

Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Edible Grass Fermentation
2.3. Cell Viability Determination
2.4. Establishment of Acetaminophen-Induced HepG2 Cells Injury Model
2.5. Determination of Biochemical Indicators of Acetaminophen-Induced HepG2 Cells
2.6. Sample Preparation
2.7. Untargeted Metabolomics Analysis
2.8. Data Processing and Statistical Analysis
3. Results and Discussion
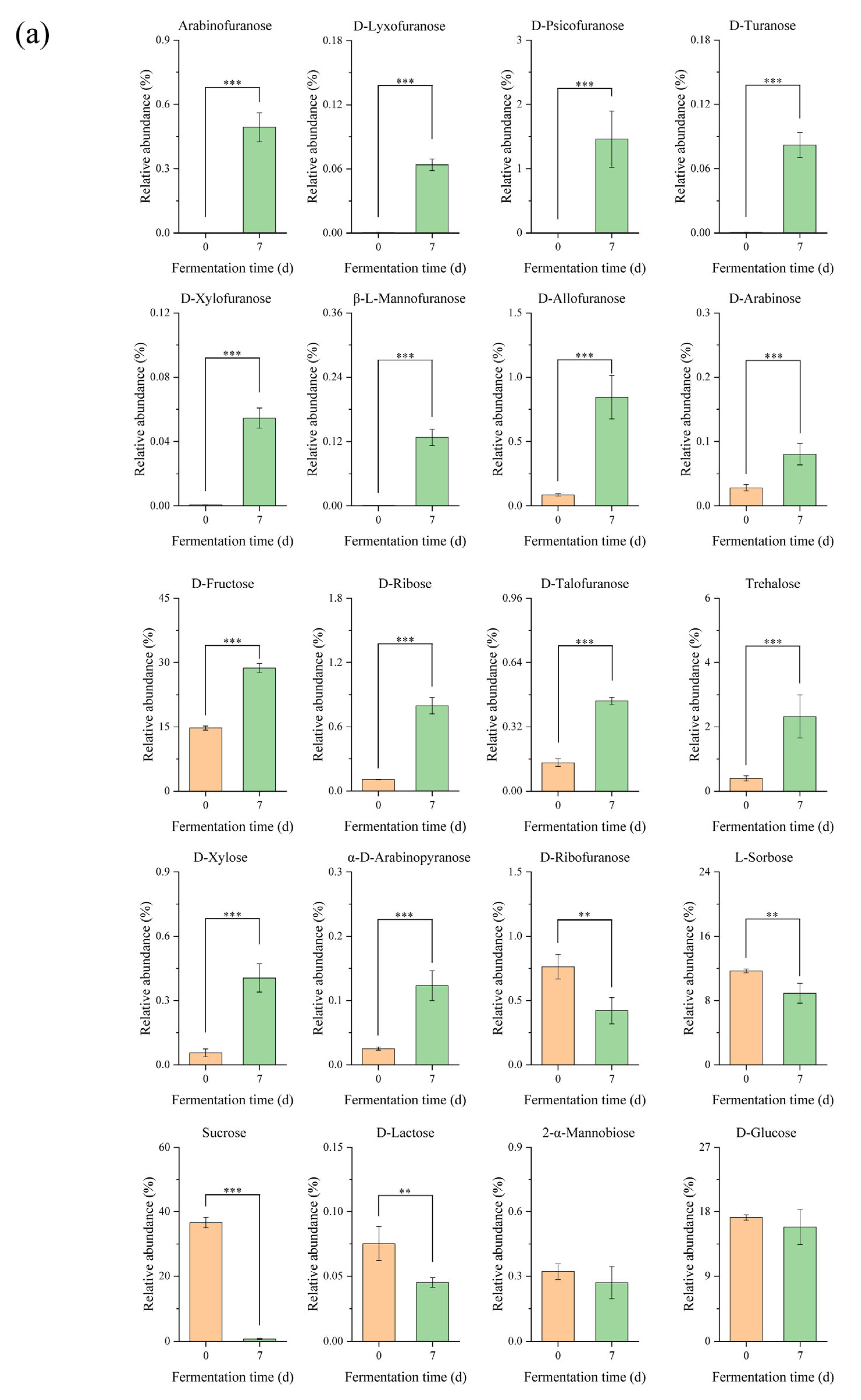
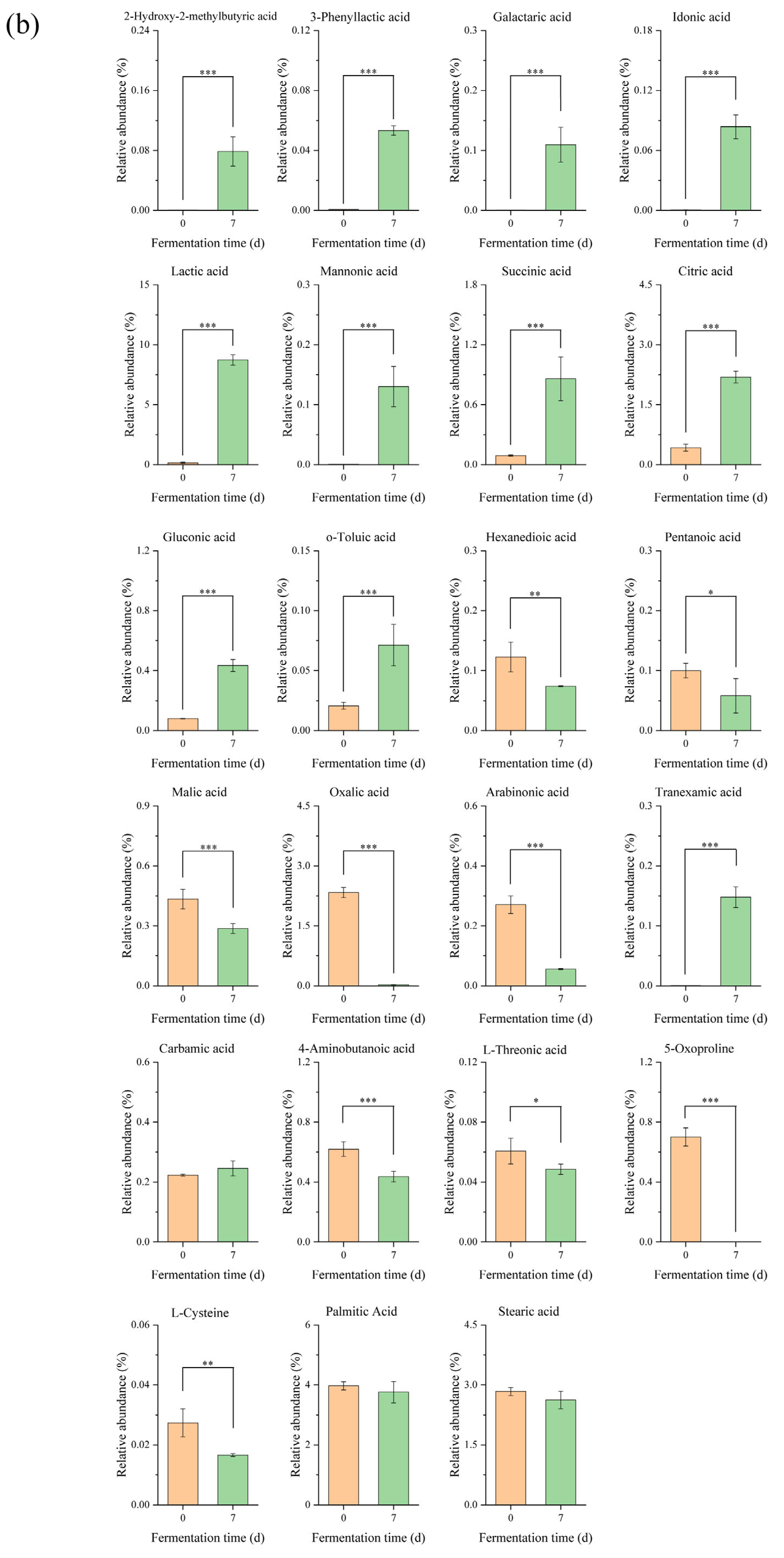
3.1. Metabolomics Analysis in Edible Grass Fermentation
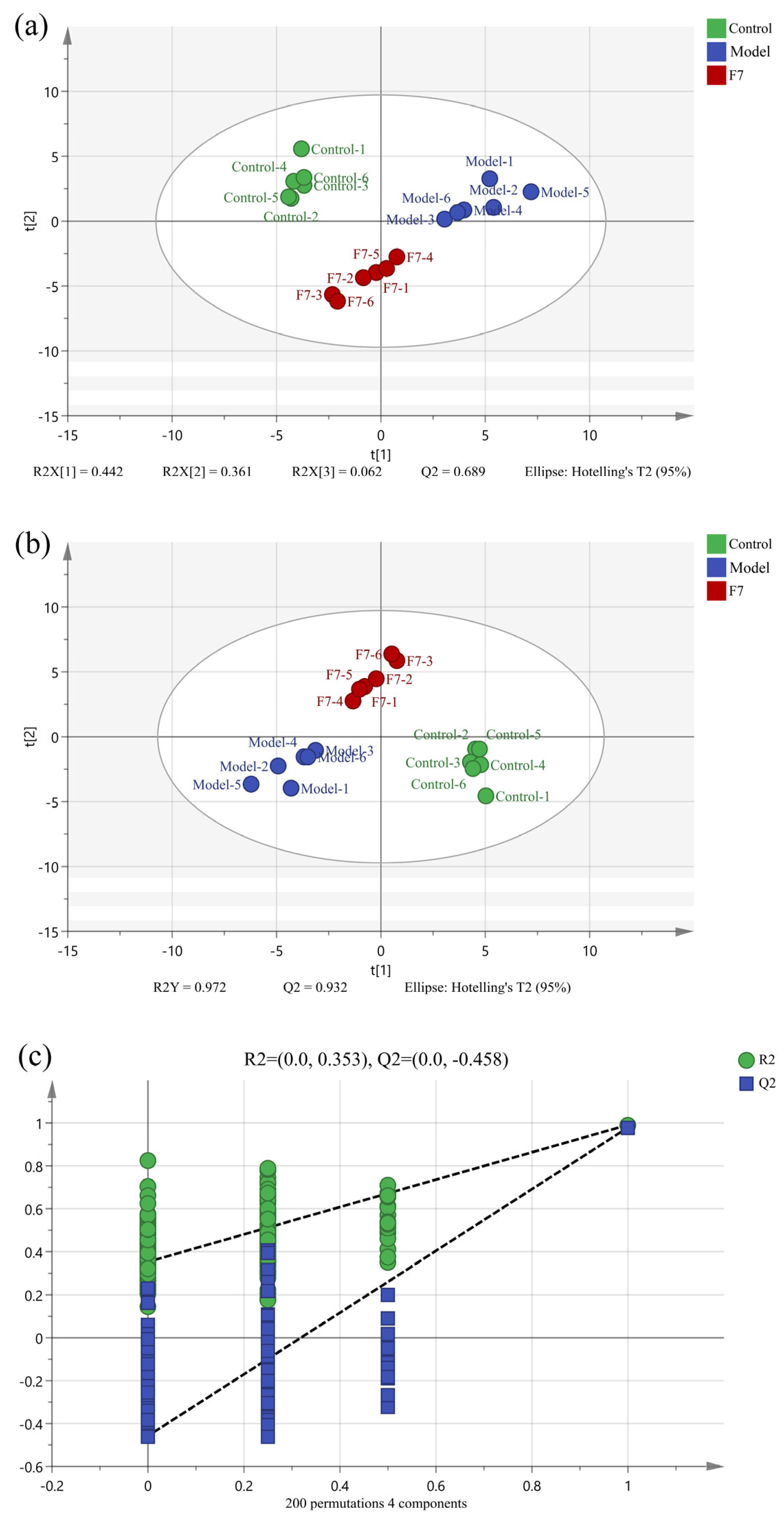
3.1.1. Multivariate Statistical Analysis
3.1.2. Differential Metabolites Analysis
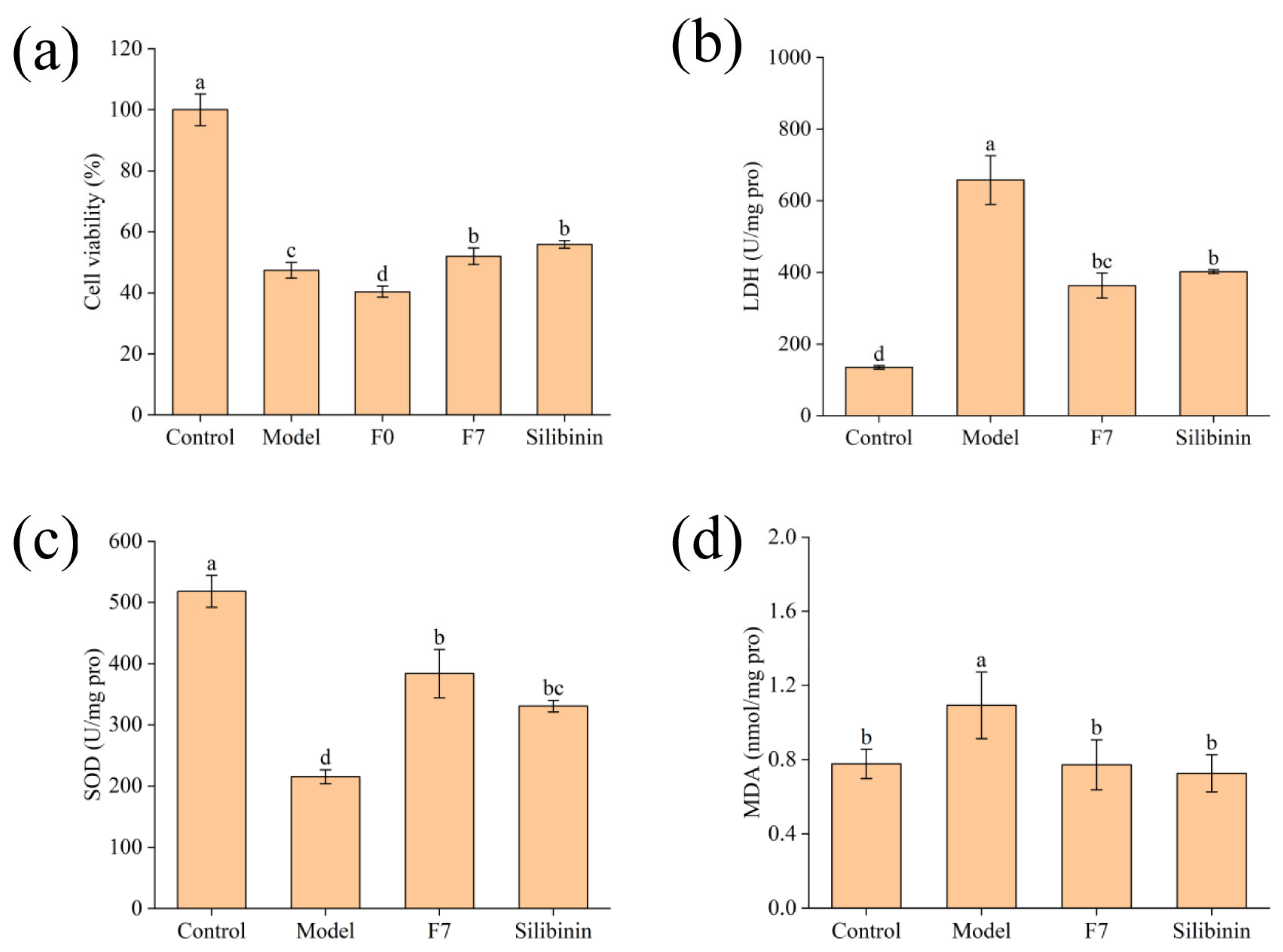
3.2. Protective Effect of Fermented Edible Grass on Acetaminophen-Induced Liver Injury at the Cellular Level
3.2.1. Effect of Edible Grass, Fermented Edible Grass and Acetaminophen on Cell Viability
3.2.2. Protective Effect of Fermented Edible Grass on Acetaminophen-Induced HepG2 Cells
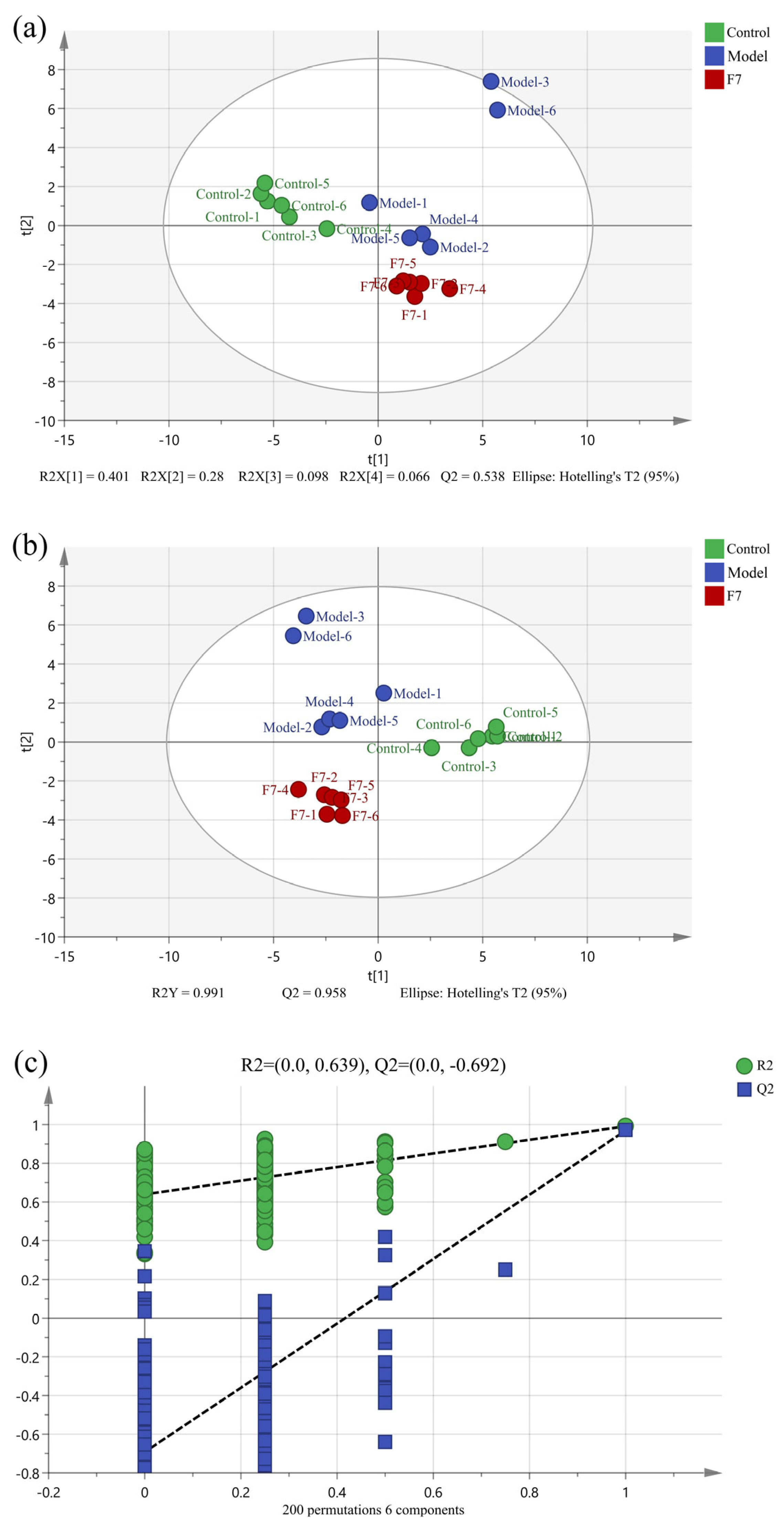
3.3. Analysis of Intracellular and Extracellular Metabolomics in HepG2 Cells
3.3.1. Multivariate Statistical Analysis
3.3.2. Identification of Differential Metabolites
3.3.3. Analysis of Metabolic Pathway
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; He, T.; Mao, J.; Sha, R. Effects of Lactic Acid Bacteria Fermentation on Physicochemical Properties, Functional Compounds and Antioxidant Activity of Edible Grass. Fermentation 2022, 8, 647. [Google Scholar] [CrossRef]
- Li, X.; He, T.; Yang, F.; Wang, C.; Zhou, Y.; Sha, R.; Mao, J. Analysis of nutritional components, functional components and bioactivity of edible grass. Sci. Technol. Food Ind. 2023, 44, 307–315. [Google Scholar]
- Li, X.; He, T.; Mao, Y.; Mao, J.; Lai, X.; Tu, H.; Zhou, Y.; Sha, R. Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation. Fermentation 2023, 9, 377. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H.; Lin, M. Research Progress of Fermented Functional Foods and Protein Factory-Microbial Fermentation Technology. Fermentation 2022, 8, 688. [Google Scholar] [CrossRef]
- Adebo, O.A.; Oyeyinka, S.A.; Adebiyi, J.A.; Feng, X.; Wilkin, J.D.; Kewuyemi, Y.O.; Abrahams, A.M.; Tugizimana, F. Application of gas chromatography-mass spectrometry (GC-MS)-based metabolomics for the study of fermented cereal and legume foods: A review. Int. J. Food Sci. Technol. 2021, 56, 1514–1534. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, L.; Li, L.; Li, Q.; Gao, M.; Yang, X.; Yang, J.; Xie, C. Dynamic changes of differential metabolites and key metabolic pathways of Gastrodia elata Blume during fermentation. Int. J. Food Prop. 2022, 25, 1539–1548. [Google Scholar] [CrossRef]
- Seo, S.-H.; Park, S.-E.; Kim, E.-J.; Lee, K.-I.; Na, C.-S.; Son, H.-S. A GC-MS based metabolomics approach to determine the effect of salinity on Kimchi. Food Res. Int. 2018, 105, 492–498. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, C.; Yang, Q.; Guo, Z.; Yang, B.; Lu, W.; Li, D.; Tian, F.; Liu, X.; Zhang, H.; et al. Selection of Taste Markers Related to Lactic Acid Bacteria Microflora Metabolism for Chinese Traditional Paocai: A Gas Chromatography-Mass Spectrometry-Based Metabolomics Approach. J. Agric. Food Chem. 2016, 64, 2415–2422. [Google Scholar] [CrossRef]
- Peng, Q.; Meng, K.; Zheng, H.; Yu, H.; Zhang, Y.; Yang, X.; Lin, Z.; Xie, G. Metabolites comparison in post-fermentation stage of manual (mechanized) Chinese Huangjiu (yellow rice wine) based on GC-MS metabolomics. Food Chem. X 2022, 14, 100324. [Google Scholar] [CrossRef]
- Mei, S.; Song, X.; Wang, Y.; Wang, J.; Su, S.; Zhu, J.; Geng, Y. Studies on Protection of Astaxanthin from Oxidative Damage Induced by H2O2 in RAW 264.7 Cells Based on 1H NMR Metabolomics. J. Agric. Food Chem. 2019, 67, 13568–13576. [Google Scholar] [CrossRef]
- Dai, J.; Hu, Y.; Si, Q.; Gu, Y.; Xiao, Z.; Ge, Q.; Sha, R. Antioxidant and Hypoglycemic Activity of Sequentially Extracted Fractions from Pingguoli Pear Fermentation Broth and Identification of Bioactive Compounds. Molecules 2022, 27, 6077. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Q.; Weng, L.; Zou, L.; Jiang, H.; Qiu, J.; Fu, J. Analysis of sucrose addition on the physicochemical properties of blueberry wine in the main fermentation. Front. Nutr. 2023, 9, 1092696. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Deng, L.; Yang, J. Review on Synthesis and Occurrence of Oligoarabino-, Oligogalacto- and Oligofructo-furanoses. Chinese J. Org. Chem. 2013, 33, 245–258. [Google Scholar] [CrossRef]
- Denman, L.J.; Morris, G.A. An experimental design approach to the chemical characterisation of pectin polysaccharides extracted from Cucumis melo Inodorus. Carbohyd. Polym. 2015, 117, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Mamat, S.F.; Azizan, K.A.; Baharum, S.N.; Noor, N.M.; Aizat, W.M. GC-MS and LC-MS analyses reveal the distribution of primary and secondary metabolites in mangosteen (Garcinia mangostana Linn.) fruit during ripening. Sci. Hortic. 2020, 262, 109004. [Google Scholar] [CrossRef]
- Kim, J.-G.; Yoo, H.; Sang, H. Compositional changes in maesil-cheong formulated with turanose during the storage period. Korean J. Food Sci. Technol. 2021, 53, 688–694. [Google Scholar]
- Tian, Y.; Deng, Y.; Zhang, W.; Mu, W. Sucrose isomers as alternative sweeteners: Properties, production, and applications. Appl. Microbiol. Biot. 2019, 103, 8677–8687. [Google Scholar] [CrossRef]
- Teitelbaum, J.E.; Johnson, C.; Cyr, J.S. The use of D-ribose in chronic fatigue syndrome and fibromyalgia: A pilot study. J. Altern. Complem. Med. 2006, 12, 857–862. [Google Scholar] [CrossRef]
- Park, H.-C.; Kim, Y.-J.; Lee, C.-W.; Rho, Y.-T.; Kang, J.; Lee, D.-H.; Seong, Y.-J.; Park, Y.-C.; Lee, D.; Kim, S.-G. Production of D-ribose by metabolically engineered Escherichia coli. Process Biochem. 2017, 52, 73–77. [Google Scholar] [CrossRef]
- Chang, S.-W.; Chang, W.-H.; Lee, M.-R.; Yang, T.-J.; Yu, N.-Y.; Chen, C.-S.; Shaw, J.-F. Simultaneous Production of Trehalose, Bioethanol, and High-Protein Product from Rice by an Enzymatic Process. J. Agric. Food Chem. 2010, 58, 2908–2914. [Google Scholar] [CrossRef]
- Evans, T.D.; Jeong, S.-J.; Zhang, X.; Sergin, I.; Razani, B. TFEB and trehalose drive the macrophage autophagy-lysosome system to protect against atherosclerosis. Autophagy 2018, 14, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Yee, D.; Nagarajan, N.; Bianchi, F.; Saito, T.; Valenti, V.; Tong, M.; Del Re, D.P.; Vecchione, C.; Schirone, L.; et al. Trehalose-Induced Activation of Autophagy Improves Cardiac Remodeling After Myocardial Infarction. J. Am. Coll. Cardiol. 2018, 71, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Cheudjeu, A. The SARS-CoV-2 Entry Inhibition Mechanisms of Serine Protease Inhibitors, OM-85, Heparin and Soluble HS Might Be Linked to HS Attachment Sites. Molecules 2022, 27, 1947. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhu, W.; Han, J.; Li, Z.; Ge, H.; Lin, D. Analysis of organic acids in Chinese raisin tree (Hovenia dulcis) peduncle and their changes in liquid fermentation process. Food Sci. Biotechnol. 2012, 21, 1119–1127. [Google Scholar] [CrossRef]
- Mu, W.; Yu, S.; Zhu, L.; Zhang, T.; Jiang, B. Recent research on 3-phenyllactic acid, a broad-spectrum antimicrobial compound. Appl. Microbiol. Biot. 2012, 95, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, C.; Zhu, Y.; Zhou, C.; Xiong, Z.; Eweys, A.S.; Zhou, H.; Dong, Y.; Xiao, X. Metabolomics strategy for revealing the components in fermented barley extracts with Lactobacillus plantarum dy-1. Food Res. Int. 2021, 139, 109808. [Google Scholar] [CrossRef] [PubMed]
- Gaenzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Chidi, B.S.; Bauer, F.F.; Rossouw, D. Organic Acid Metabolism and the Impact of Fermentation Practices on Wine Acidity—A Review. S. Afr. J. Enol. Vitic. 2018, 39, 315–329. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Brar, S.K.; Verma, M.; Tyagi, R.D. Recent Advances in Citric Acid Bio-production and Recovery. Food Bioprocess Technol. 2011, 4, 505–529. [Google Scholar] [CrossRef]
- Ramachandran, S.; Fontanille, P.; Pandey, A.; Larroche, C. Gluconic acid: Properties, applications and microbial production. Food Technol. Biotech. 2006, 44, 185–195. [Google Scholar]
- Monedero, V.; Perez-Martinez, G.; Yebra, M.J. Perspectives of engineering lactic acid bacteria for biotechnological polyol production. Appl. Microbiol. Biot. 2010, 86, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-E.; Yoo, S.-A.; Seo, S.-H.; Lee, K.-I.; Na, C.-S.; Son, H.-S. GC-MS based metabolomics approach of Kimchi for the understanding of Lactobacillus plantarum fermentation characteristics. LWT 2016, 68, 313–321. [Google Scholar] [CrossRef]
- Ji, X.-J.; Huang, H.; Ouyang, P.-K. Microbial 2,3-butanediol production: A state-of-the-art review. Biotechnol. Adv. 2011, 29, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Grobben, G.J.; Peters, S.W.P.G.; Wisselink, H.W.; Weusthuis, R.A.; Eggink, G. Spontaneous Formation of a Mannitol-Producing Variant of Leuconostoc pseudomesenteroides Grown in the Presence of Fructose. Appl. Environ. Microb. 2001, 67, 2867–2870. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-J.; Chiu, C.-H.; Tseng, J.-K.; Yang, K.-T.; Chen, Y.-C. Ameliorative effects of D-glucuronolactone on oxidative stress and inflammatory/fibrogenic responses in livers of thioacetamide-treated rats. J. Funct. Foods 2015, 14, 154–162. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, J.; Tian, C.; Ren, C.; Chen, P.; Men, Y.; Sun, Y. High-Yield Biosynthesis of Glucosylglycerol through Coupling Phosphorolysis and Transglycosylation Reactions. J. Agric. Food Chem. 2020, 68, 15249–15256. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yao, Y.; Wei, Z.-y.; Wang, S.-x.; Wu, Y.-c.; Hu, Y.; Yang, C.-c.; Min, J.-l.; Li, L.-y.; Zhou, H.; et al. Deletion of p38γ attenuates ethanol consumption- and acetaminophen-induced liver injury in mice through promoting Dlg1. Acta Pharmacol. Sin. 2022, 43, 1733–1748. [Google Scholar] [CrossRef] [PubMed]
- Salamone, F.; Galvano, F.; Marino, A.; Paternostro, C.; Tibullo, D.; Bucchieri, F.; Mangiameli, A.; Parola, M.; Bugianesi, E.; Volti, G.L. Silibinin improves hepatic and myocardial injury in mice with nonalcoholic steatohepatitis. Digest. Liver Dis. 2012, 44, 334–342. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, T.; Gan, Z.; Li, H.; Li, Y.; Zhang, Y.; Zhao, X. Metabolomic analysis of untargeted bovine uterine secretions in dairy cows with endometritis using ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry. Res. Vet. Sci. 2021, 139, 51–58. [Google Scholar] [CrossRef]
- Chen, T.; Ni, Y.; Ma, X.; Bao, Y.; Liu, J.; Huang, F.; Hu, C.; Xie, G.; Zhao, A.; Jia, W.; et al. Branched-chain and aromatic amino acid profiles and diabetes risk in Chinese populations. Sci. Rep. 2016, 6, 20594. [Google Scholar] [CrossRef]
- Tajiri, K.; Shimizu, Y. Branched-chain amino acids in liver diseases. World J. Gastroenterol. 2013, 19, 7620–7629. [Google Scholar] [CrossRef]
- Brezmes, J.; Llambrich, M.; Cumeras, R.; Guma, J. Urine NMR Metabolomics for Precision Oncology in Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 11171. [Google Scholar] [CrossRef]
- Jiang, J.; Srivastava, S.; Seim, G.; Pavlova, N.N.; King, B.; Zou, L.; Zhang, C.; Zhong, M.; Feng, H.; Kapur, R.; et al. Promoter demethylation of the asparagine synthetase gene is required for ATF4-dependent adaptation to asparagine depletion. J. Biol. Chem. 2019, 294, 18674–18684. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gao, P.; Cheng, F.; Wang, X.; Duan, Y. Measurement of salivary metabolite biomarkers for early monitoring of oral cancer with ultra performance liquid chromatography-mass spectrometry. Talanta 2014, 119, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Cui, C.; Wang, C.; Lu, S.; Zhang, M.; Chen, D.; Jiang, P. Systematic Evaluations of Doxorubicin-Induced Toxicity in Rats Based on Metabolomics. Acs Omega 2021, 6, 358–366. [Google Scholar] [CrossRef]
- Vyshtakalyuk, A.B.; Parfenov, A.A.; Gumarova, L.F.; Khasanshina, L.R.; Belyaev, G.P.; Nazarov, N.G.; Kondrashina, D.A.; Galyametdinova, I.V.; Semenov, V.E.; Zobov, V.V. Conjugate of pyrimidine derivative, the drug xymedon with succinic acid protects liver cells. J. Biochem. Mol. Toxic. 2021, 35, 22660. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Liu, X.; Zhao, S.; Li, Z.; Qin, X. 1H NMR-Based Fecal Metabolomics Reveals Changes in Gastrointestinal Function of Aging Rats Induced byd-Galactose. Rejuv. Res. 2021, 24, 86–96. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R.T. | Metabolite | VIP | Trend | |
|---|---|---|---|---|
| Mod vs. Con | F7 vs. Mod | |||
| 11.976 | L-Valine | 1.35 | NC | ↑ |
| 13.567 | Glycine | 1.02 | ↓ | NC |
| 14.551 | Serine | 1.12 | ↓ | NC |
| 17.177 | Succinic acid | 1.38 | NC | ↑ |
| 18.533 | L-Glutamic acid | 1.01 | ↓ | NC |
| 22.499 | D-Mannose | 1.18 | ↓ | ↑ |
| 22.779 | L-Phenylalanine | 1.25 | ↑ | ↓ |
| 23.489 | Gluconic acid | 1.38 | NC | ↓ |
| 29.354 | Eicosyl isopropyl ether | 1.08 | NC | ↓ |
| 30.618 | Sucrose | 1.15 | ↓ | NC |
| 34.715 | Cholesterol | 1.03 | ↓ | NC |
| 34.944 | 3-Cyclohexene-1-ethanol | 1.14 | ↑ | NC |
| 36.729 | Propyleneglycol monoleate | 1.33 | ↑ | ↓ |
| R.T. | Metabolite | VIP | Trend | |
|---|---|---|---|---|
| Mod vs. Con | F7 vs. Mod | |||
| 9.069 | Propanoic acid | 1.04 | ↑ | ↓ |
| 9.865 | Alanine | 1.04 | NC | ↓ |
| 11.976 | L-Valine | 1.01 | ↑ | ↓ |
| 12.371 | Urea | 1.12 | ↓ | ↓ |
| 12.977 | L-Leucine | 1.02 | ↑ | ↓ |
| 13.378 | L-Proline | 1.10 | ↓ | ↓ |
| 13.567 | Glycine | 1.09 | ↓ | NC |
| 14.551 | Serine | 1.11 | ↓ | ↓ |
| 14.992 | L-Threonine | 1.12 | ↑ | ↓ |
| 16.062 | n-Butylamine | 1.03 | ↓ | NC |
| 16.994 | Butane | 1.10 | ↓ | ↑ |
| 18.528 | Glutamine | 1.09 | ↑ | ↓ |
| 18.596 | L-Phenylalanine | 1.07 | ↑ | ↓ |
| 19.283 | L-Asparagine | 1.04 | ↑ | ↓ |
| 21.263 | L-Ornithine | 1.03 | ↓ | ↓ |
| 21.372 | 1,2,3-Propanetricarboxylic acid | 1.05 | ↓ | NC |
| 22.007 | 3-Methyl-3-pentanol | 1.01 | ↑ | ↓ |
| 22.722 | D-Galactose | 1.14 | ↑ | ↓ |
| 22.991 | D-Mannitol | 1.10 | NC | ↓ |
| 24.770 | Myo-inositol | 1.11 | ↓ | ↓ |
| No. | Metabolic Pathway Name | Impact a | Hits b |
|---|---|---|---|
| 1 | D-Glutamine and D-glutamate metabolism | 0.500 | 1 |
| 2 | Phenylalanine, tyrosine and tryptophan biosynthesis | 0.500 | 1 |
| 3 | Glycine, serine and threonine metabolism | 0.463 | 2 |
| 4 | Phenylalanine metabolism | 0.357 | 1 |
| 5 | Alanine, aspartate and glutamate metabolism | 0.197 | 2 |
| 6 | Aminoacyl-tRNA biosynthesis | 0.167 | 5 |
| 7 | Glyoxylate and dicarboxylate metabolism | 0.148 | 3 |
| 8 | Arginine biosynthesis | 0.117 | 1 |
| 9 | Glutathione metabolism | 0.108 | 2 |
| No. | Metabolic Pathway Name | Impact a | Hits b |
|---|---|---|---|
| 1 | Phenylalanine, tyrosine and tryptophan biosynthesis | 0.500 | 1 |
| 2 | Glycine, serine and threonine metabolism | 0.463 | 3 |
| 3 | Phenylalanine metabolism | 0.357 | 1 |
| 4 | Galactose metabolism | 0.356 | 2 |
| 5 | Arginine and proline metabolism | 0.188 | 2 |
| 6 | Aminoacyl-tRNA biosynthesis | 0.167 | 10 |
| 7 | Glyoxylate and dicarboxylate metabolism | 0.148 | 3 |
| 8 | Inositol phosphate metabolism | 0.129 | 1 |
| 9 | Alanine, aspartate and glutamate metabolism | 0.114 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, T.; Li, X.; Wang, Z.; Mao, J.; Mao, Y.; Sha, R. Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells. Foods 2024, 13, 470. https://doi.org/10.3390/foods13030470
He T, Li X, Wang Z, Mao J, Mao Y, Sha R. Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells. Foods. 2024; 13(3):470. https://doi.org/10.3390/foods13030470
Chicago/Turabian StyleHe, Tao, Xianxiu Li, Zhenzhen Wang, Jianwei Mao, Yangchen Mao, and Ruyi Sha. 2024. "Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells" Foods 13, no. 3: 470. https://doi.org/10.3390/foods13030470
APA StyleHe, T., Li, X., Wang, Z., Mao, J., Mao, Y., & Sha, R. (2024). Studies on the Changes of Fermentation Metabolites and the Protective Effect of Fermented Edible Grass on Stress Injury Induced by Acetaminophen in HepG2 Cells. Foods, 13(3), 470. https://doi.org/10.3390/foods13030470