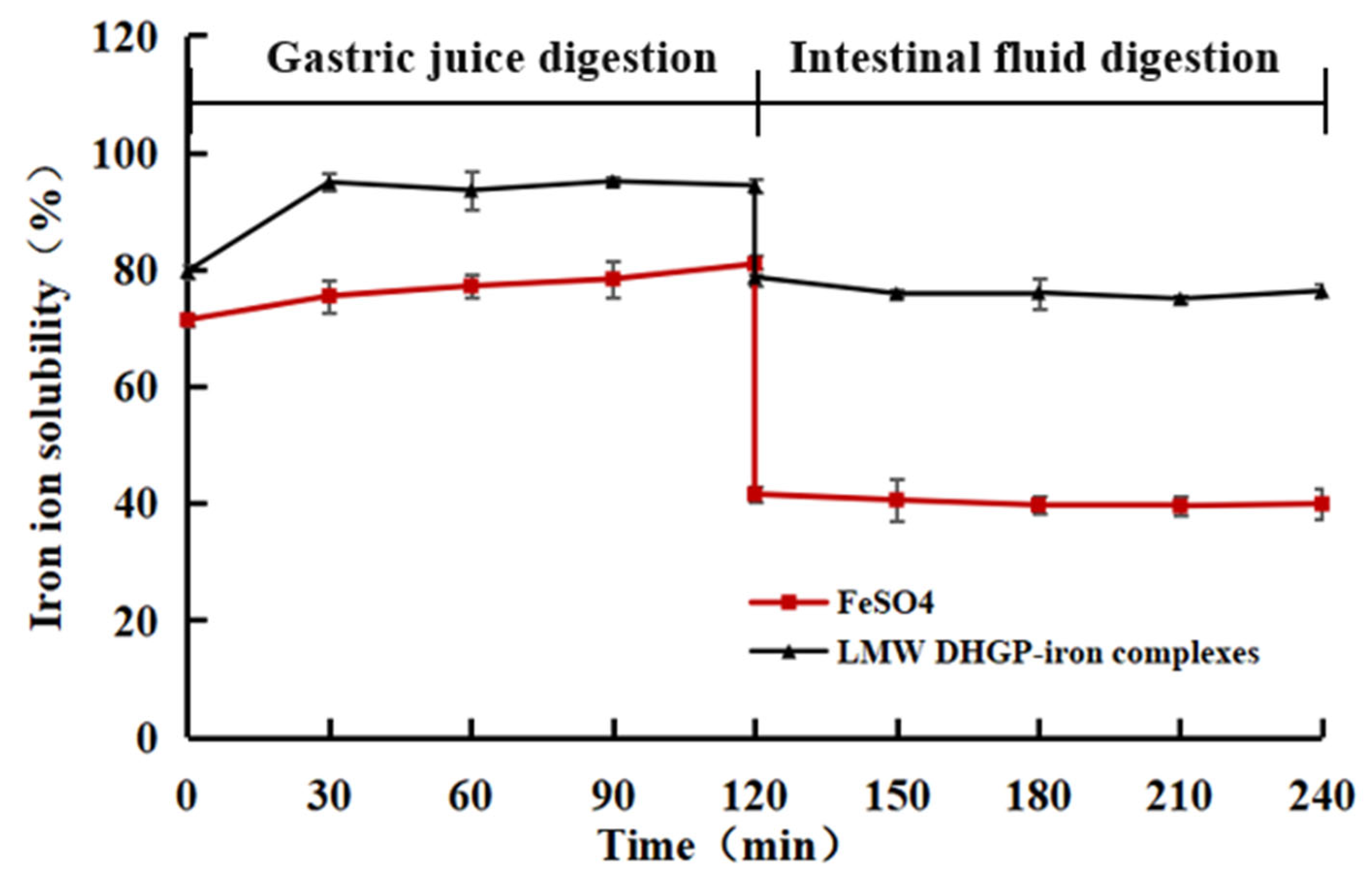
3.7. In Vitro Assessment of the Solubility Stability of LMW DHGP and LMW DHGP-Iron Complexes
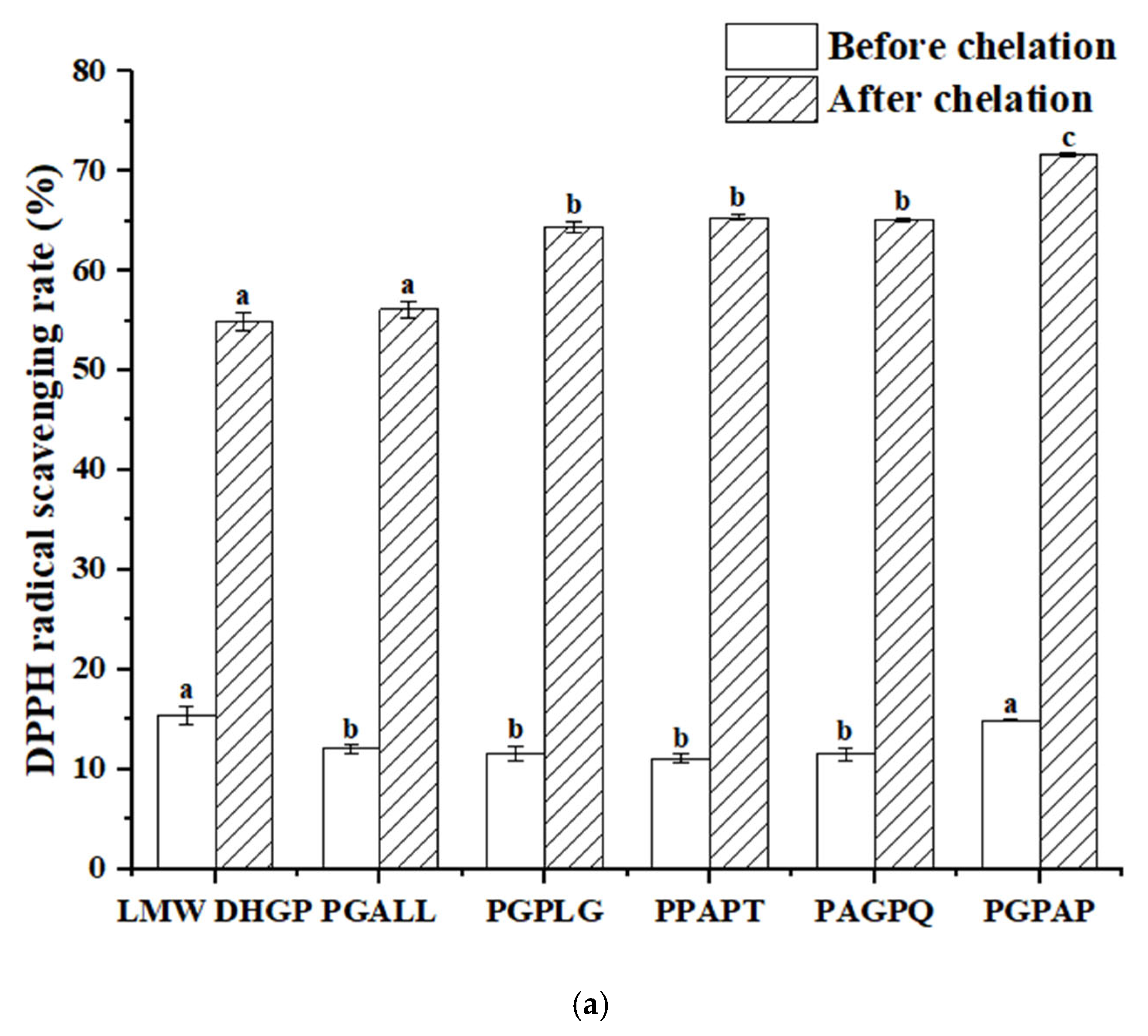
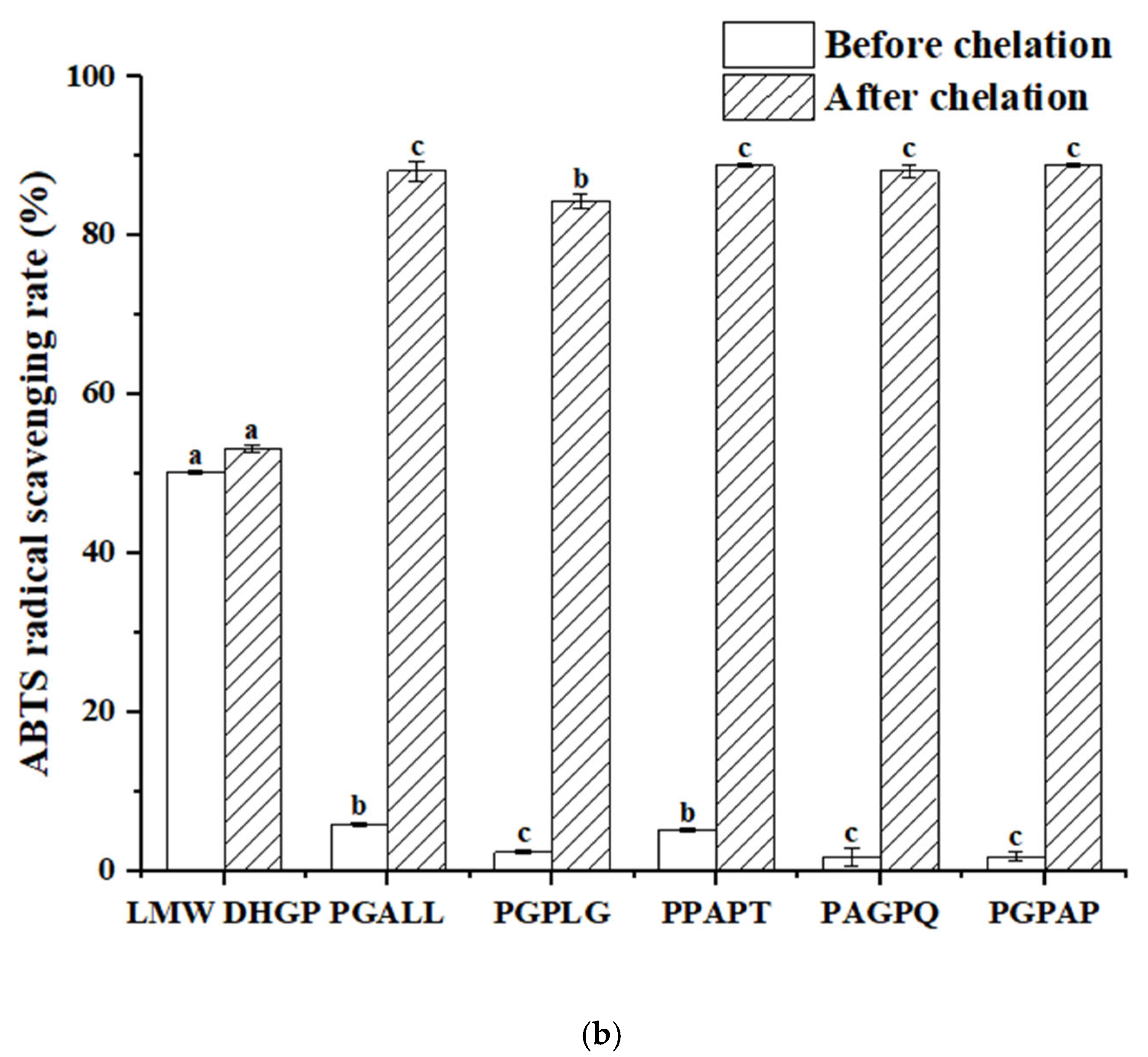
3.8. Determination of the Antioxidant Activity
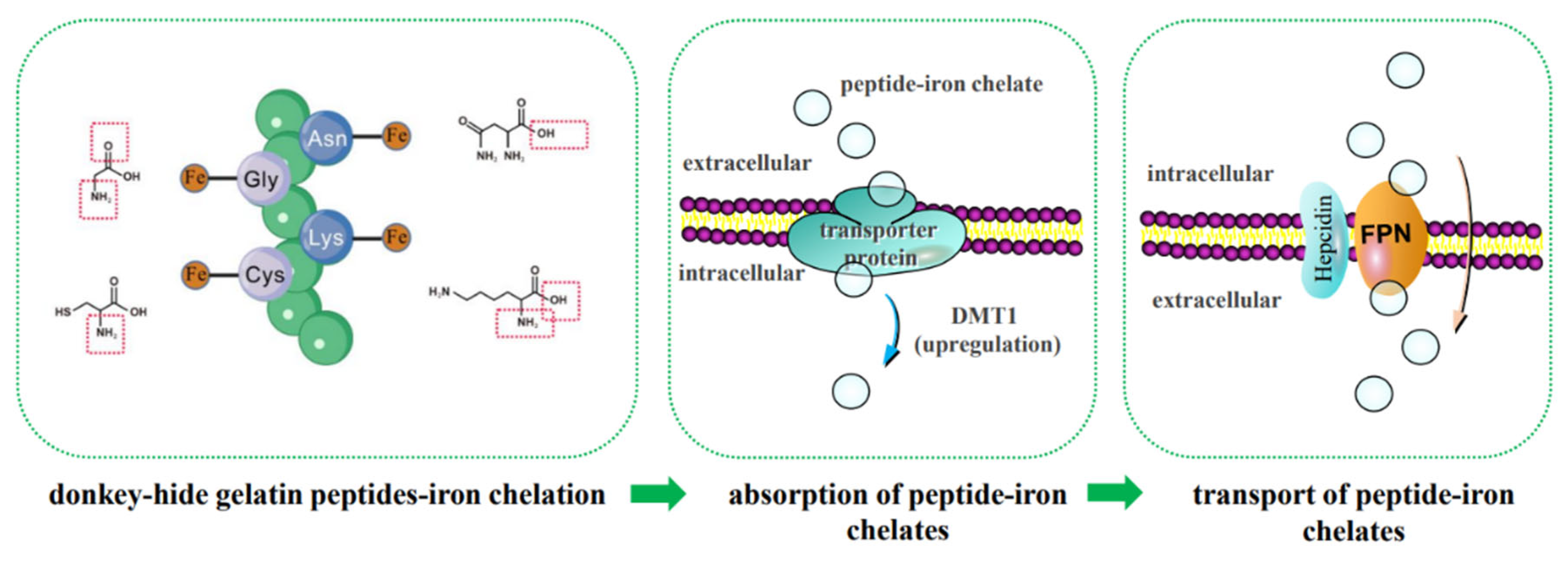
3.9. Prediction of the Mechanism of Iron Absorption Promotion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LMW | Low molecular weight |
| DHGP | Donkey-hide gelatin peptide |
| FTIR | Fourier transform infrared |
| CD | Circular dichroism |
| SEM | Scanning electron microscopy |
| XRD | X-ray diffraction |
| FL | Fluorescence |
References
- Kazemi-Taskooh, Z.; Varidi, M. Food-based iron delivery systems: A review. Trends Food Sci. Technol. 2021, 116, 75–89. [Google Scholar] [CrossRef]
- Sun, N.; Cui, P.B.; Jin, Z.Q.; Wu, H.T.; Wang, Y.X.; Lin, S.Y. Contributions of molecular size, charge distribution, and specific amino acids to the iron-binding capacity of sea cucumber (Stichopus japonicus) ovum hydrolysates. Food Chem. 2017, 230, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.J.; Frazer, D.M. Current understanding of iron homeostasis. Am. J. Clin. Nutr. 2017, 106, 1559S–1566S. [Google Scholar] [CrossRef]
- Gupta, C.; Chawla, P.; Arora, S.; Tomar, S.K.; Singh, A.K. Iron microencapsulation with blend of gum arabic, maltodextrin and modified starch using modified solvent evaporation method–Milk fortification. Food Hydrocoll. 2015, 43, 622–628. [Google Scholar] [CrossRef]
- Lin, S.; Hu, X.; Yang, X.; Chen, S.; Wu, Y.; Hao, S.; Huang, H.; Li, L. GLPGPSGEEGKR: Fe2+ chelating characterization and potential transport pathways for improving Fe2+ bioavailability in Caco-2 cells. Food Biosci. 2022, 48, 101806. [Google Scholar] [CrossRef]
- Wu, W.F.; Yang, Y.Y.; Sun, N.; Bao, Z.J.; Lin, S.Y. Food protein-derived iron-chelating peptides: The binding mode and promotive effects of iron bioavailability. Food Res. Int. 2020, 131, 108976. [Google Scholar] [CrossRef]
- Caetano-Silva, M.E.; Cilla, A.; Bertoldo-Pacheco, M.T.; Netto, F.M.; Alegría, A. Evaluation of in vitro iron bioavailability in free form and as whey peptide-iron complexes. J. Food Compos. Anal. 2018, 68, 95–100. [Google Scholar] [CrossRef]
- Shilpashree, B.G.; Arora, S.; Sharma, V.; Bajaj, R.K.; Tomar, S.K. Preparation of iron bound succinylated milk protein concentrate and evaluation of its stability. Food Chem. 2016, 196, 800–807. [Google Scholar] [CrossRef]
- Yuan, B.; Zhao, C.; Cheng, C.; Huang, D.-C.; Cheng, S.-J.; Cao, C.-J.; Chen, G.-T. A peptide-Fe(II) complex from Grifola frondosa protein hydrolysates and its immunomodulatory activity. Food Biosci. 2019, 32, 100459. [Google Scholar] [CrossRef]
- Chen, K.; Guo, L.; Lao, L.; Ma, F.; Cao, Y.; Miao, J. Preparation, characterization and enhancement of intestinal iron absorption activity of β-casein phosphopeptides-iron chelate. Process Biochem. 2024, 146, 401–411. [Google Scholar] [CrossRef]
- Athira, S.; Mann, B.; Sharma, R.; Pothuraju, R.; Bajaj, R.K. Preparation and characterization of iron-chelating peptides from whey protein: An alternative approach for chemical iron fortification. Food Res. Int. 2021, 141, 110133. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Jia, J.; Wen, C.; Yu, C.; Zhao, Q.; Hu, J. Optimization of ultrasound assisted extraction of abalone viscera protein and its effect on the iron-chelating activity. Ultrason. Sonochem. 2021, 77, 105670. [Google Scholar] [CrossRef] [PubMed]
- Cian, R.E.; Garzón, A.G.; Ancona, D.B.; Guerrero, L.C.; Drago, S.R. Chelating Properties of Peptides from Red Seaweed Pyropia columbina and Its Effect on Iron Bio-Accessibility. Plant Foods Hum. Nutr. 2016, 71, 96–101. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, X.Y.; Si, X.X.; Lai, W.F. Preparation, characterization, and properties of wampee seed antioxidant peptides-iron chelate. J. Food Process. Preserv. 2022, 46, e16513. [Google Scholar] [CrossRef]
- Ma, N.; Guan, R.; Zhao, R.T.; Geng, Y. GC-IMS-Based Preliminary Analysis of Volatile Flavor Compounds in Ejiao at Different Processing Stages. J. Food Qual. 2022, 2022, 3961593. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Z.J.; Jiang, T.; Zhang, J.L.; Huang, P.X.; Tan, J.Q.; Chen, G.; Yuan, M.; Li, Z.; Liu, H.B.; et al. Efficacy and Safety of Ejiao (Asini Corii Colla) in Women With Blood Deficient Symptoms: A Randomized, Double-Blind, and Placebo-Controlled Clinical Trial. Front. Pharmacol. 2021, 12, 718154. [Google Scholar] [CrossRef]
- Yu, X.-J.; Kong, L.-M.; Wang, B.; Zhai, C.-Q.; Lao, Y.-Z.; Zhang, L.-J.; Gao, Y.; Zhao, B.-N. Asini Corii Colla (Ejiao) as a health-care food and traditional Chinese medicine: A review of its chemical composition, pharmacological activity, quality control, modern applications. J. Funct. Foods 2025, 125, 106678. [Google Scholar] [CrossRef]
- Cheng, X.-R.; Chen, Y.-Y.; Cao, C.-C.; Ma, J.-H.; Zhang, C.-X.; Zhao, Z.-W.; Wu, Q.; Li, Y.-Y.; Muskat, M.N.; Wu, G.-Q. Ejiao peptide-iron chelates regulate the metabolism of iron deficiency anemia mice and improve the bioavailability of iron. Food Biosci. 2023, 54, 102835. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kim, D.; Kim, H.-J.; Jang, A. Protection effect of donkey hide gelatin hydrolysates on UVB-induced photoaging of human skin fibroblasts. Process Biochem. 2018, 67, 118–126. [Google Scholar] [CrossRef]
- Yan, X.; Yue, Y.; Guo, B.; Zhang, S.; Ji, C.; Chen, Y.; Dai, Y.; Dong, L.; Zhu, B.; Lin, X. Novel microbial fermentation for the preparation of iron-chelating scallop skirts peptides-its profile, identification, and possible binding mode. Food Chem. 2024, 451, 139493. [Google Scholar] [CrossRef]
- Wang, Y.H.; Cai, M.; Zeng, H.; Zhao, H.; Zhang, M.; Yang, Z.N. Preparation, Characterization and Iron Absorption by Caco-2 Cells of the Casein Peptides-Iron Chelate. Int. J. Pept. Res. Ther. 2022, 28, 116. [Google Scholar] [CrossRef]
- Chen, J.-Q.; Zhang, Z.; Fei, Y.-Y.; Chen, X.-Q. Preparation process optimization and structure characterization of sheep bone polypeptide chelate with ferrous iron. Food Ferment. Sci. Technol. 2021, 57, 1–21. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Han, Z.; Zeng, X.-A.; Wang, M.-S. The preparation of Fe-glycine complexes by a novel method (pulsed electric fields). Food Chem. 2017, 219, 468–476. [Google Scholar] [CrossRef]
- Yuan, H.; Chai, L.; Wang, L.; Tian, C.; Wang, X. Preparation of iron-chelating peptides of Zanthoxylum bungeanum seed and its physicochemical properties. Food Ferment. Ind. 2018, 44, 164–171. [Google Scholar] [CrossRef]
- Zheng, Y.; Shi, P.; Wang, G.; Wang, X.; Li, Y.; Liu, L.; Li, G. Preparation of Coconut Globulin Peptide-Ferrous Chelate. Chin. J. Trop. Crops 2021, 42, 1485–1491. [Google Scholar] [CrossRef]
- Ding, X.; Li, H.; Xu, M.; Li, X.; Li, M. Peptide composition analysis, structural characterization, and prediction of iron binding modes of small molecular weight peptides from mung bean. Food Res. Int. 2024, 175, 113735. [Google Scholar] [CrossRef]
- Qu, W.; Feng, Y.; Xiong, T.; Qayum, A.; Ma, H. Preparation, structural and functional characterization of corn peptide-chelated calcium microcapsules using synchronous dual frequency ultrasound. Ultrason. Sonochem. 2024, 102, 106732. [Google Scholar] [CrossRef]
- Liang, R.; Xu, L.; Fan, C.; Cao, L.L.; Guo, X.F. Structural Characteristics and Antioxidant Mechanism of Donkey-Hide Gelatin Peptides by Molecular Dynamics Simulation. Molecules 2023, 28, 7975. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Hu, X.; Li, L.; Yang, X.; Chen, S.; Wu, Y.; Yang, S. Preparation, purification and identification of iron-chelating peptides derived from tilapia (Oreochromis niloticus) skin collagen and characterization of the peptide-iron complexes. LWT 2021, 149, 111796. [Google Scholar] [CrossRef]
- Wang, Q.-Q.; Cai, W.-R.; Wang, Y.-L.; Zhuo, Y.-Y.; Huang, W.; Cao, Y.-F. Preparation, characterization, and proliferation by Caco-2 cells of egg white polypeptide-iron chelate. Food Ferment. Ind. 2024, 50, 197–203. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Ma, Z.; Mu, G.; Qian, F. Novel insights into whey protein peptide-iron chelating agents: Structural characterization, in vitro stability and functional properties. Food Biosci. 2024, 60, 104317. [Google Scholar] [CrossRef]
- Qin, Y.; Ju, S.-Y. Preparation and structure characterization for iron chelates of fish scale polypeptide. Southwest China J. Agric. Sci. 2020, 33, 456–463. [Google Scholar] [CrossRef]
- Fan, C.; Ge, X.; Hao, J.; Wu, T.; Liu, R.; Sui, W.; Geng, J.; Zhang, M. Identification of high iron–chelating peptides with unusual antioxidant effect from sea cucumbers and the possible binding mode. Food Chem. 2023, 399, 133912. [Google Scholar] [CrossRef]
- Yang, J.; Shi, J.; Zou, Y.; Yang, B.; Ma, J.-J.; Xu, W.-M.; Wang, D.-Y. Preparation process optimization and structural characterization of chicken blood peptides-iron chelate. Jiangsu J. Agric. Sci. 2022, 38, 1678–1685. [Google Scholar] [CrossRef]
- Du, Y.-N.; Hong, J.-N.; Xu, S.-Q.; Wang, Y.-Q.; Wang, X.-C.; Yan, J.-N.; Bin, L.; Wu, H.-T. Iron-chelating activity of large yellow croaker (Pseudosciaena crocea) roe hydrolysates. J. Food Process. Preserv. 2022, 46, e17080. [Google Scholar] [CrossRef]
- Yang, Y.; Li, A.; Zhong, Z.; Li, G. Structural Characterization and in Vitro Simulated Digestion of Peach Kernel Peptide-Ferrous Complex. J. Chin. Inst. Food Sci. Technol. 2020, 20, 61–69. [Google Scholar] [CrossRef]
- Fan, C.; Wang, X.; Song, X.; Sun, R.; Liu, R.; Sui, W.; Jin, Y.; Wu, T.; Zhang, M. Identification of a Novel Walnut Iron Chelating Peptide with Potential High Antioxidant Activity and Analysis of Its Possible Binding Sites. Foods 2023, 12, 226. [Google Scholar] [CrossRef]
- Zhao, Q.; Liang, W.; Xiong, Z.; Li, C.; Zhang, L.; Rong, J.; Xiong, S.; Liu, R.; You, J.; Yin, T.; et al. Digestion and absorption characteristics of iron-chelating silver carp scale collagen peptide and insights into their chelation mechanism. Food Res. Int. 2024, 190, 114612. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, Z.; Zhao, Y.; Zeng, M. Enzymatic preparation and characterization of iron-chelating peptides from anchovy (Engraulis japonicus) muscle protein. Food Res. Int. 2012, 48, 435–441. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, X.; Li, M. Preparation, characterization and in vitro stability of iron-chelating peptides from mung beans. Food Chem. 2021, 349, 129101. [Google Scholar] [CrossRef]
- Lv, J.; Feng, J.; Zhong, H.; Lou, Y.; Wang, Y.; Liu, S.; Xu, H.; Xia, G. Preparation, characterization and antioxidant effect of polypeptide mineral-chelate from Yanbian cattle bone. LWT 2023, 187, 115353. [Google Scholar] [CrossRef]
- Li, Y.N.; Jiang, H.; Huang, G.R. Protein Hydrolysates as Promoters of Non-Haem Iron Absorption. Nutrients 2017, 9, 609. [Google Scholar] [CrossRef] [PubMed]
- Chaud, M.V.; Izumi, C.; Nahaal, Z.; Shuhama, T.; Bianchi, M.D.P.; de Freitas, O. Iron derivatives from casein hydrolysates as a potential source in the treatment of iron deficiency. J. Agric. Food Chem. 2002, 50, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Yuanqing, H.; Pengyao, Y.; Yangyang, D.; Min, C.; Rui, G.; Yuqing, D.; Haihui, Z.; Haile, M. The Preparation, Antioxidant Activity Evaluation, and Iron-Deficient Anemic Improvement of Oat (Avena sativa L.) Peptides-Ferrous Chelate. Front. Nutr. 2021, 8, 687133. [Google Scholar] [CrossRef]
- Zhou, J.; Mao, X.-Y.; Wang, X.; Ai, T.; Ma, J.-J.; Li, Y.-H. Anti-anaemia efficacy of β-lactoglobulin hydrolysate-iron complex on iron-deficient anaemic rats. Eur. J. Nutr. 2014, 53, 877–884. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Amino Acid Type | Amino Acid Content of Donkey-Hide Gelatin Peptides (mg/g) | |
|---|---|---|---|
| LMW DHGP | LMW DHGP-Iron Complexes | ||
| 1 | Asn (N) | 54.40 | 59.61 |
| 2 | Thr (T) | 18.14 | 14.24 |
| 3 | Ser (S) | 34.67 | 29.18 |
| 4 | Glu (E) | 96.13 | 95.40 |
| 5 | Gly (G) | 127.91 | 128.24 |
| 6 | Ala (A) | 72.32 | 54.26 |
| 7 | Cys (C) | 16.96 | 21.37 |
| 8 | Val (V) | 24.85 | 18.20 |
| 9 | Met (M) | 9.49 | 7.73 |
| 10 | Ile (I) | 14.08 | 8.38 |
| 11 | Leu (L) | 30.43 | 18.43 |
| 12 | Tyr (Y) | 9.18 | 8.60 |
| 13 | Phe (F) | 19.68 | 13.22 |
| 14 | His (H) | 7.74 | 7.70 |
| 15 | Lys (K) | 38.54 | 39.07 |
| 16 | Arg (R) | 64.85 | 59.80 |
| 17 | Pro (P) | 110.37 | 80.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Lv, C.; Guo, X.; Liang, R. Donkey-Hide Gelatin Peptide-Iron Complexes: Structural Characterization, Enhanced Iron Solubility Under Simulated Digestion, and Dual Iron Chelation-Antioxidant Functions. Foods 2025, 14, 2117. https://doi.org/10.3390/foods14122117
Yang L, Lv C, Guo X, Liang R. Donkey-Hide Gelatin Peptide-Iron Complexes: Structural Characterization, Enhanced Iron Solubility Under Simulated Digestion, and Dual Iron Chelation-Antioxidant Functions. Foods. 2025; 14(12):2117. https://doi.org/10.3390/foods14122117
Chicago/Turabian StyleYang, Lili, Chenyan Lv, Xingfeng Guo, and Rong Liang. 2025. "Donkey-Hide Gelatin Peptide-Iron Complexes: Structural Characterization, Enhanced Iron Solubility Under Simulated Digestion, and Dual Iron Chelation-Antioxidant Functions" Foods 14, no. 12: 2117. https://doi.org/10.3390/foods14122117
APA StyleYang, L., Lv, C., Guo, X., & Liang, R. (2025). Donkey-Hide Gelatin Peptide-Iron Complexes: Structural Characterization, Enhanced Iron Solubility Under Simulated Digestion, and Dual Iron Chelation-Antioxidant Functions. Foods, 14(12), 2117. https://doi.org/10.3390/foods14122117