

Optimization of Extraction Process and Activity of Angiotensin-Converting Enzyme (ACE) Inhibitory Peptide from Walnut Meal
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Defatted Walnut Meal
2.3. Preparation of Walnut Protein
2.4. Optimization of Preparation Process of Walnut Peptide by Enzymatic Hydrolysis
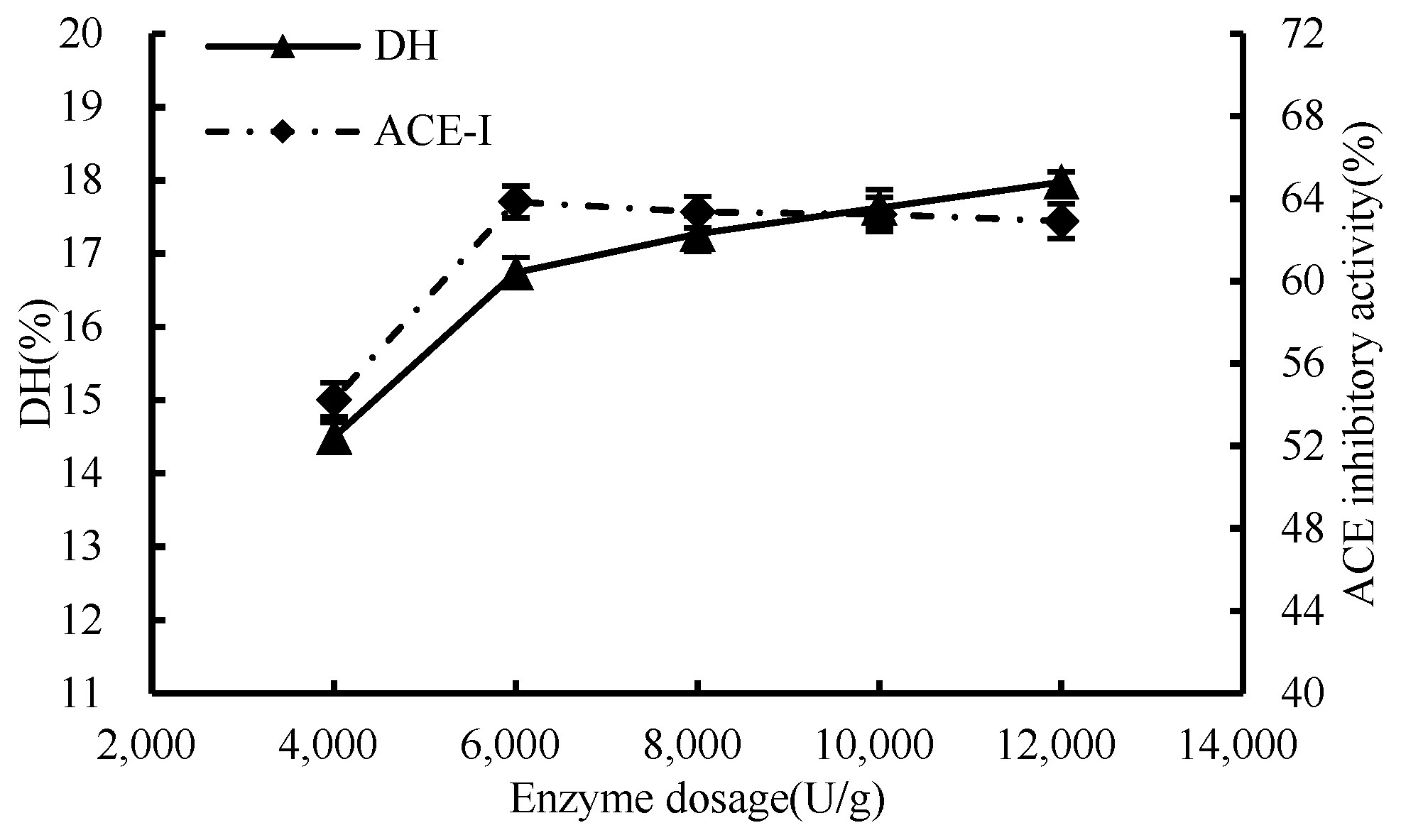
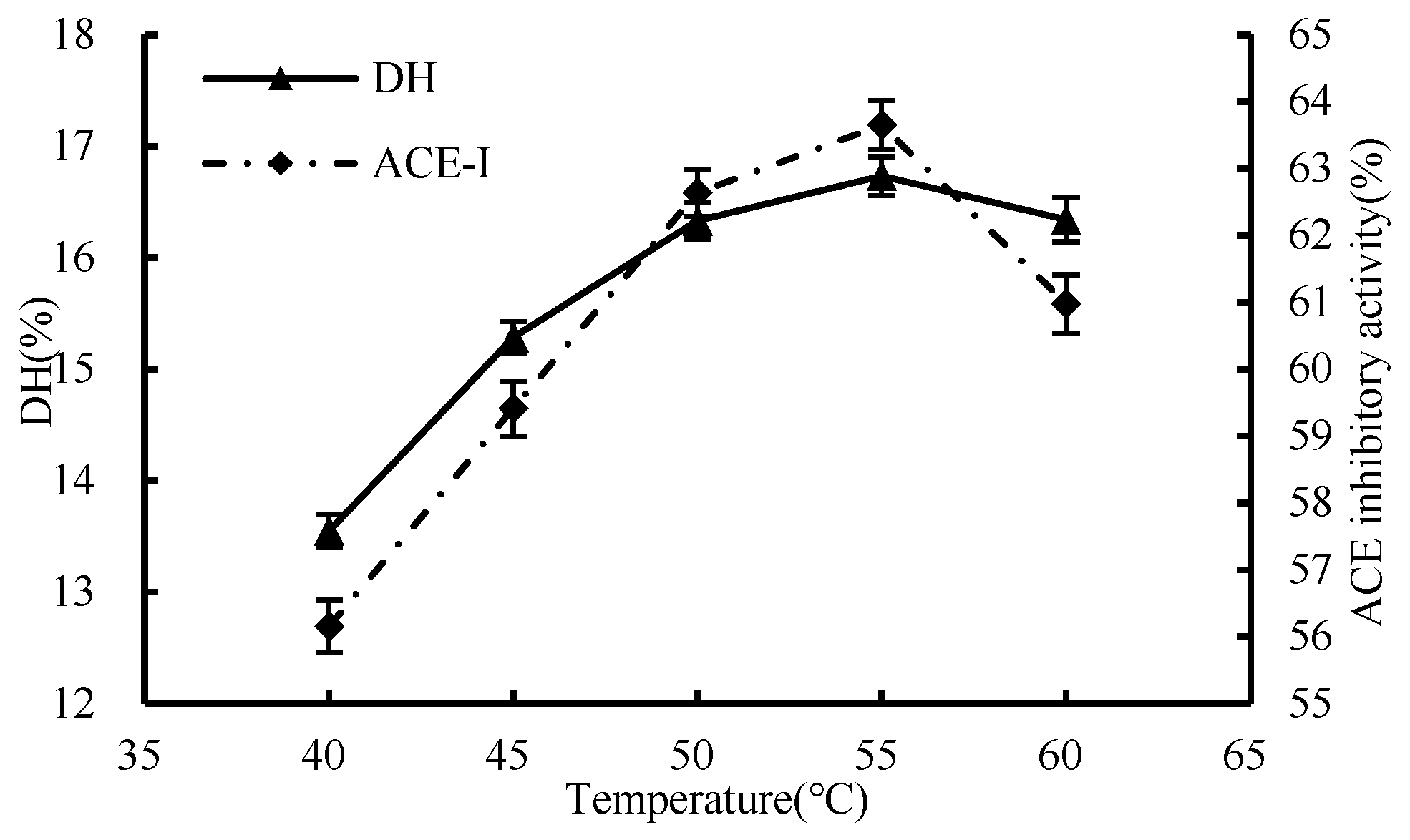
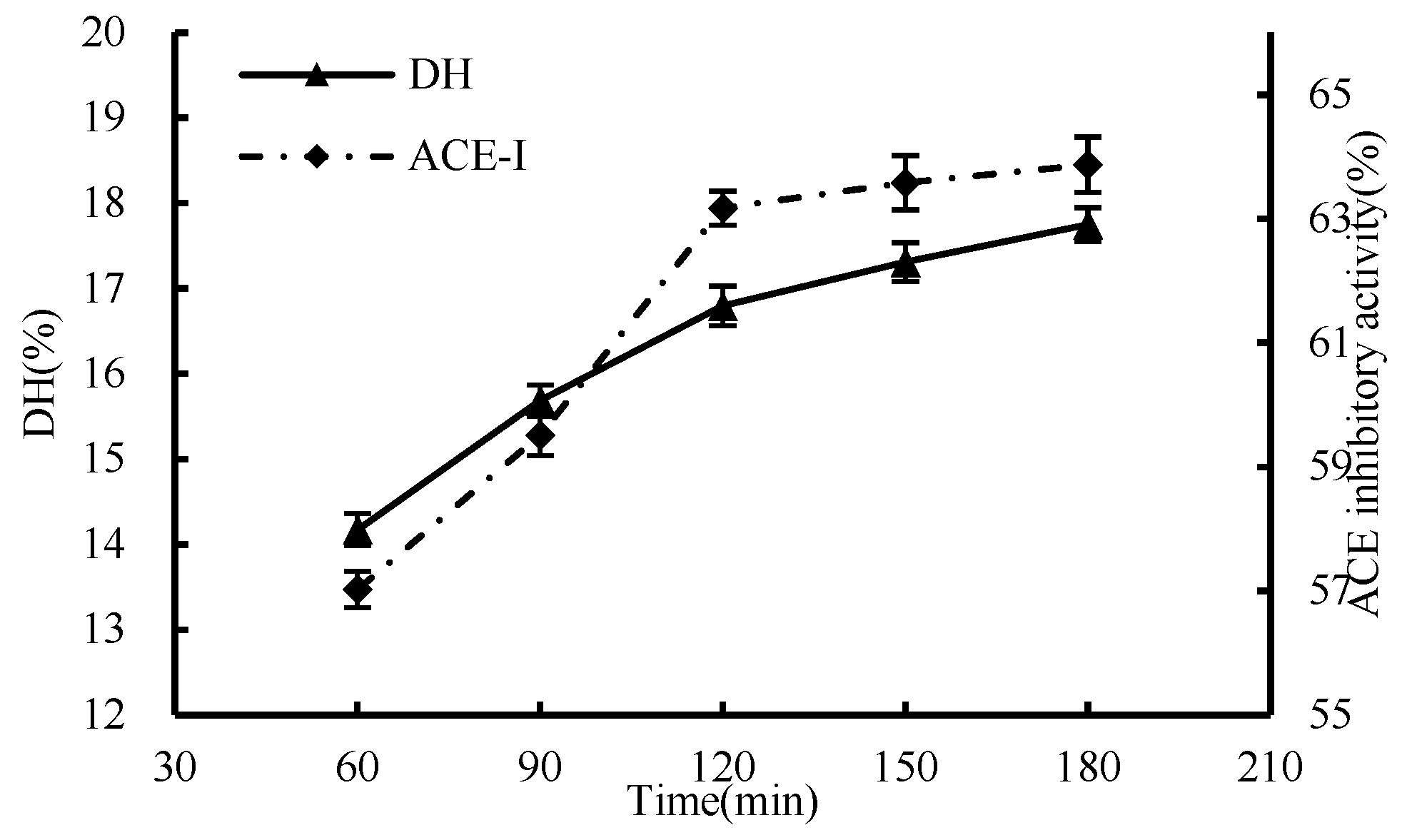
2.4.1. Single-Factor Test
2.4.2. Response Surface Optimization Method
2.5. Degree of Hydrolysis
2.6. Evaluation of ACE Inhibitory Activity
2.7. Ultrafiltration Separation
2.8. Determination of Antioxidant Capacity
2.8.1. ·OH Free Radical Scavenging Ability
2.8.2. DPPH Free Radical Scavenging Ability
2.8.3. O2− Scavenging Ability
2.8.4. Total Reducing Power
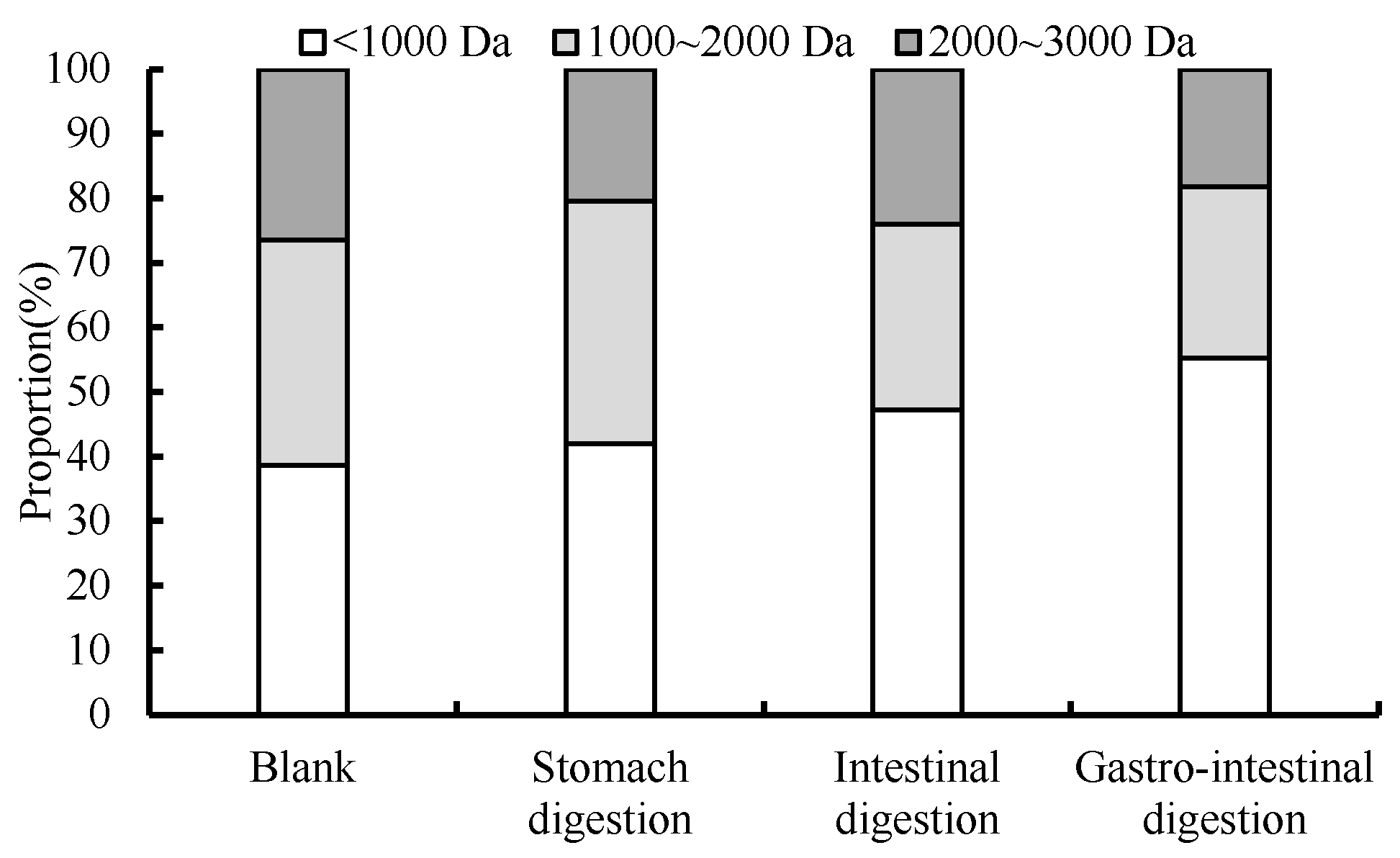
2.9. Simulating Gastrointestinal Digestion In Vitro
2.10. Determination of the Relative Molecular Mass of Peptides
3. Results
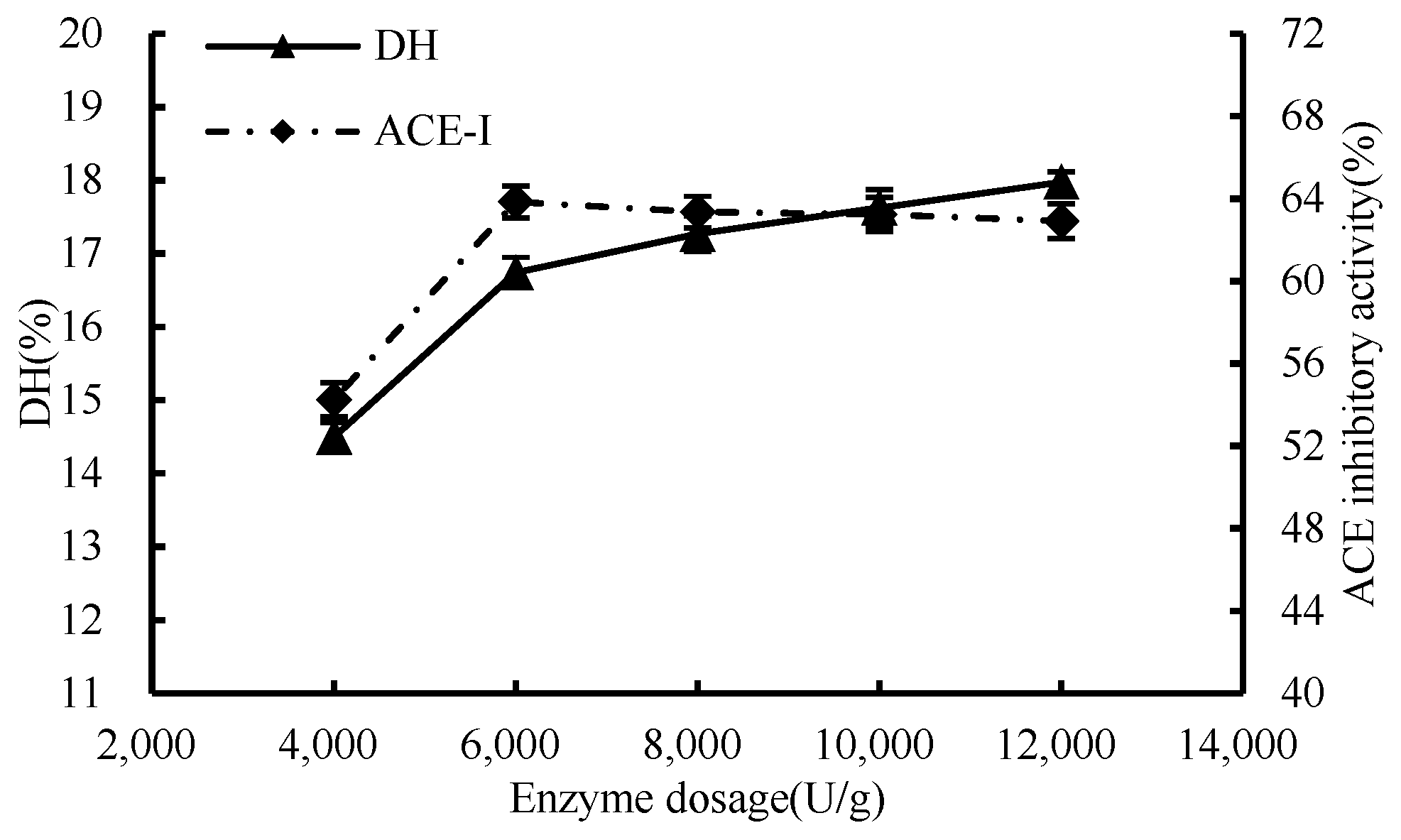
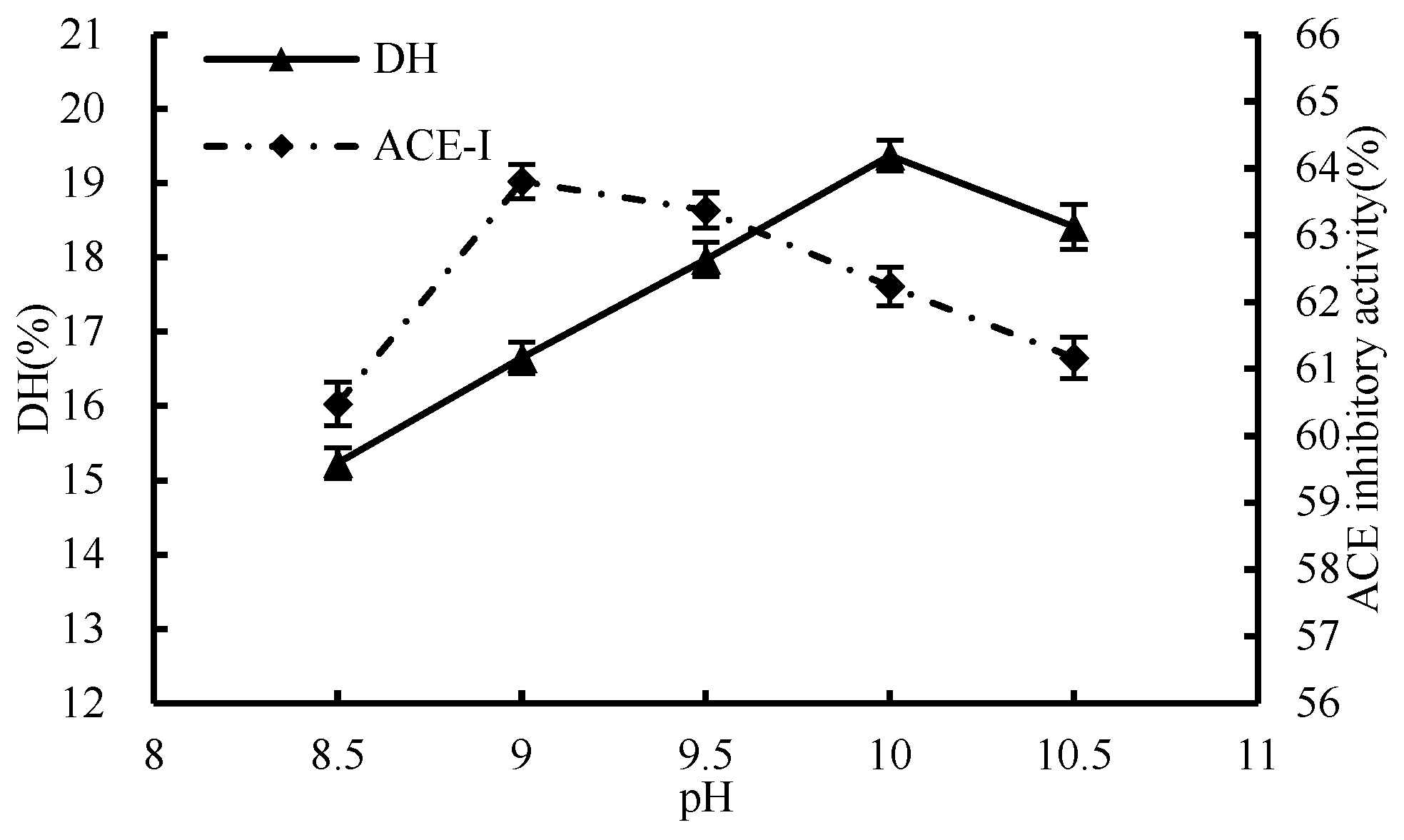
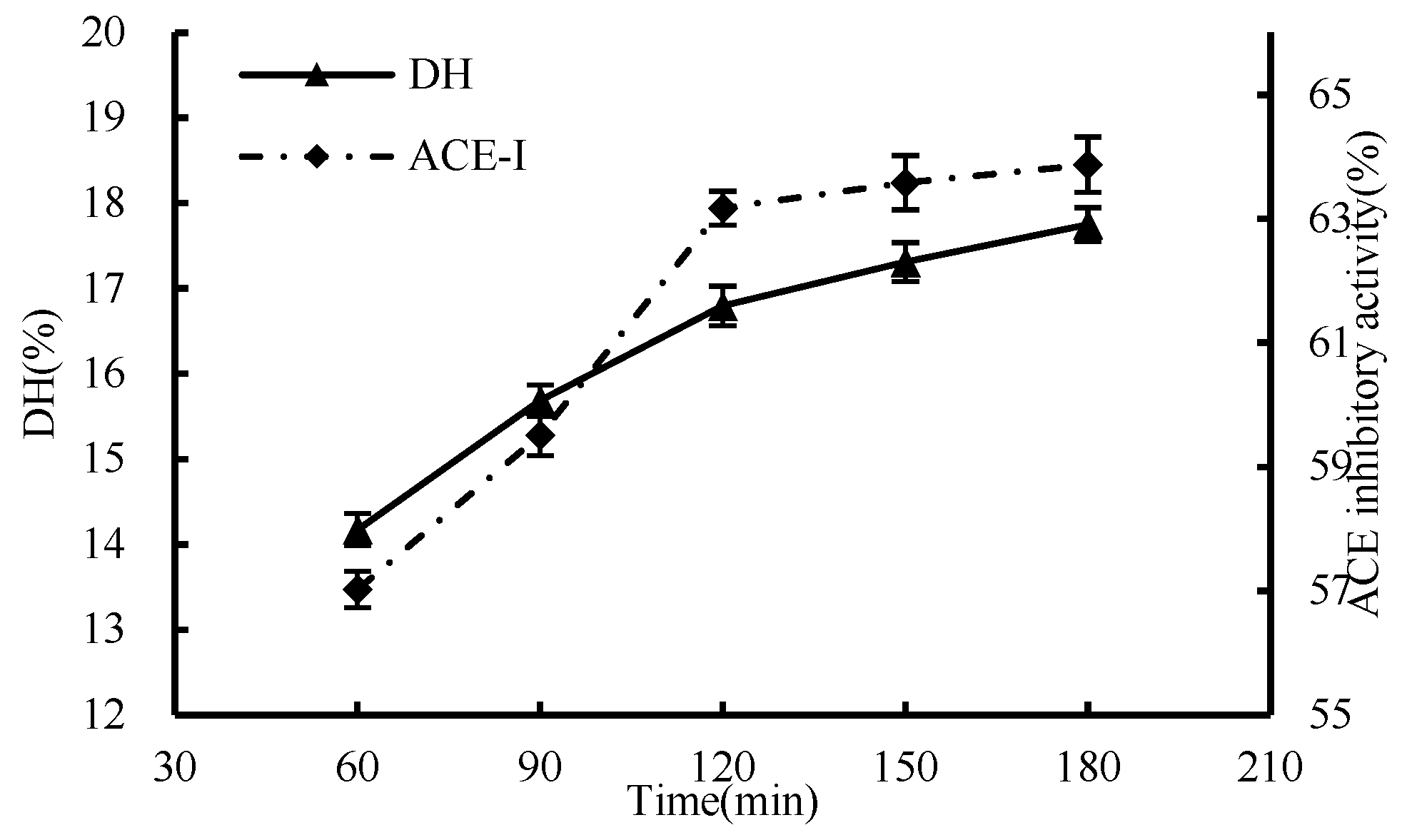
3.1. Single-Factor Test of Enzymatic Hydrolysis Process
3.1.1. Enzymatic Effect
3.1.2. Effect of Temperature
3.1.3. Effect of pH
3.1.4. Effect of Time
3.2. Response Surface Optimization of Enzymatic Hydrolysis Process
3.2.1. Response Surface Experiments—Design and Results
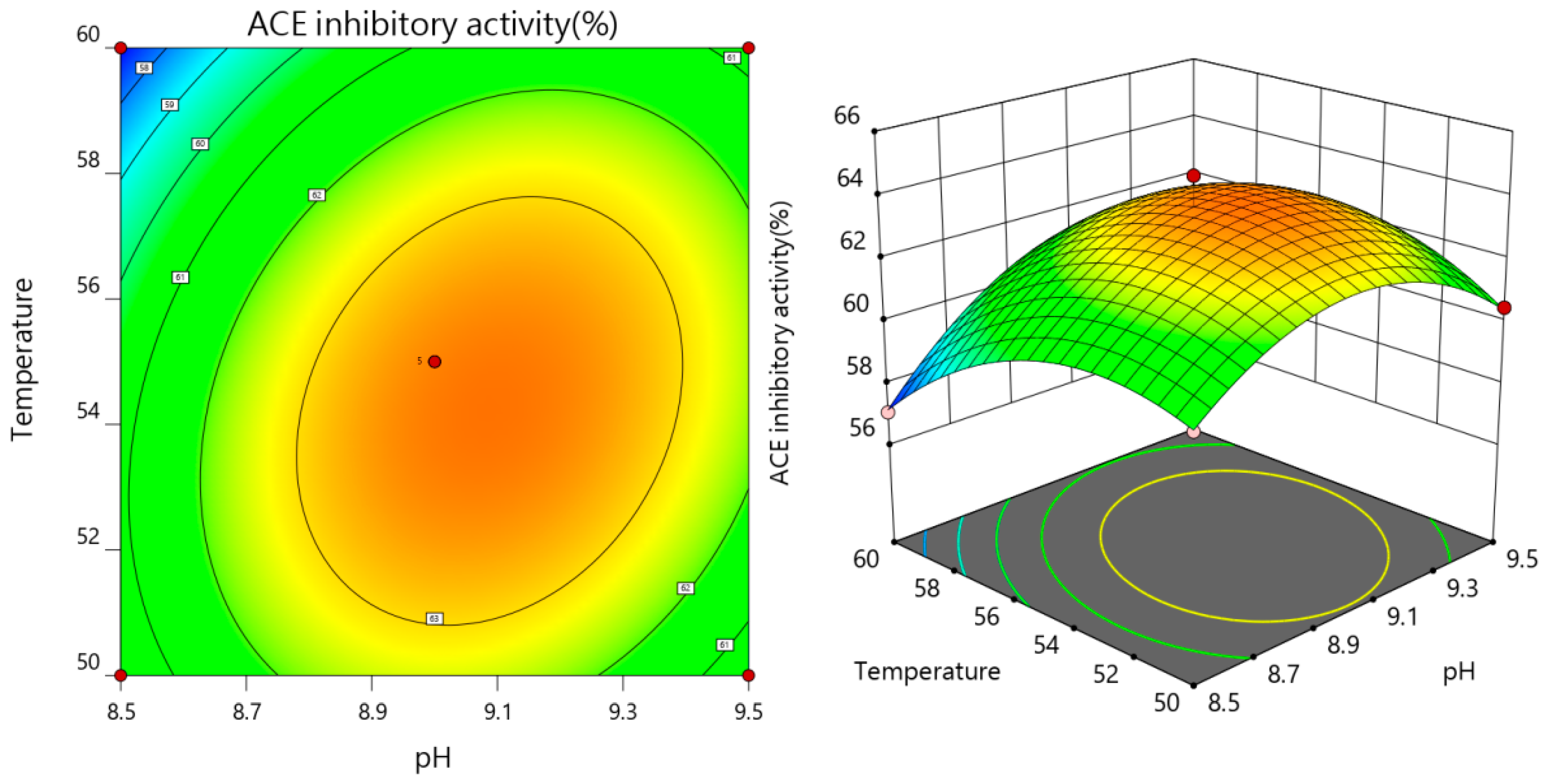
3.2.2. Response Surface Regression Equation
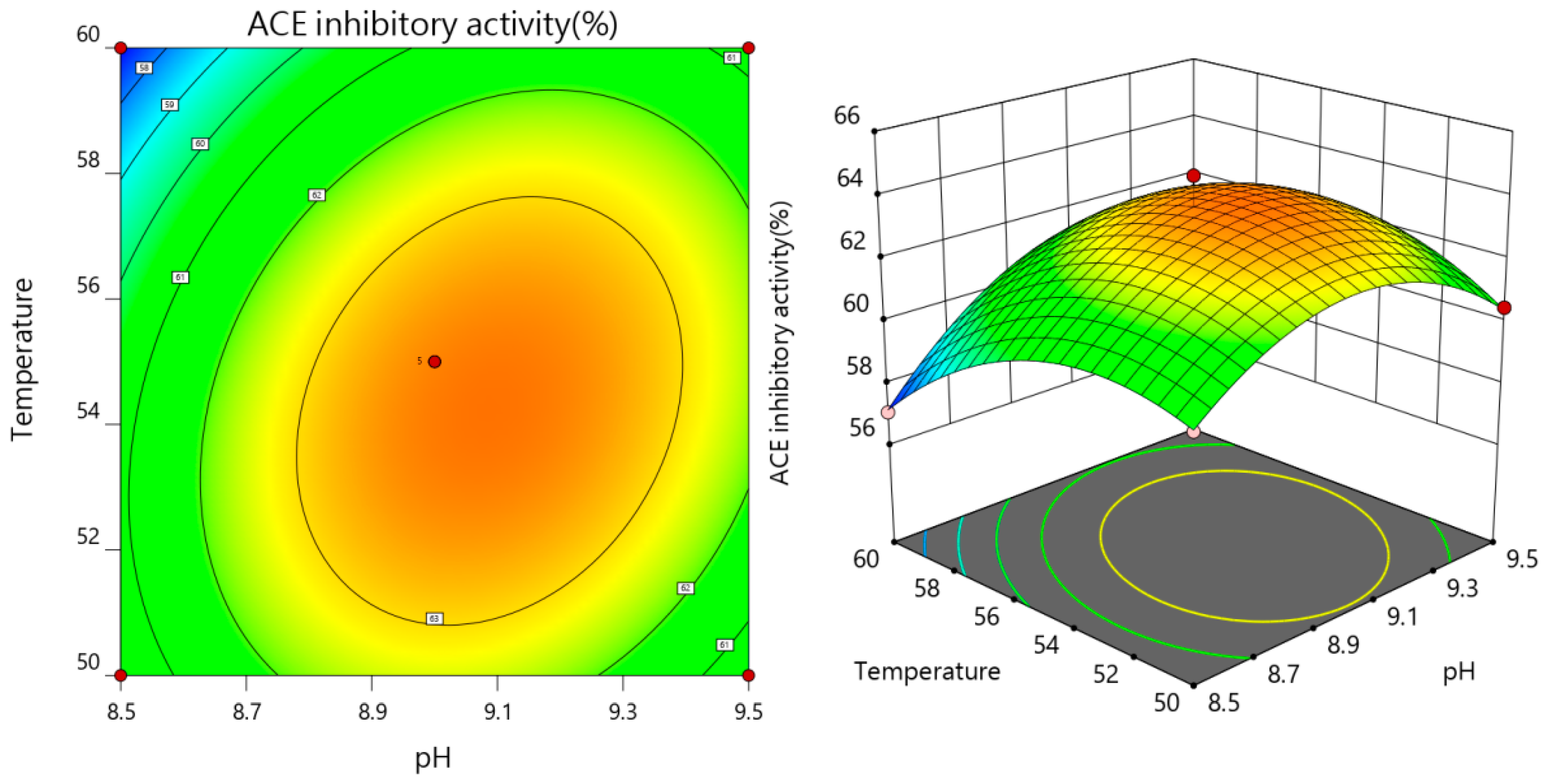
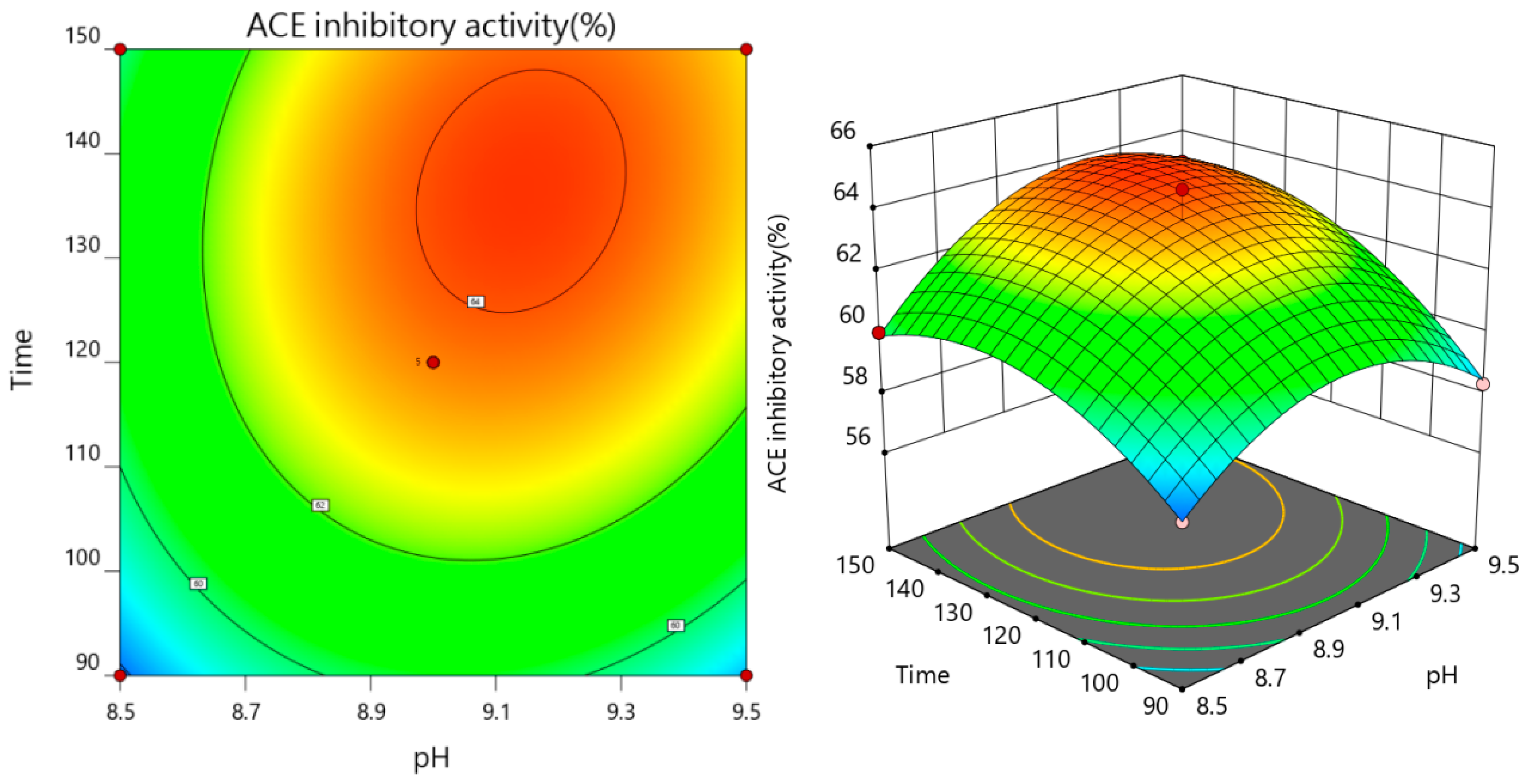
3.2.3. The Results of Interaction
3.2.4. Verification of the Optimal Enzymatic Hydrolysis Process
3.3. Ultrafiltration Separation
3.4. In Vitro Antioxidant Activity
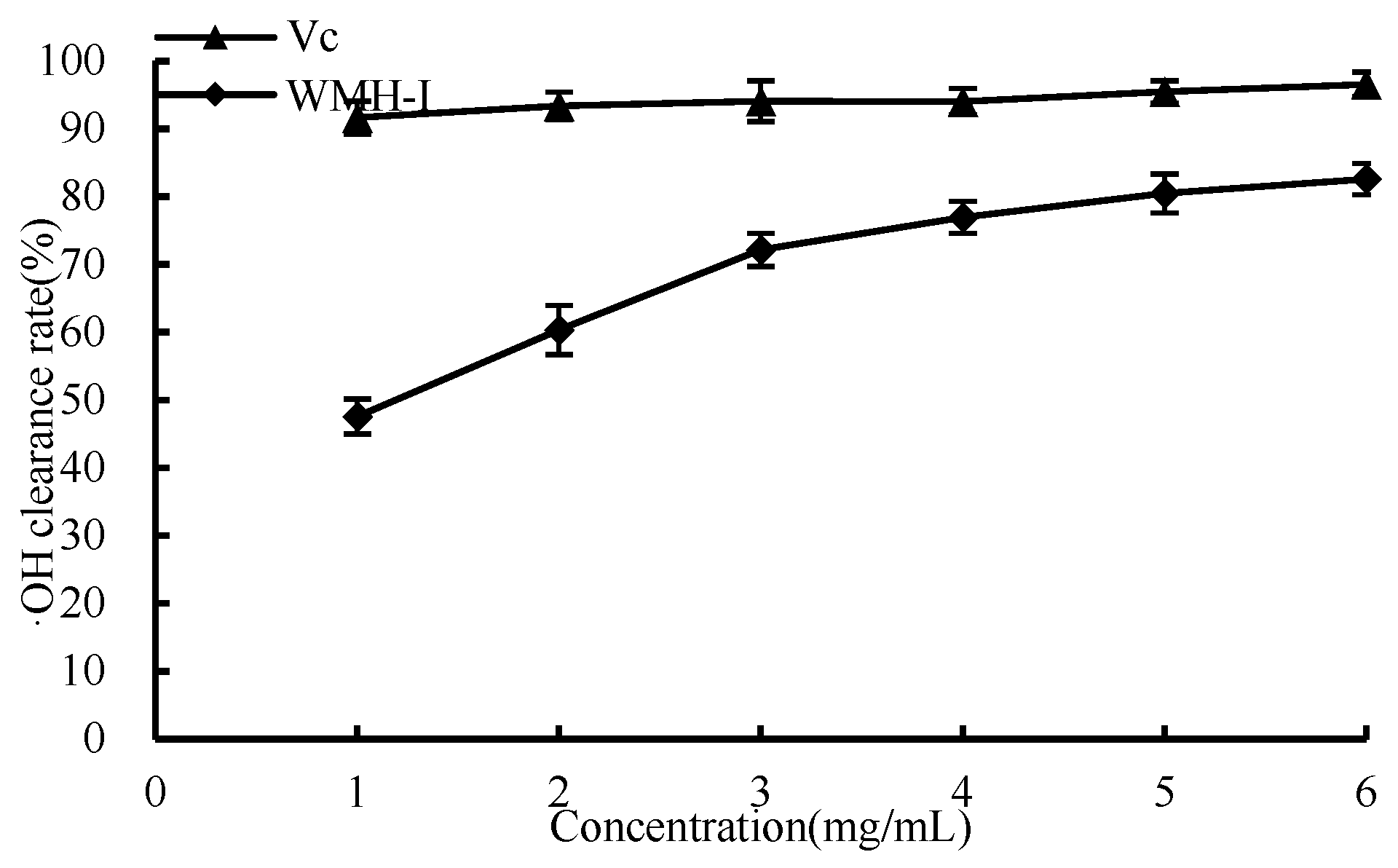
3.4.1. ·OH Scavenging Ability
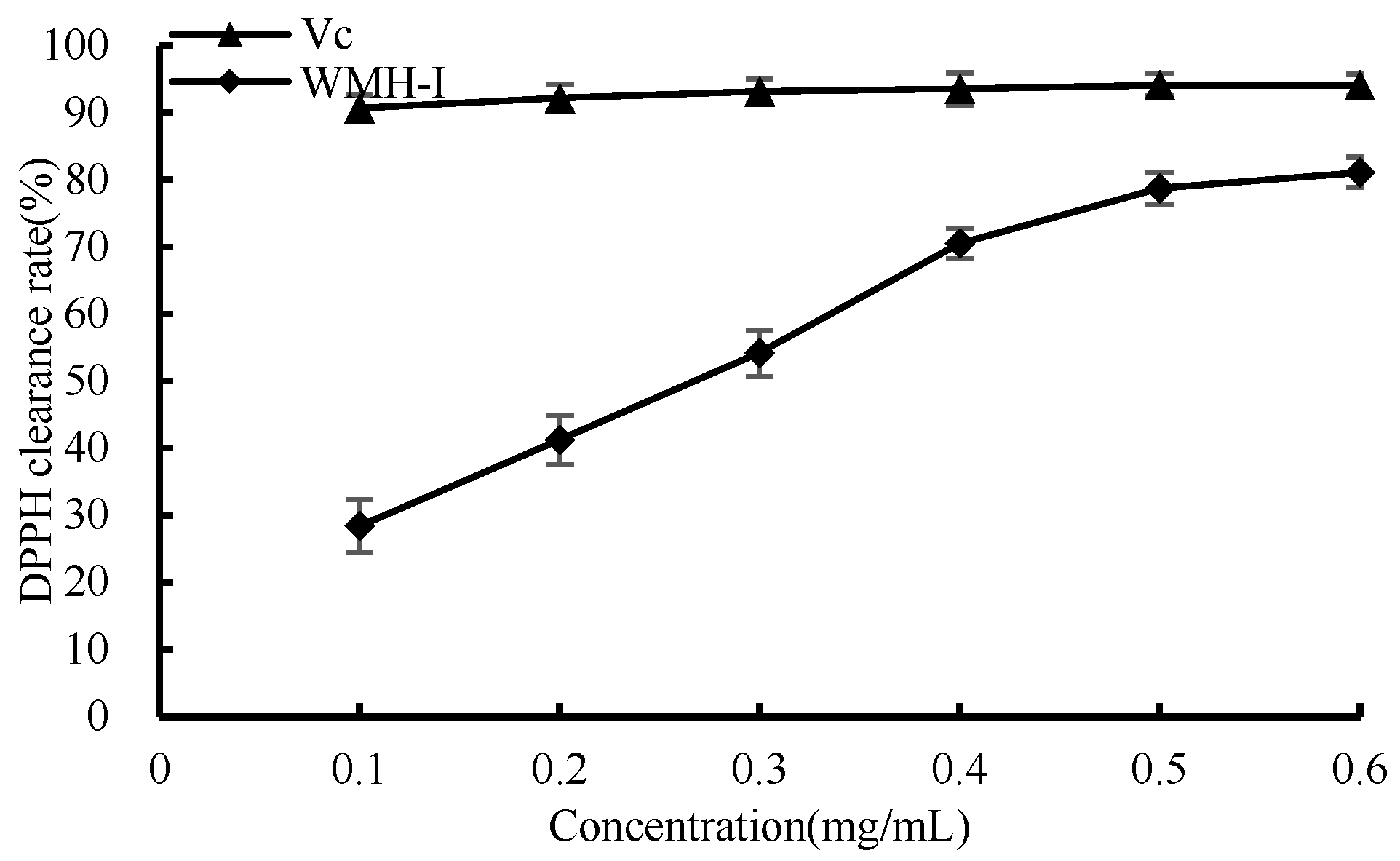
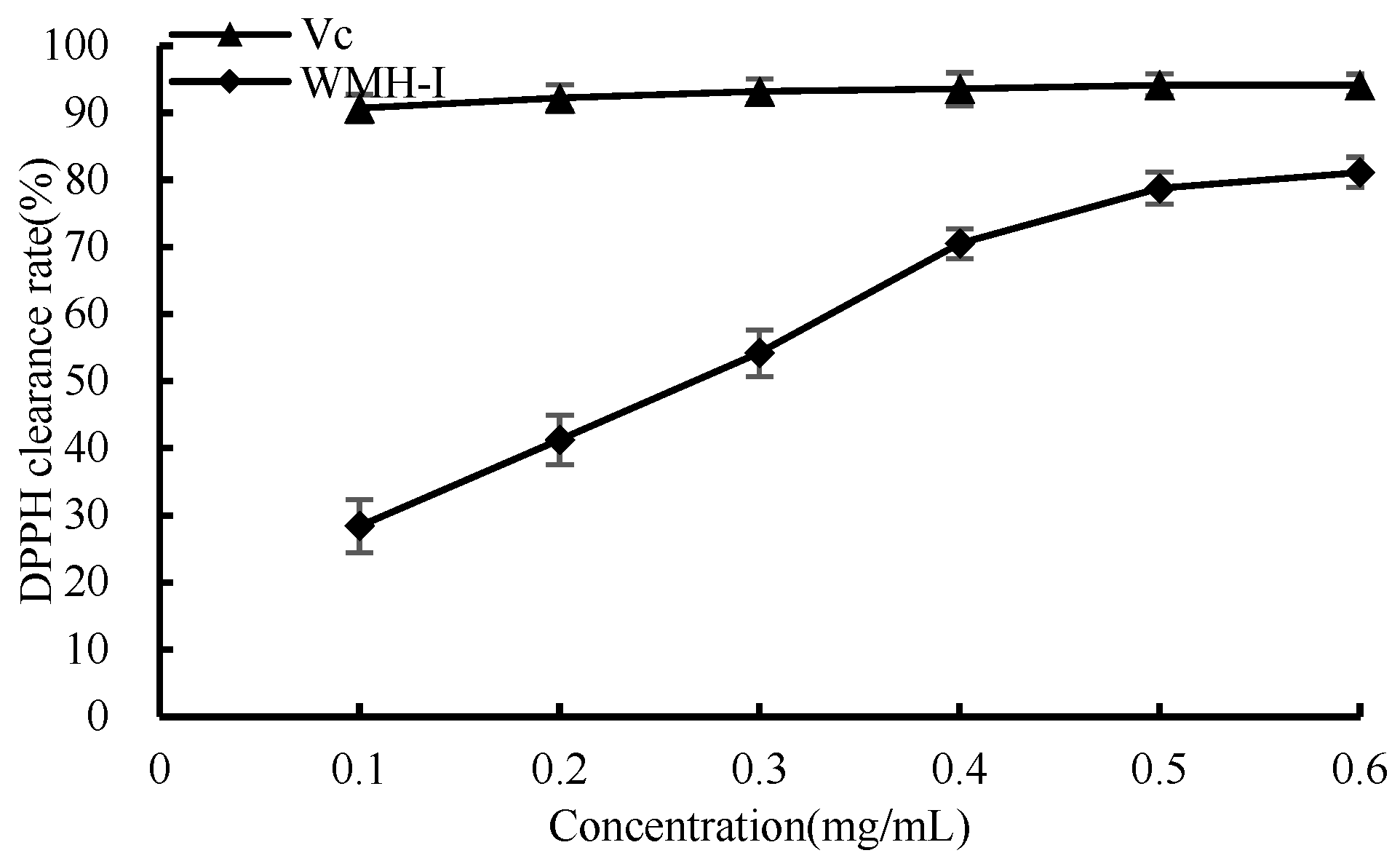
3.4.2. DPPH Scavenging Ability
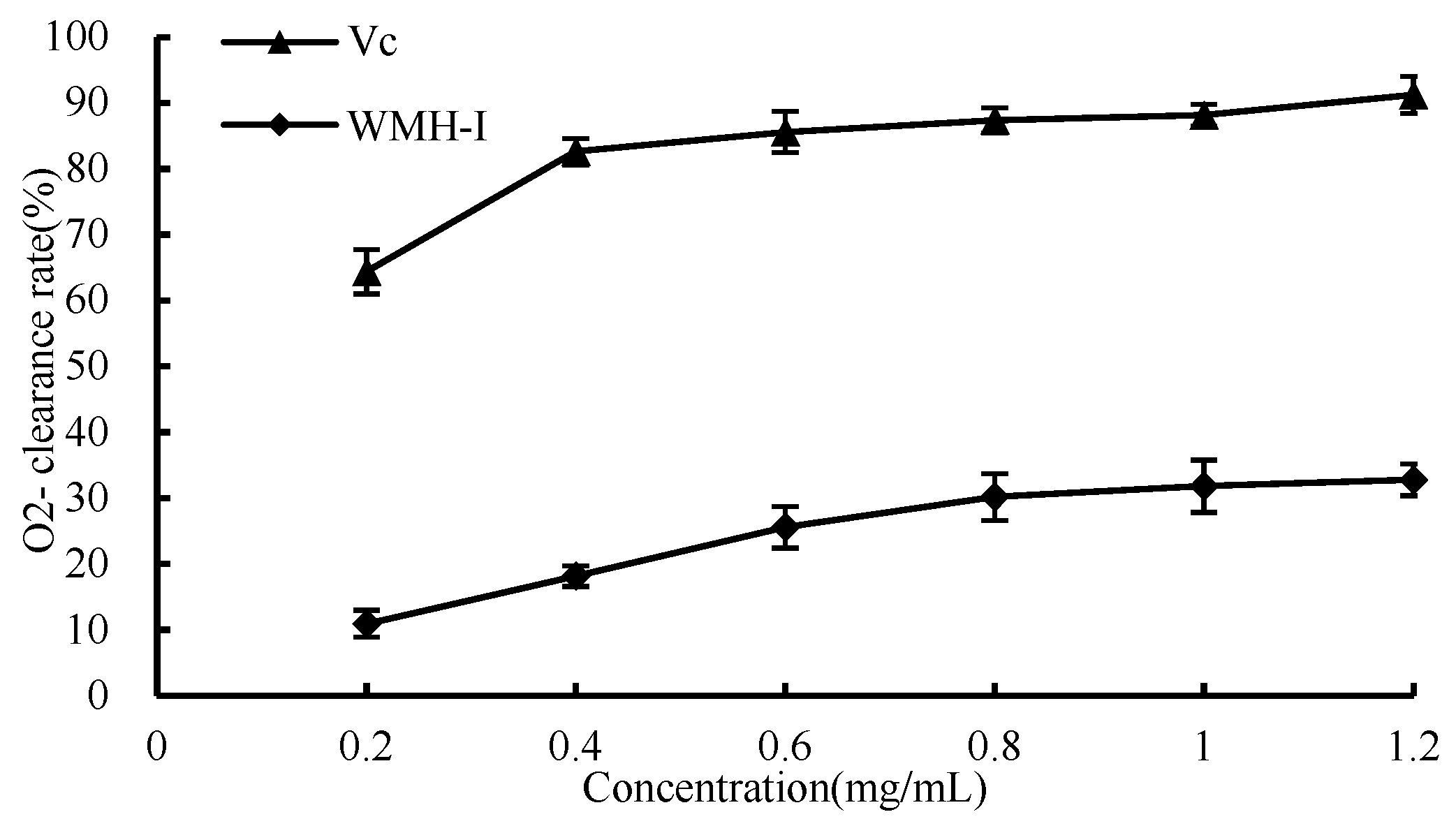
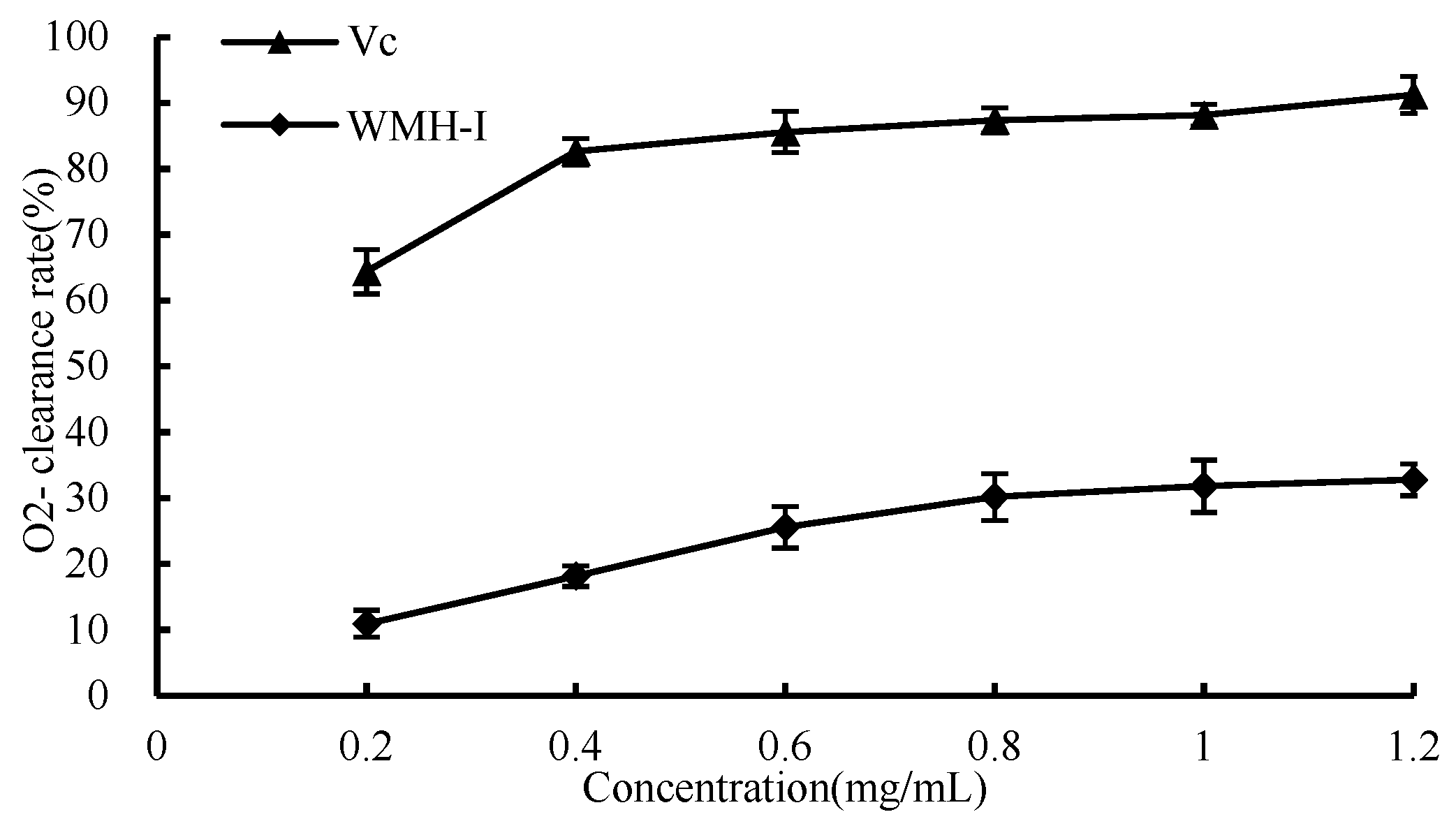
3.4.3. O2− Scavenging Ability
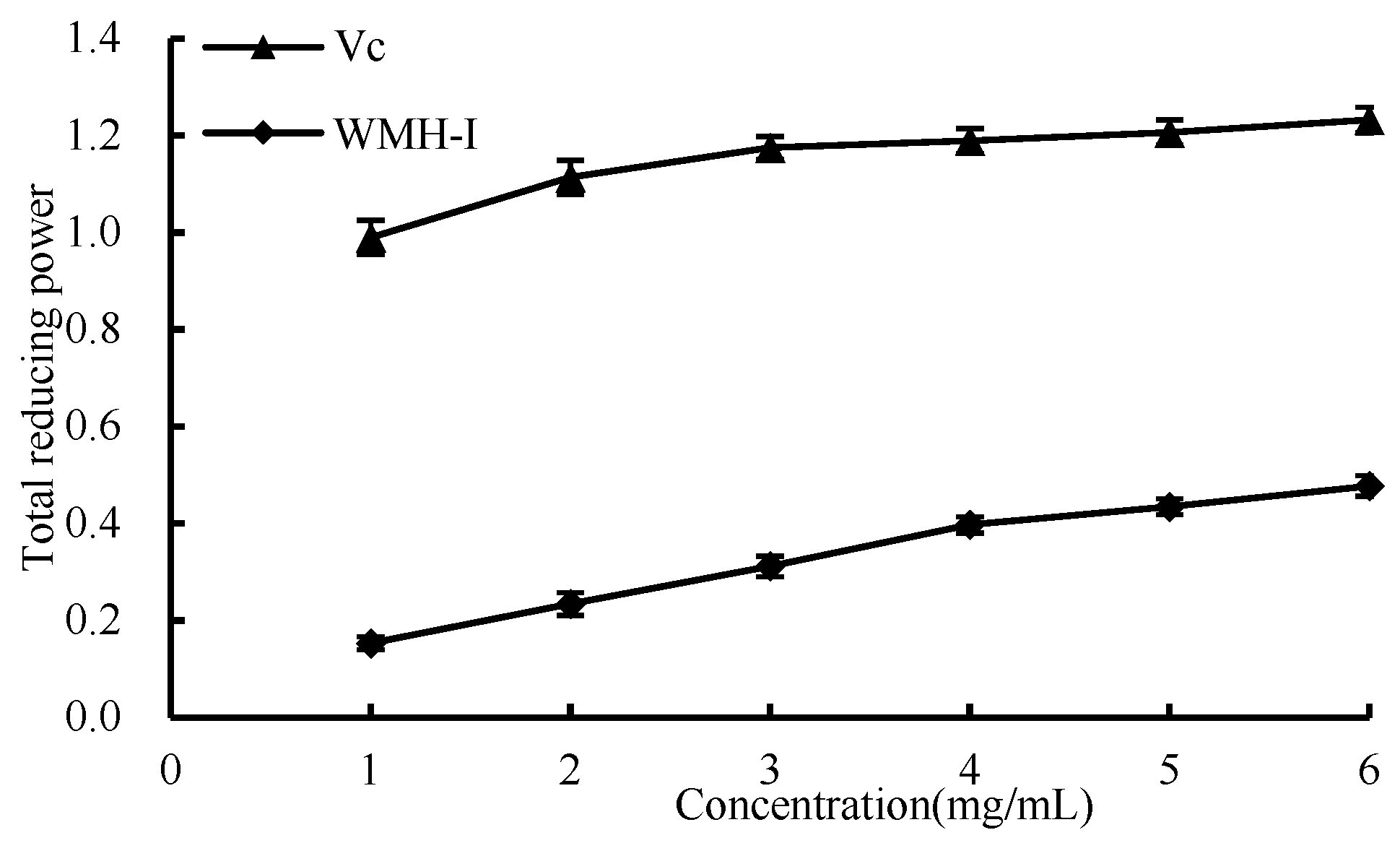
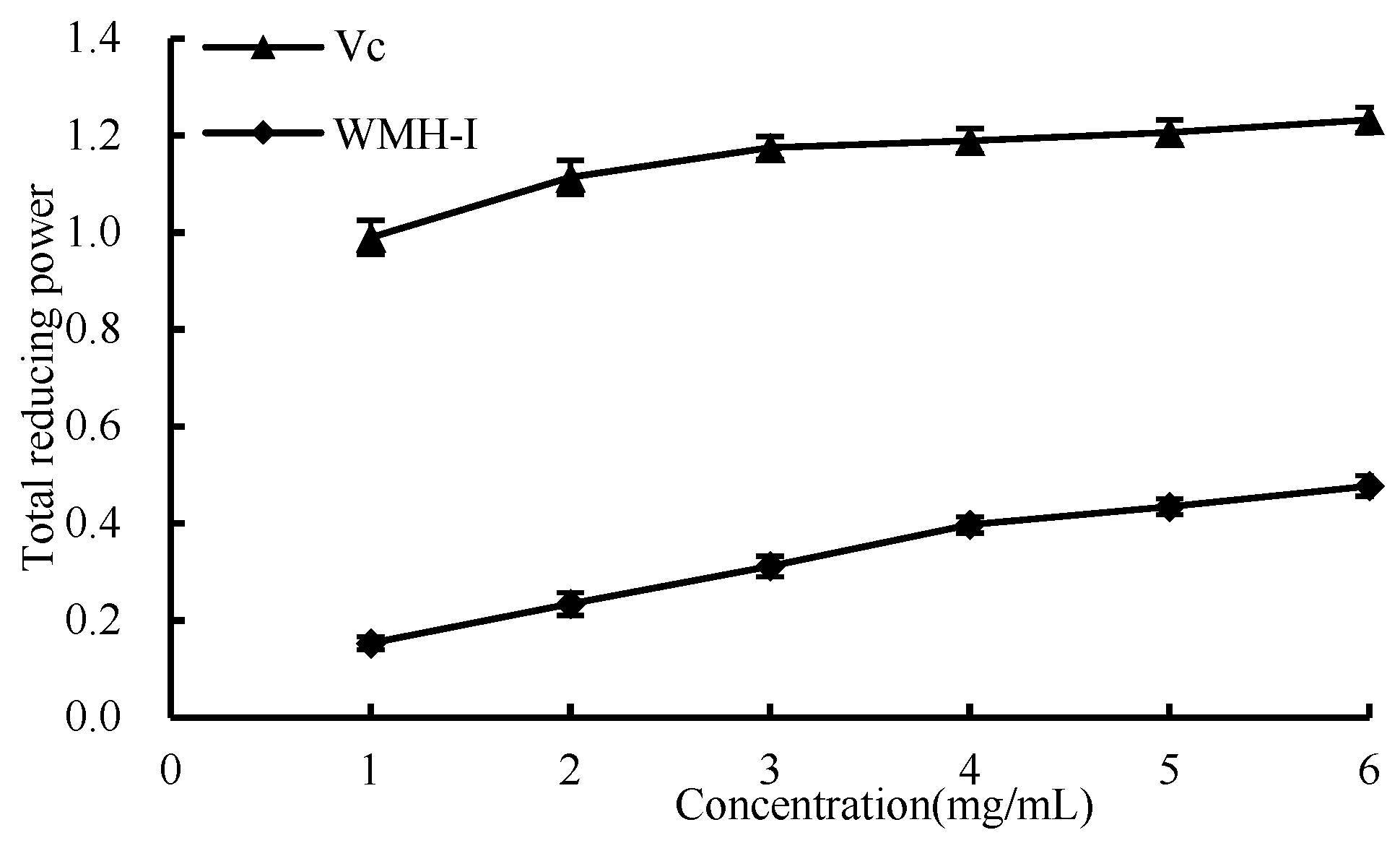
3.4.4. Total Reducing Power
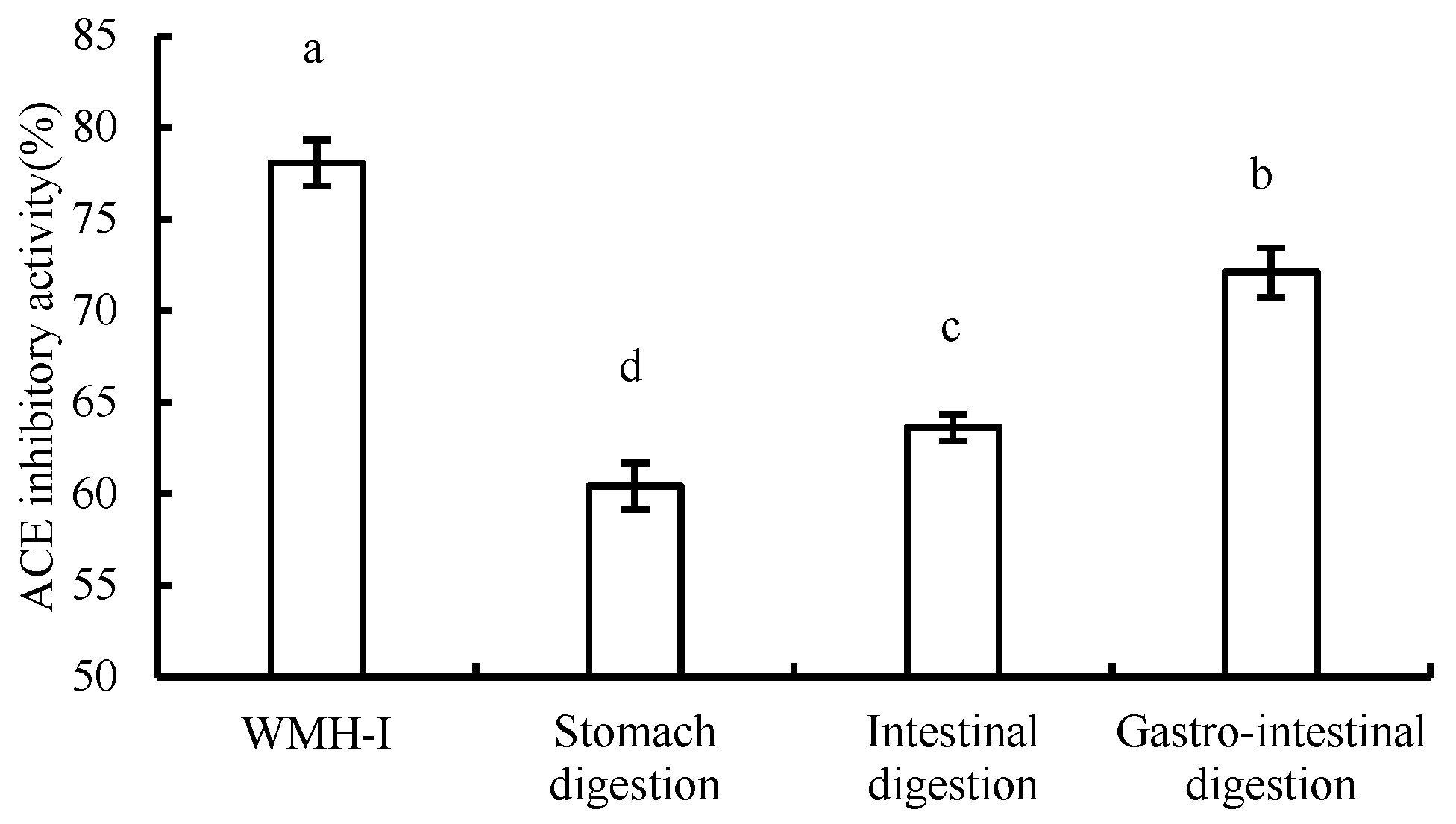
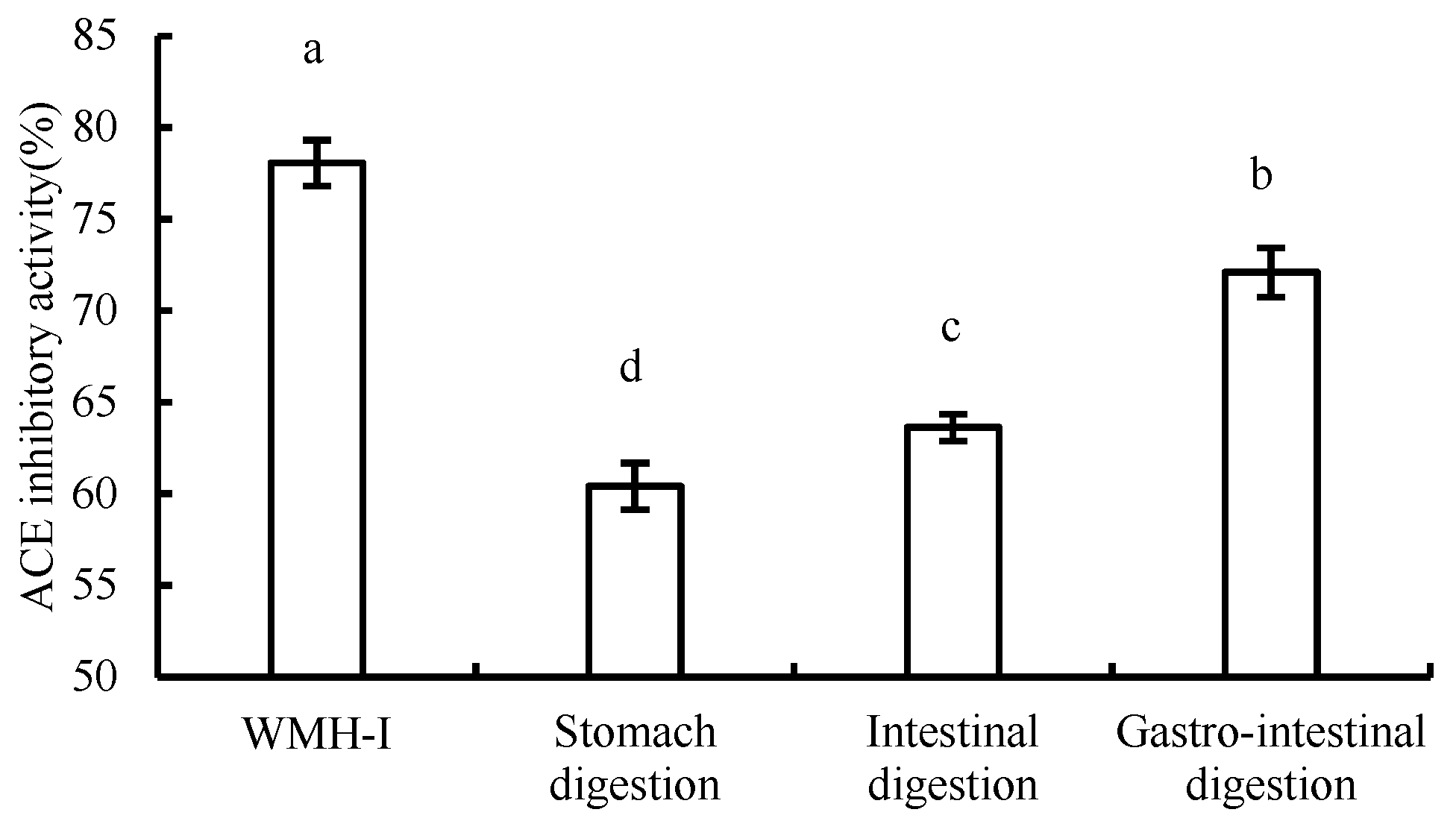
3.5. Changes in ACE Inhibition Rate during Digestion
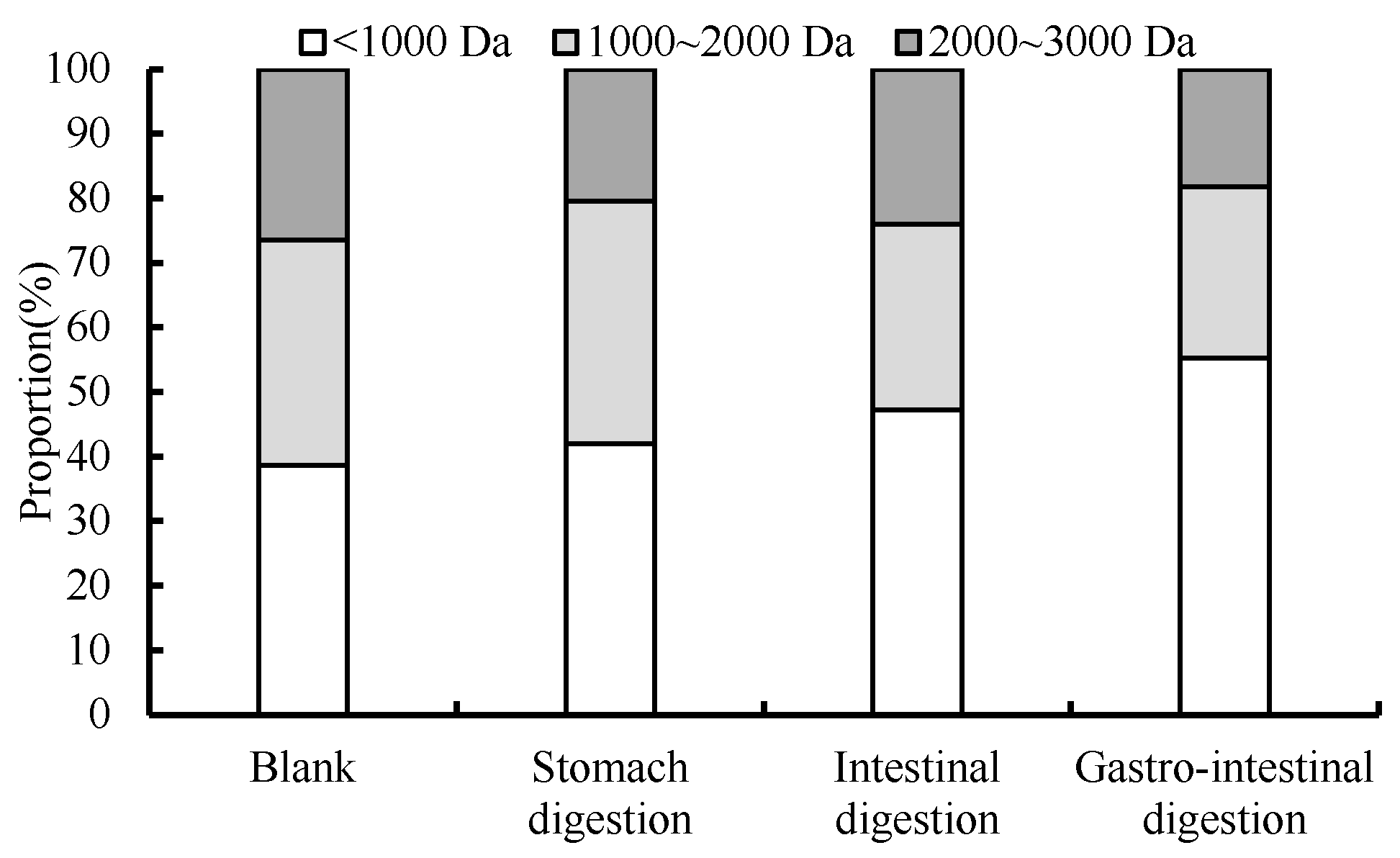
3.6. Changes in Molecular Mass Distribution during Digestion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.; Zhang, Z.; Shang, Y.; Li, S.; Xia, J.; Tian, Y.; Jia, Y.; Ma, A. Progress in the preparation, identification and biological activity of walnut peptides. J. Future Foods 2024, 4, 205–220. [Google Scholar] [CrossRef]
- Xie, J.; Hong, Z.; Dai, J.; Li, T.; Bai, Y.; Zhang, L.; Hu, X.; Chen, J.; Sheng, J.; Tian, Y. Isolation and identification of anti-colorectal cancer peptides from walnut proteins and associated in silico analysis. J. Funct. Foods 2024, 112, 105952. [Google Scholar] [CrossRef]
- Liu, K.L.; Gao, Z.R.; Li, Q.Y.; Zhang, H.Y. Identification and mechanistic study of four novel ACE inhibitory peptides from maize germ protein hydrolysates. LWT 2023, 186, 115254. [Google Scholar] [CrossRef]
- Duan, X.J.; Dong, Y.F.; Zhang, M.; Li, Z.H.; Bu, G.H.; Chen, F.S. Identification and molecular interactions of novel ACE inhibitory peptides from rapeseed protein. Food Chem. 2023, 422, 136085. [Google Scholar] [CrossRef] [PubMed]
- Asoodeh, A.; Yazdi, M.M.; Chamani, J. Purification and characterisation of angiotensin I converting enzyme inhibitory peptides from lysozyme hydrolysates. Food Chem. 2012, 131, 291–295. [Google Scholar] [CrossRef]
- Xue, L.; Yin, R.; Howell, K.; Zhang, P. Activity and bioavailability of food protein-derived angiotensin-I-converting enzyme–inhibitory peptides. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1150–1187. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hur, S.J. Antihypertensive peptides from animal products, marine organisms, and plants. Food Chem. 2017, 228, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.; Mora, L.; Toldrá, F. Health relevance of antihypertensive peptides in foods. Curr. Opin. Food Sci. 2018, 19, 8–14. [Google Scholar] [CrossRef]
- Shao, B.; Huang, X.Y.; Xu, M.D.; Cheng, D.W.; Li, X.L.; Li, M.Q. Peptides isolated from black soybean synergistically inhibit the activity of angiotensin converting enzyme (ACE). J. Funct. Foods 2023, 106, 105604. [Google Scholar] [CrossRef]
- Zhang, J.; Li, M.; Zhang, G.; Tian, Y.; Kong, F.; Xiong, S.; Zhao, S.; Jia, D.; Manyande, A.; Du, H. Identification of novel antioxidant peptides from snakehead (Channa argus) soup generated during gastrointestinal digestion and insights into the anti-oxidation mechanisms. Food Chem. 2021, 337, 127921. [Google Scholar] [CrossRef]
- Lin, Z.L.; Wu, H.; Zhang, M.M. Isolation, identification, and structure-activity relationship of novel ACE inhibitory peptides from earthworm protein in vitro gastrointestinal digestion product. Food Biosci. 2023, 55, 103010. [Google Scholar] [CrossRef]
- Liu, M.; Du, M.; Zhang, Y.; Xu, W.; Wang, C.; Wang, K.; Zhang, L. Purification and Identification of an ACE Inhibitory Peptide from Walnut Protein. J. Agric. Food Chem. 2013, 61, 4097–4100. [Google Scholar] [CrossRef] [PubMed]
- Obeng, E.; Ding, F.; He, X.J.; Shen, J.L. Bioimaging of superoxide anions in living cells and Perfect visualization with fluorescence probes and their applications. Dye. Pigment. 2022, 199, 109964. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, C.; Sun-Waterhouse, D.; Zhao, T.; Waterhouse, G.I.N.; Zhao, M.; Su, G. Identification of post-digestion angiotensin-I converting enzyme (ACE) inhibitory peptides from soybean protein Isolate: Their production conditions and in silico molecular docking with ACE. Food Chem. 2021, 345, 128855. [Google Scholar] [CrossRef] [PubMed]
- Magouz, O.; Mehanna, N.; Khalifa, M.; Sakr, H.; Gensberger-Reigl, S.; Dalabasmaz, S.; Pischetsrieder, M. Profiles, antioxidative and ACE inhibitory activity of peptides released from fermented buttermilk before and after simulated gastrointestinal digestion. Innov. Food Sci. Emerg. Technol. 2023, 84, 103266. [Google Scholar] [CrossRef]
- Escudero, E.; Mora, L.; Toldrá, F. Stability of ACE inhibitory ham peptides against heat treatment and digestion. Food Chem. 2014, 161, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.J.; Li, S.J.; Gu, Y.; Wang, S.; Wang, J.P. Effects of Starch on the Digestibility of Gluten under Different Thermal Processing Conditions. J. Agric. Food Chem. 2019, 67, 7120–7127. [Google Scholar] [CrossRef] [PubMed]
- Garzón, A.G.; Veras, F.F.; Brandelli, A.; Drago, S.R. Purification, identification and in silico studies of antioxidant, antidiabetogenic and antibacterial peptides obtained from sorghum spent grain hydrolysate. LWT 2022, 153, 112414. [Google Scholar] [CrossRef]
- Silvestre, M.P.C.; Silva, M.R.; Silva, V.D.M.; de Souza, M.W.S.; Lopes, C.D.; Afonso, W.D. Analysis of whey protein hydrolysates: Peptide profile and ACE inhibitory activity. Braz. J. Pharm. Sci. 2012, 48, 747–757. [Google Scholar] [CrossRef]
- Hasan, M.R.; Haque, M.M.; Hoque, M.A.; Sultana, S.; Rahman, M.M.; Ali Shaikh, M.A.; Sarker, M.K.U. Antioxidant activity study and GC-MS profiling of Camellia sinensis Linn. Heliyon 2024, 10, e23514. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Li, P.; Liu, J.; Su, D.; Tang, B. Two-photon fluorescence imaging reveals a Golgi apparatus superoxide anion-mediated hepatic ischaemia-reperfusion signalling pathway. Chem. Sci. 2019, 10, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, D.T.; Valentine, J.S. How super is superoxide? Acc. Chem. Res. 1981, 14, 393–400. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Laursen, J.B.; Somers, M.; Kurz, S.; McCann, L.; Warnholtz, A.; Freeman, B.A.; Tarpey, M.; Fukai, T.; Harrison, D.G.J.C. Endothelial regulation of vasomotion in apoE-deficient mice: Implications for interactions between peroxynitrite and tetrahydrobiopterin. Circulation 2001, 103, 1282–1288. [Google Scholar] [CrossRef]
- Bitar, M.S.; Wahid, S.; Mustafa, S.; Al-Saleh, E.; Dhaunsi, G.S.; Al-Mulla, F. Nitric oxide dynamics and endothelial dysfunction in type II model of genetic diabetes. Eur. J. Pharmacol. 2005, 511, 53–64. [Google Scholar] [CrossRef]
- Hu, X.T.; Xu, F.R.; Li, J.L.; Li, J.; Mo, C.; Zhao, M.; Wang, L.F. Ultrasonic-assisted extraction of polysaccharides from coix seeds: Optimization, purification, and digestibility. Food Chem. 2022, 374, 131636. [Google Scholar] [CrossRef] [PubMed]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef]
- Li, X.; Wei, L.; Jia, L.; Li, M.; Zhu, L.; Liu, L.; Gao, Y.J.P. Identification and characterization of cow’s milk proteins from the rat intestinal lymph using a proteomic strategy. Proteomics 2013, 13, 2649–2656. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Influencing Factors | Level Setting | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Amount of enzyme added (U/g) | 2000 | 4000 | 6000 | 8000 | 10,000 |
| Temperature (°C) | 40 | 45 | 50 | 55 | 60 |
| pH | 8.5 | 9 | 9.5 | 10 | 10.5 |
| Time (min) | 60 | 90 | 120 | 150 | 180 |
| Influencing Factors | Level Setting | |||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| A | pH | 8.5 | 9 | 9.5 |
| B | Temperature (°C) | 50 | 55 | 60 |
| C | Time (min) | 60 | 120 | 150 |
| Reagent | Amount Added (μL) | ||
|---|---|---|---|
| Experimental | Control | Blank | |
| 1 mol/L HCl | 0 | 0 | 100 |
| HHL | 30 | 30 | 30 |
| Peptide | 10 | 0 | 10 |
| Buffer | 0 | 10 | 0 |
| Operation 1 | A water bath at 37 °C for 5 min | ||
| ACE | 10 | 10 | 10 |
| Operation 2 | A water bath at 37 °C for 30 min | ||
| 1 mol/L HCl | 100 | 100 | 0 |
| Test No. | A pH | B Temperature | C Time | Y ACE Inhibition Rate (%) |
|---|---|---|---|---|
| 1 | 0 | 0 | 0 | 64.64 |
| 2 | −1 | 0 | 1 | 60.03 |
| 3 | 0 | 0 | 0 | 63.34 |
| 4 | 1 | 0 | −1 | 58.32 |
| 5 | −1 | 1 | 0 | 57.06 |
| 6 | −1 | −1 | 0 | 60.26 |
| 7 | 0 | −1 | 1 | 62.18 |
| 8 | 0 | 0 | 0 | 63.53 |
| 9 | 0 | −1 | −1 | 59.63 |
| 10 | 1 | 1 | 0 | 60.74 |
| 11 | 1 | −1 | 0 | 60.48 |
| 12 | −1 | 0 | −1 | 57.83 |
| 13 | 0 | 0 | 0 | 63.66 |
| 14 | 0 | 0 | 0 | 63.16 |
| 15 | 1 | 0 | 1 | 62.96 |
| 16 | 0 | 1 | 1 | 61.37 |
| 17 | 0 | 1 | −1 | 57.64 |
| Source | Sum of Square | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 88.58 | 9 | 9.84 | 49.21 | <0.0001 ** |
| A-pH | 6.70 | 1 | 6.70 | 33.49 | 0.0007 ** |
| B-temperature | 4.12 | 1 | 4.12 | 20.59 | 0.0027 ** |
| C-time | 21.52 | 1 | 21.52 | 107.59 | <0.0001 ** |
| AB | 2.99 | 1 | 2.99 | 14.96 | 0.0061 ** |
| AC | 1.49 | 1 | 1.49 | 7.44 | 0.0294 * |
| BC | 0.3481 | 1 | 0.3481 | 1.74 | 0.2286 |
| A2 | 20.85 | 1 | 20.85 | 104.27 | <0.0001 ** |
| B2 | 13.73 | 1 | 13.73 | 68.63 | <0.0001 ** |
| C2 | 11.54 | 1 | 11.54 | 57.70 | 0.0001 ** |
| Residual | 1.40 | 7 | 0.20 | ||
| Lack of Fit | 0.0704 | 3 | 0.0235 | 0.0707 | 0.9726 |
| Pure Error | 1.33 | 4 | 0.3324 | ||
| Cor Total | 89.98 | 16 | |||
| R2 = 0.9844 Adj R2 = 0.9644 C.V.% = 0.73 | |||||
| Group | Molecular Weight (Da) | Rate of Yield (%) | IC50 (mg/mL) |
|---|---|---|---|
| WMH | Stoste | 100 | 0.401 b |
| WMH-Ι | <3000 | 26.16 | 0.299 a |
| WMH-ΙΙ | 3000~10,000 | 39.82 | 0.652 c |
| WMH-ΙΙΙ | >10,000 | 32.48 | 1.518 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, M.; She, Z.; Feng, Y.; Zhang, J.; Han, R.; Qi, Y.; Sun, L.; Sun, H. Optimization of Extraction Process and Activity of Angiotensin-Converting Enzyme (ACE) Inhibitory Peptide from Walnut Meal. Foods 2024, 13, 1067. https://doi.org/10.3390/foods13071067
Meng M, She Z, Feng Y, Zhang J, Han R, Qi Y, Sun L, Sun H. Optimization of Extraction Process and Activity of Angiotensin-Converting Enzyme (ACE) Inhibitory Peptide from Walnut Meal. Foods. 2024; 13(7):1067. https://doi.org/10.3390/foods13071067
Chicago/Turabian StyleMeng, Meng, Ziyi She, Yinyin Feng, Junhan Zhang, Ran Han, Yanlong Qi, Lina Sun, and Huiqing Sun. 2024. "Optimization of Extraction Process and Activity of Angiotensin-Converting Enzyme (ACE) Inhibitory Peptide from Walnut Meal" Foods 13, no. 7: 1067. https://doi.org/10.3390/foods13071067
APA StyleMeng, M., She, Z., Feng, Y., Zhang, J., Han, R., Qi, Y., Sun, L., & Sun, H. (2024). Optimization of Extraction Process and Activity of Angiotensin-Converting Enzyme (ACE) Inhibitory Peptide from Walnut Meal. Foods, 13(7), 1067. https://doi.org/10.3390/foods13071067