
Human Norovirus Surrogate Is Highly Stable in Berry Smoothies and under In Vitro Simulated Digestion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Tulane Virus Stock Preparation and the TCID50 Infectivity Assay
2.2. TV Survival in Berry Smoothies
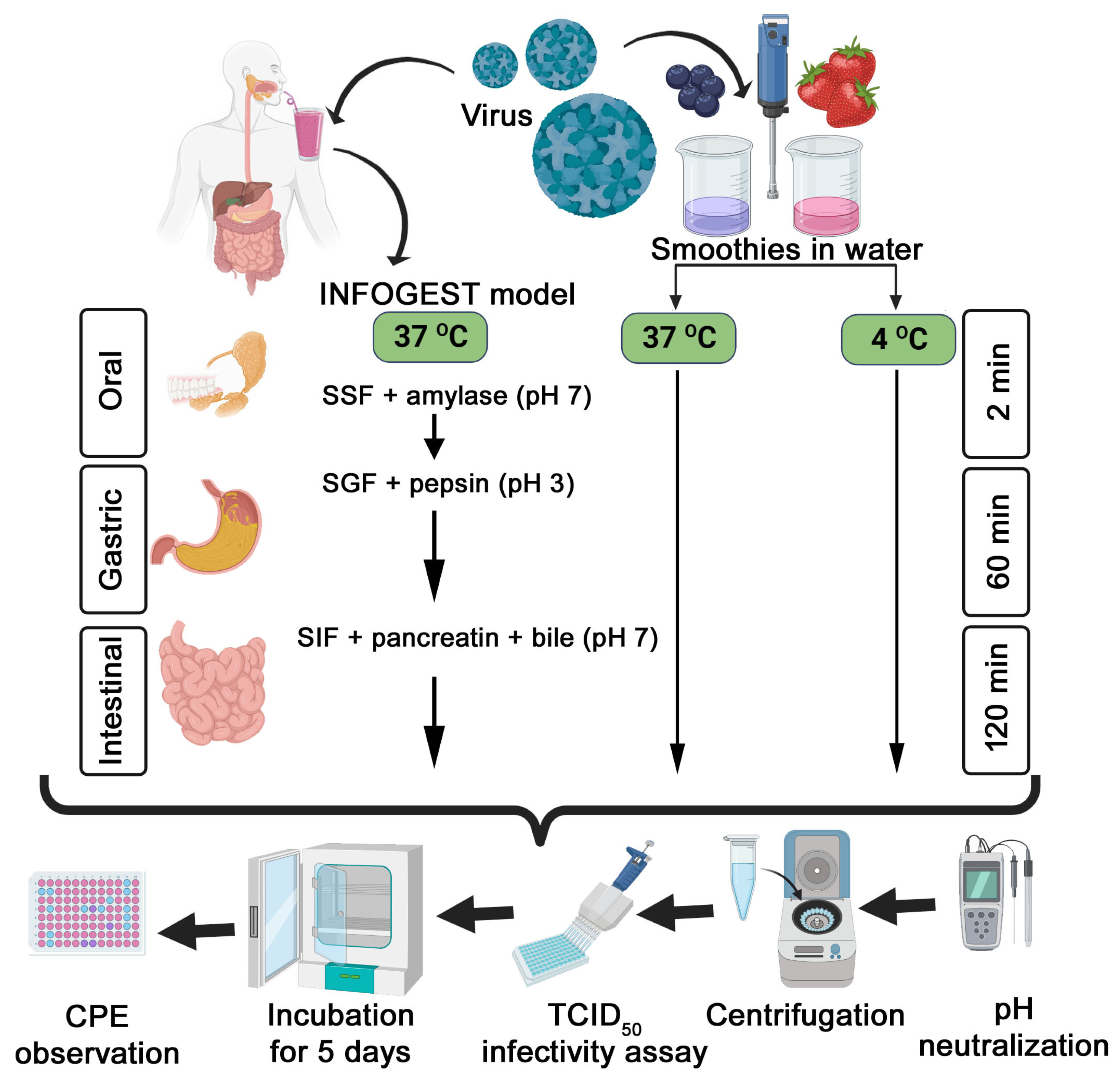
2.3. Simulated In Vitro Digestion of TV Spiked into Berry Smoothies
2.4. Statistics Analyses
3. Results
3.1. Survival of Infectious TV Spiked into Berry Smoothies
3.2. Time and Temperature Effects on TV Infectivity Log Reductions in Berry Smoothies
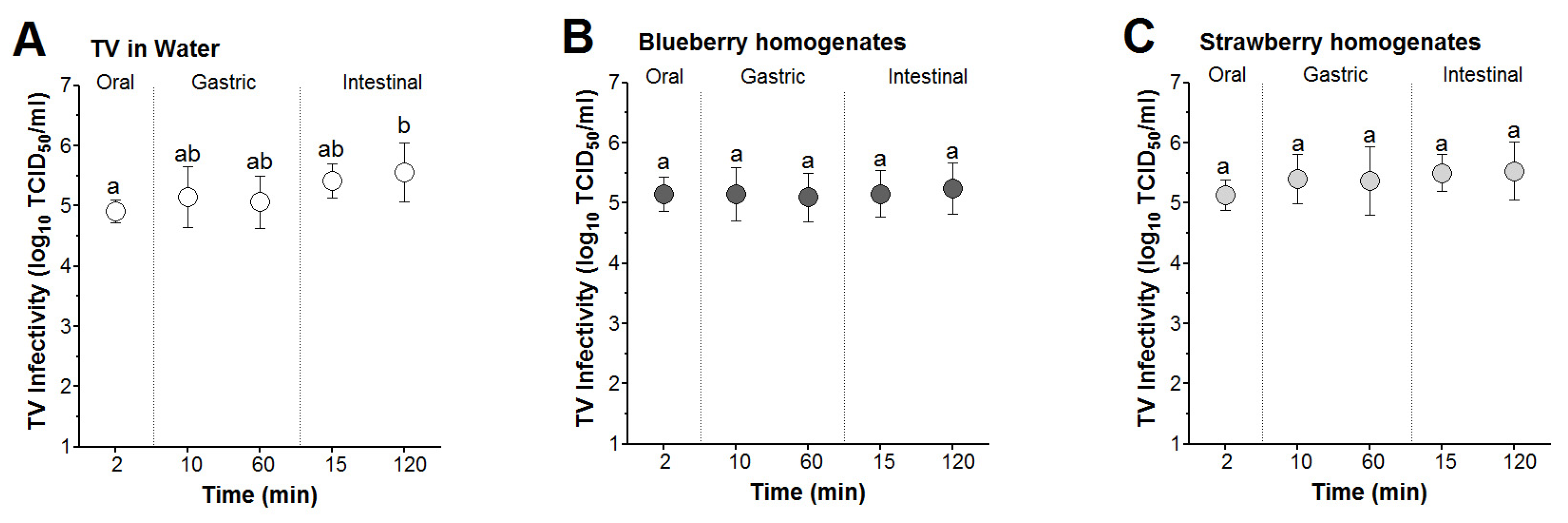
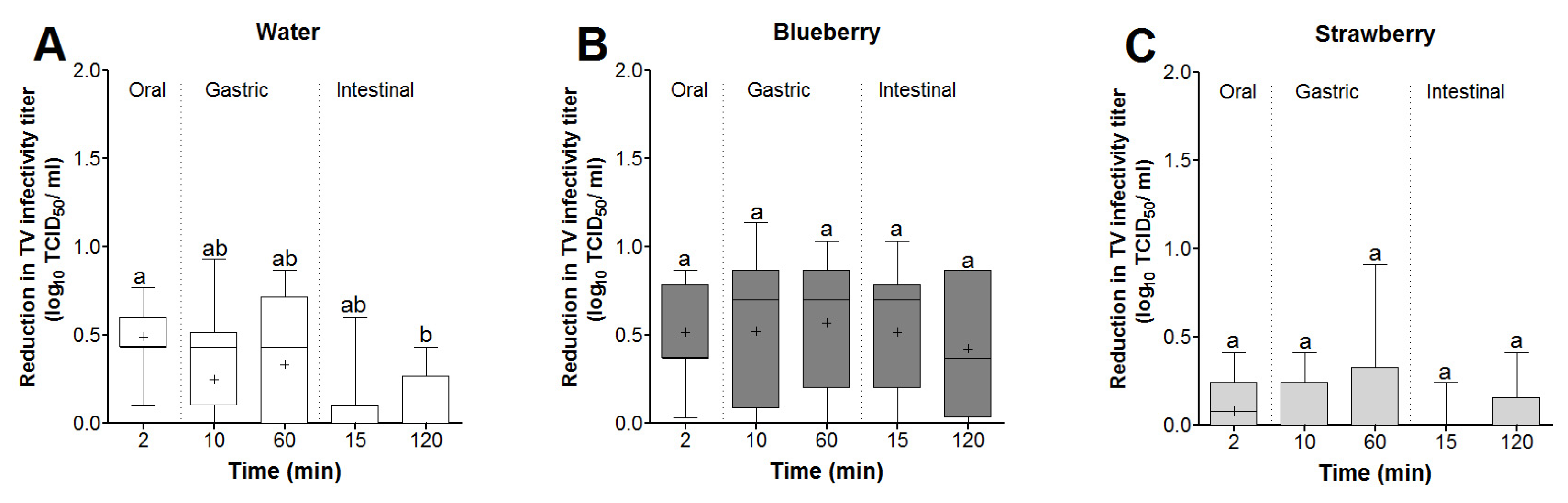
3.3. Survival of Infectious TV Spiked in Berry Smoothies under Simulated In Vitro Digestion
3.4. Time and Matrix Effects on TV Infectivity Log Reductions following In Vitro Digestion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lopman, B.A.; Steele, D.; Kirkwood, C.D.; Parashar, U.D. The Vast and Varied Global Burden of Norovirus: Prospects for Prevention and Control. PLoS Med. 2016, 13, e1001999. [Google Scholar] [CrossRef]
- Liao, Y.; Hong, X.; Wu, A.; Jiang, Y.; Liang, Y.; Gao, J.; Xue, L.; Kou, X. Global prevalence of norovirus in cases of acute gastroenteritis from 1997 to 2021: An updated systematic review and meta-analysis. Microb. Pathog. 2021, 161, 105259. [Google Scholar] [CrossRef]
- Burke, R.M.; Mattison, C.P.; Marsh, Z.; Shioda, K.; Donald, J.; Salas, S.B.; Naleway, A.L.; Biggs, C.; Schmidt, M.A.; Hall, A.J. Norovirus and Other Viral Causes of Medically Attended Acute Gastroenteritis Across the Age Spectrum: Results from the Medically Attended Acute Gastroenteritis Study in the United States. Clin. Infect. Dis. 2021, 73, e913–e920. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.J.; Wikswo, M.E.; Pringle, K.; Gould, L.H.; Parashar, U.D.; Division of Viral Diseases, N.C.f.I.; Respiratory Diseases, C.D.C. Vital signs: Foodborne norovirus outbreaks—United States, 2009–2012. MMWR. Morb. Mortal. Wkly. Rep. 2014, 63, 491–495. [Google Scholar]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Hoffmann, S.; Batz, M.B.; Morris, J.G., Jr. Annual cost of illness and quality-adjusted life year losses in the United States due to 14 foodborne pathogens. J. Food Prot. 2012, 75, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Esseili, M.A.; Gao, X.; Tegtmeier, S.; Saif, L.J.; Wang, Q. Abiotic stress and Phyllosphere Bacteria Influence the Survival of Human Norovirus and its surrogates on Preharvest Leafy Greens. Appl. Environ. Microbiol. 2015, 82, 352–363. [Google Scholar] [CrossRef]
- Esseili, M.A.; Meulia, T.; Saif, L.J.; Wang, Q. Tissue Distribution and Visualization of Internalized Human Norovirus in Leafy Greens. Appl. Environ. Microbiol. 2018, 84, e00292-18. [Google Scholar] [CrossRef]
- Esseili, M.A.; Saif, L.J.; Farkas, T.; Wang, Q. Feline Calicivirus, Murine Norovirus, Porcine Sapovirus, and Tulane Virus Survival on Postharvest Lettuce. Appl. Environ. Microbiol. 2015, 81, 5085–5092. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Selma, M.V.; Suslow, T.; Jacxsens, L.; Uyttendaele, M.; Allende, A. Pre- and postharvest preventive measures and intervention strategies to control microbial food safety hazards of fresh leafy vegetables. Crit. Rev. Food Sci. Nutr. 2015, 55, 453–468. [Google Scholar] [CrossRef]
- Taban, B.M.; Halkman, A.K. Do leafy green vegetables and their ready-to-eat [RTE] salads carry a risk of foodborne pathogens? Anaerobe 2011, 17, 286–287. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.; Phan-Thien, K.Y.; van Ogtrop, F.; Bell, T.; McConchie, R. Outbreaks, occurrence, and control of norovirus and hepatitis a virus contamination in berries: A review. Crit. Rev. Food Sci. Nutr. 2021, 61, 116–138. [Google Scholar] [CrossRef] [PubMed]
- Baert, L.; Mattison, K.; Loisy-Hamon, F.; Harlow, J.; Martyres, A.; Lebeau, B.; Stals, A.; Van Coillie, E.; Herman, L.; Uyttendaele, M. Review: Norovirus prevalence in Belgian, Canadian and French fresh produce: A threat to human health? Int. J. Food Microbiol. 2011, 151, 261–269. [Google Scholar] [CrossRef]
- Macori, G.; Gilardi, G.; Bellio, A.; Bianchi, D.M.; Gallina, S.; Vitale, N.; Gullino, M.L.; Decastelli, L. Microbiological Parameters in the Primary Production of Berries: A Pilot Study. Foods 2018, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, Z.; Wang, Y.; Liu, Z.; Guan, X.; Ma, Y.; Zhou, H.; Jiang, Y.; Cui, W.; Wang, L.; et al. Surveillance of norovirus contamination in commercial fresh/frozen berries from Heilongjiang Province, China, using a TaqMan real-time RT-PCR assay. Food Microbiol. 2019, 82, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, D.; Gagne, M.J.; Poitras, E.; Brassard, J. Persistence of murine norovirus, bovine rotavirus, and hepatitis A virus on stainless steel surfaces, in spring water, and on blueberries. Food Microbiol. 2019, 84, 103257. [Google Scholar] [CrossRef]
- Zhao, Z.; Hossain, M.I.; Jung, S.; Wang, Z.; Yeo, D.; Song, M.; Min, A.; Park, S.; Choi, C. Survival of murine norovirus and hepatitis A virus in bottled drinking water, strawberries, and oysters. Food Control 2022, 133, 108623. [Google Scholar] [CrossRef]
- Butot, S.; Putallaz, T.; Sanchez, G. Effects of sanitation, freezing and frozen storage on enteric viruses in berries and herbs. Int. J. Food Microbiol. 2008, 126, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Verhaelen, K.; Bouwknegt, M.; Lodder-Verschoor, F.; Rutjes, S.A.; de Roda Husman, A.M. Persistence of human norovirus GII.4 and GI.4, murine norovirus, and human adenovirus on soft berries as compared with PBS at commonly applied storage conditions. Int. J. Food Microbiol. 2012, 160, 137–144. [Google Scholar] [CrossRef]
- Bernier, C.; Goetz, C.; Jubinville, E.; Jean, J. The New Face of Berries: A Review of Their Antiviral Proprieties. Foods 2021, 11, 102. [Google Scholar] [CrossRef]
- Joshi, S.S.; Howell, A.B.; D’Souza, D.H. Reduction of Enteric Viruses by Blueberry Juice and Blueberry Proanthocyanidins. Food Environ. Virol. 2016, 8, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi Gorji, M.; Tan, M.T.H.; Zhao, M.Y.; Li, D. No Clinical Symptom Experienced after Consumption of Berry Fruits with Positive RT-qPCR Signals of Human Norovirus. Pathogens 2021, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Esseili, M.A.; Mann, A.; Narwankar, R.; Kassem, I.I.; Diez-Gonzalez, F.; Hogan, R.J. SARS-CoV-2 remains infectious for at least a month on artificially-contaminated frozen berries. Food Microbiol. 2022, 107, 104084. [Google Scholar] [CrossRef] [PubMed]
- Esseili, M.A. In vitro digestion of SARS-CoV-2 contaminated berries reveals high inactivation of infectious virus during gastrointestinal passage. Appl. Environ. Microbiol. 2023, 89, e0133923. [Google Scholar] [CrossRef] [PubMed]
- Esseili, M.A. Great escape: How infectious SARS-CoV-2 avoids inactivation by gastric acidity and intestinal bile. Gut 2023, 72, 808–810. [Google Scholar] [CrossRef]
- Joshi, S.; Howell, A.B.; D’Souza, D.H. Blueberry proanthocyanidins against human norovirus surrogates in model foods and under simulated gastric conditions. Food Microbiol. 2017, 63, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.S.; Su, X.; D’Souza, D.H. Antiviral effects of grape seed extract against feline calicivirus, murine norovirus, and hepatitis A virus in model food systems and under gastric conditions. Food Microbiol. 2015, 52, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Falco, I.; Randazzo, W.; Rodriguez-Diaz, J.; Gozalbo-Rovira, R.; Luque, D.; Aznar, R.; Sanchez, G. Antiviral activity of aged green tea extract in model food systems and under gastric conditions. Int. J. Food Microbiol. 2019, 292, 101–106. [Google Scholar] [CrossRef]
- Zivkovic, I.; Savikin, K.; Zivkovic, J.; Zdunic, G.; Jankovic, T.; Lazic, D.; Radin, D. Antiviral Effects of Pomegranate Peel Extracts on Human Norovirus in Food Models and Simulated Gastrointestinal Fluids. Plant Foods Hum. Nutr. 2021, 76, 203–209. [Google Scholar] [CrossRef]
- Lee, E.J.; Han, S.; Hyun, S.W.; Song, G.B.; Ha, S.D. Survival of human coronavirus 229E at different temperatures on various food-contact surfaces and food and under simulated digestive conditions. Food Res. Int. 2022, 162, 112014. [Google Scholar] [CrossRef] [PubMed]
- Zang, R.; Gomez Castro, M.F.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Liu, Z.; Brulois, K.F.; Wang, X.; Greenberg, H.B.; et al. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Sci. Immunol. 2020, 5, eabc3582. [Google Scholar] [CrossRef] [PubMed]
- Dubois, E.; Agier, C.; Traore, O.; Hennechart, C.; Merle, G.; Cruciere, C.; Laveran, H. Modified concentration method for the detection of enteric viruses on fruits and vegetables by reverse transcriptase-polymerase chain reaction or cell culture. J. Food Prot. 2002, 65, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- de Souza, V.R.; Pereira, P.A.; da Silva, T.L.; de Oliveira Lima, L.C.; Pio, R.; Queiroz, F. Determination of the bioactive compounds, antioxidant activity and chemical composition of Brazilian blackberry, red raspberry, strawberry, blueberry and sweet cherry fruits. Food Chem. 2014, 156, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.; Feucht, W.; Schmid, M. Bioactive Compounds of Strawberry and Blueberry and Their Potential Health Effects Based on Human Intervention Studies: A Brief Overview. Nutrients 2019, 11, 1510. [Google Scholar] [CrossRef] [PubMed]
- Arthur, S.E.; Gibson, K.E. Physicochemical stability profile of Tulane virus: A human norovirus surrogate. J. Appl. Microbiol. 2015, 119, 868–875. [Google Scholar] [CrossRef]
- Tian, P.; Yang, D.; Quigley, C.; Chou, M.; Jiang, X. Inactivation of the Tulane virus, a novel surrogate for the human norovirus. J. Food Prot. 2013, 76, 712–718. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hooda, R.; Esseili, M.A. Human Norovirus Surrogate Is Highly Stable in Berry Smoothies and under In Vitro Simulated Digestion. Foods 2024, 13, 1066. https://doi.org/10.3390/foods13071066
Hooda R, Esseili MA. Human Norovirus Surrogate Is Highly Stable in Berry Smoothies and under In Vitro Simulated Digestion. Foods. 2024; 13(7):1066. https://doi.org/10.3390/foods13071066
Chicago/Turabian StyleHooda, Riya, and Malak A. Esseili. 2024. "Human Norovirus Surrogate Is Highly Stable in Berry Smoothies and under In Vitro Simulated Digestion" Foods 13, no. 7: 1066. https://doi.org/10.3390/foods13071066
APA StyleHooda, R., & Esseili, M. A. (2024). Human Norovirus Surrogate Is Highly Stable in Berry Smoothies and under In Vitro Simulated Digestion. Foods, 13(7), 1066. https://doi.org/10.3390/foods13071066