
Microbial Detoxification of Residual Pesticides in Fermented Foods: Current Status and Prospects



Abstract
1. Introduction
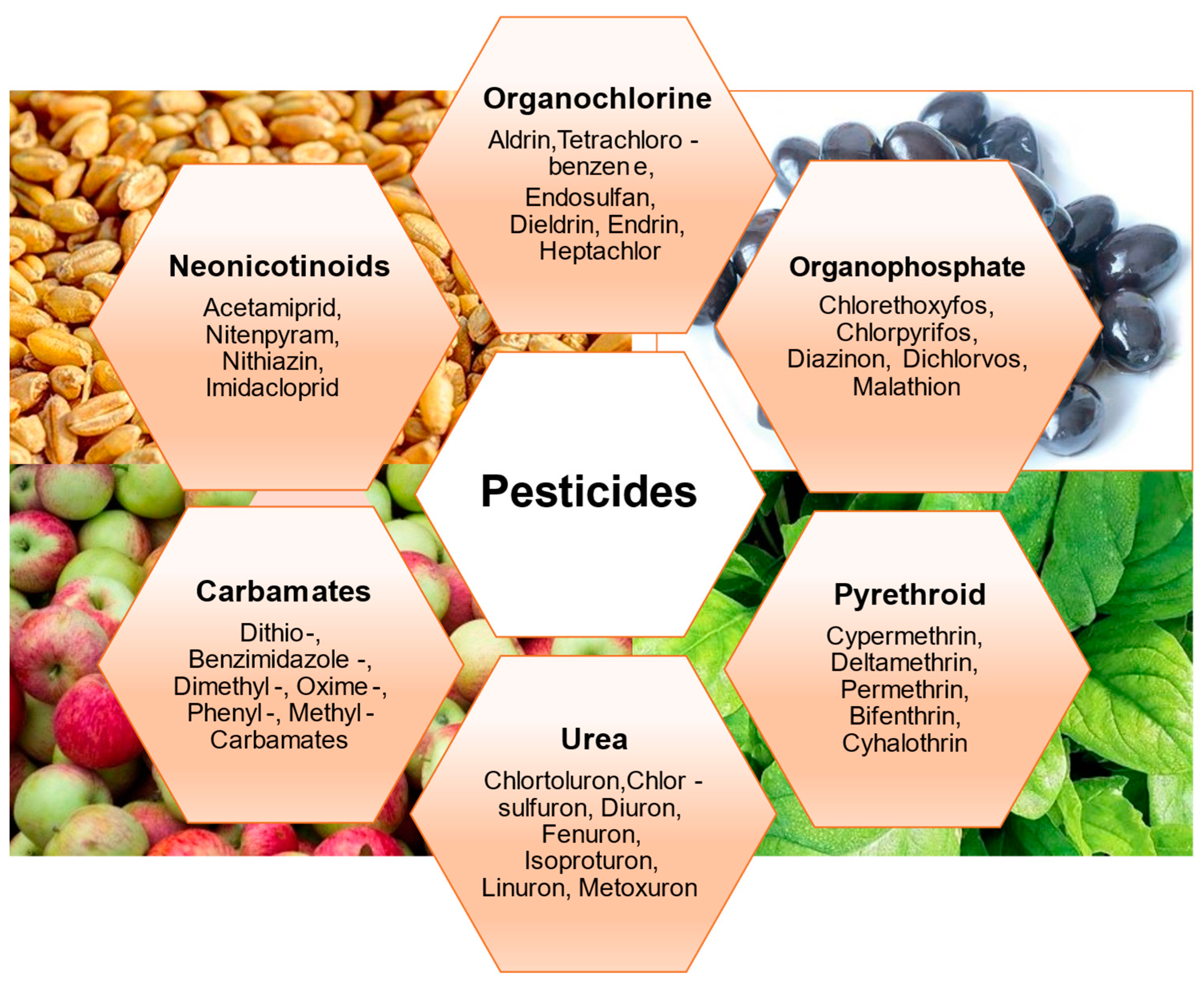
2. Overview of Pesticides in Food
2.1. Organochlorine Pesticides (OCP)
2.2. Organophosphate Pesticides (OPP)
2.3. Pyrethroids
2.4. Urea Pesticides
2.5. Carbamates
2.6. Neonicotinoids
3. Microbial Detoxification of Fermented Foods Containing High Amounts of Pesticides
3.1. Milk and Yogurt
3.2. Pickled Vegetables
3.3. Grains, Flours, and Sourdough
3.4. Tea, Wine, and Fruit Juices
3.5. Meat and Sausages
4. Molecular Mechanisms of Pesticides Degradation
5. Prospects in Fermented Food Detoxification
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ambrus, Á.; Doan, V.V.N.; Szenczi-Cseh, J.; Szemánné-Dobrik, H.; Vásárhelyi, A. Quality Control of Pesticide Residue Measurements and Evaluation of Their Results. Molecules 2023, 28, 954. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.M.; Verma, V.K.; Rawat, B.S.; Kaur, B.; Babu, N.; Sharma, A.; Dewali, S.; Yadav, M.; Kumari, R.; Singh, S.; et al. Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: A comprehensive review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef] [PubMed]
- Méndez, J.M.; Gutiérrez-Fernández, Á.J.; Hardisson, A.; Niebla-Canelo, D.; Alejandro-Vega, S.; Rubio-Armendáriz, C.; Paz-Montelongo, S. Pesticide Residues in Bananas from the Canary Islands. Foods 2023, 12, 437. [Google Scholar] [CrossRef] [PubMed]
- Nasreddine, L.; Parent-Massin, D. Food contamination by metals and pesticides in the European Union. Should we worry? Toxicol. Lett. 2002, 127, 29–41. [Google Scholar] [CrossRef]
- Rather, I.; Koh, W.; Paek, W.; Lim, J. The sources of chemical contaminants in food and their health implications. Front. Pharmacol. 2017, 8, 830. [Google Scholar] [CrossRef]
- Bogialli, S.; Curini, R.; Di Corcia, A.; Laganà, A.; Stabile, A.; Sturchio, E. Development of a multiresidue method for analyzing herbicide and fungicide residues in bovine milk based on solid-phase extraction and liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2006, 1102, 1–10. [Google Scholar] [CrossRef]
- Ozbey, A.; Uygun, U. Behaviour of some organophosphorus pesticide residues in thyme and stinging nettle tea during infusion process. Int. J. Food Sci. Technol. 2007, 42, 380–383. [Google Scholar] [CrossRef]
- Fleurat-Lessard, F.; Chaurand, M.; Marchegay, G.; Abecassis, J. Effects of processing on the distribution of pirimiphos-methyl residues in milling fractions of durum wheat. J. Stored Prod. Res. 2007, 43, 384–395. [Google Scholar] [CrossRef]
- ALS Life Sciences. About Pesticides and Pesticide Residue. Available online: https://www.alsglobal.eu/specialities/pesticides-testing/about-pesticides-and-pesticide-residue (accessed on 12 December 2022).
- EU Pesticides Database. Available online: https://food.ec.europa.eu/plants/pesticides/eu-pesticides-database_en (accessed on 30 January 2023).
- Parte, S.; Mohekar, A.; Kharat, A. Microbial degradation of pesticide: A review. Afr. J. Microbiol. Res. 2017, 24, 992–1012. [Google Scholar] [CrossRef]
- Pesticides Indicators (National—Global—Annual)—FAOSTAT. Available online: https://data.apps.fao.org/catalog/dataset/pesticides-indicators-national-global-annual-faostat/resource/0d25a9a1-6c8f-48e2-8d14-3492b00de4ed (accessed on 29 January 2023).
- Anastasis, C.; George, P.; Panagiotis, D.; Vasileios, F.; Costas, M.; Josep, M.; Benjamin, P.; Despo, F.-K. Ranking of crop plants according to their potential to uptake and accumulate contaminants of emerging concern. Environ. Res. 2019, 170, 422–432. [Google Scholar] [CrossRef]
- Schleiffer, M.; Speiser, B. Presence of pesticides in the environment, transition into organic food, and implications for quality assurance along the European organic food chain—A review. Environ. Poll. 2022, 313, 120116. [Google Scholar] [CrossRef]
- Bajwa, U.; Sandhu, K.S. Effect of handling and processing on pesticide residues in food—A review. J. Food Sci. Technol. 2014, 51, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, J.; Wang, T.; Li, C.; Wu, Z. Effects of ozone treatment on pesticide residues in food: A review. Int. J. Food Sci. Technol. 2019, 54, 301–312. [Google Scholar] [CrossRef]
- Michaels, B.; Gangar, V.; Schattenberg, H.; Blevins, M.; Ayers, T. Effectiveness of cleaning methodologies used for removal of physical, chemical and microbiological residues from produce. Food Serv. Technol. 2003, 3, 9–15. [Google Scholar] [CrossRef]
- Heshmati, A.; Hamidi, M.; Nili-Ahmadabadi, A. Effect of storage, washing, and cooking on the stability of five pesticides in edible fungi of Agaricus bisporus: A degradation kinetic study. Food Sci. Nutr. 2019, 7, 3993–4000. [Google Scholar] [CrossRef]
- Trinder, M.; Bisanz, J.E.; Burton, J.P.; Reid, G. Probiotic lactobacilli: A potential prophylactic treatment for reducing pesticide absorption in humans and wildlife. Benef. Microbes 2015, 6, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-Mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients 2022, 14, 2038. [Google Scholar] [CrossRef]
- Lopes-Ferreira, M.; Maleski, A.L.A.; Balan-Lima, L.; Bernardo, J.T.G.; Hipolito, L.M.; Seni-Silva, A.C.; Batista-Filho, J.; Falcao, M.A.P.; Lima, C. Impact of Pesticides on Human Health in the Last Six Years in Brazil. Int. J. Environ. Res. Public Health 2022, 19, 3198. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-W.; Santos, H.M.; Que, D.E.; Gou, Y.-Y.; Tayo, L.L.; Hsu, Y.-C.; Chen, Y.-B.; Chen, F.-A.; Chao, H.-R.; Huang, K.-L. Association between Organochlorine Pesticide Levels in Breast Milk and Their Effects on Female Reproduction in a Taiwanese Population. Int. J. Environ. Res. Public Health 2018, 15, 931. [Google Scholar] [CrossRef]
- Xie, H.; Liu, R.; Xu, Y.; Liu, X.; Sun, F.; Ma, Y.; Wang, Y. Effect of In Situ Bioremediation of Soil Contaminated with DDT and DDE by Stenotrophomonas sp. Strain DXZ9 and Ryegrass on Soil Microorganism. Microbiol. Res. 2022, 13, 64–86. [Google Scholar] [CrossRef]
- Al-Shamary, N.; Al-Ghouti, M.; Al-Shaikh, I.; Al-Meer, S.; Ahmad, T. Evaluation of pesticide residues of organochlorine in vegetables and fruits in Qatar: Statistical analysis. Environ. Monit. Assess. 2016, 188, 198. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Ke, R.; Gao, W.; Tian, F.; Wang, Y. Analysis of Organochlorine Pesticide Residues in Various Vegetable Oils Collected in Chinese Markets. J. Agric. Food Chem. 2020, 68, 14594–14602. [Google Scholar] [CrossRef] [PubMed]
- Lankondjoa, K.; Gbénonchi, M.; Komla, S. Assessment of Organochlorine Pesticide Residues in Vegetable Samples from Some Agricultural Areas in Togo. Am. J. Analyt. Chem. 2016, 7, 332–341. [Google Scholar] [CrossRef]
- Odewale, G.; Sosan, M.; Oyekunle, J.; Adeleye, A. Human health risk assessment of dichlorodiphenyltrichloroethane (DDT) and hexachlorocyclohexane (HCH) pesticide residues in fruits and vegetables in Nigeria. Environ. Sci. Pollut. Res. 2021, 28, 33133–33145. [Google Scholar] [CrossRef]
- Wang, N.; Cui, Z.; Wang, Y.; Zhang, J. Characteristics and Residual Health Risk of Organochlorine Pesticides in Fresh Vegetables in the Suburb of Changchun, Northeast China. Int. J. Environ. Res. Public Health 2022, 19, 12547. [Google Scholar] [CrossRef]
- Ishaq, Z.; Nawaz, M. Analysis of contaminated milk with organochlorine pesticide residues using gas chromatography. Int. J. Food Prop. 2018, 21, 879–891. [Google Scholar] [CrossRef]
- Akhtar, S.; Ahad, K. Pesticides Residue in Milk and Milk Products: Mini Review. Pak. J. Anal. Environ. Chem. 2017, 18, 37–45. [Google Scholar] [CrossRef]
- Zhuang, M.; Bi, Y.; Han, L.; Qiao, C.; Xie, H.; Qin, F.; Lv, X.; Dong, Q.; Cao, M. Residual pattern, dietary risk assessment and livestock dietary burden of five fungicides on wheat in twelve different regions of China. J. Food Compos. Anal. 2023, 115, 104974. [Google Scholar] [CrossRef]
- Mebdoua, S.; Ounane, G. Evaluation of pesticide residues in wheat grains and its products from Algeria. Food Addit. Contam. B Surveill. 2019, 12, 289–295. [Google Scholar] [CrossRef]
- Kovač, M.; Bulaić, M.; Jakovljević, J.; Nevistić, A.; Rot, T.; Kovač, T.; Dodlek Šarkanj, I.; Šarkanj, B. Mycotoxins, Pesticide Residues, and Heavy Metals Analysis of Croatian Cereals. Microorganisms 2021, 9, 216. [Google Scholar] [CrossRef]
- Galani, Y.J.H.; Houbraken, M.; Wumbei, A.; Djeugap, J.F.; Fotio, D.; Gong, Y.Y.; Spanoghe, P. Monitoring and dietary risk assessment of 81 pesticide residues in 11 local agricultural products from the 3 largest cities of Cameroon. Food Control 2020, 118, 107416. [Google Scholar] [CrossRef]
- Ping, H.; Wang, B.; Li, C.; Li, Y.; Ha, X.; Jia, W.; Li, B.; Ma, Z. Potential health risk of pesticide residues in greenhouse vegetables under modern urban agriculture: A case study in Beijing, China. J. Food Compos. Anal. 2022, 105, 104222. [Google Scholar] [CrossRef]
- Zhang, Q.; Ying, Z.; Tang, T.; Guo, B.; Gu, S.; Fu, L.; Dai, W.; Lin, S. Residual characteristics and potential integrated risk assessment of synthetic pyrethroids in leafy vegetables from Zhejiang in China—Based on a 3-year investigation. Food Chem. 2021, 365, 130389. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cho, Y.-J.; Song, J.-W.; Kim, Y.-J.; Seo, J.-S.; Kim, J.-H. Residue Behavior of Methoxyfenozide and Pymetrozine in Chinese Cabbage and Their Health Risk Assessment. Foods 2022, 11, 2995. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.W.; Moon, J.K. Translocation of chlorpyrifos residue from soil to Korean cabbage. Appl. Biol. Chem. 2018, 61, 145–152. [Google Scholar] [CrossRef]
- Montiel-León, J.; Duy, S.; Munoz, G.; Verner, M.; Hendawi, M.Y.; Moya, H.; Amyot, M.; Sauvé, S. Occurrence of pesticides in fruits and vegetables from organic and conventional agriculture. Food Control 2019, 104, 74–82. [Google Scholar] [CrossRef]
- Calderon, R.; García-Hernández, J.; Palma, P.; Leyva-Morales, J.; Zambrano-Soria, M.; Bastidas-Bastidas, P.; Godoy, M. Assessment of pesticide residues in vegetables commonly consumed in Chile and Mexico: Potential impacts for public health. J. Food Compos. Anal. 2022, 108, 104420. [Google Scholar] [CrossRef]
- EFSA. National summary reports on pesticide residue analysis performed in 2019. EFSA J. 2021, 18, EN-6487. [Google Scholar] [CrossRef]
- Ali, S.; Aziz, M.; Mohamed, S. Determination of Pesticides Residues in Eggplant and Tomatoes from Central Marked in Khartoum State Using Quechers Method and Gas Liquid Chromatography-Mass Spectrometry. Biomed. J. Sci. Tech. Res. 2020, 24, 18165–18173. [Google Scholar] [CrossRef]
- Ahmed, M.A.I.; Khalil, N.; Rahman, T. Carbamate pesticide residues analysis of potato tuber samples using high-performance liquid chromatography (HPLC). J. Environ. Chem. Ecotoxicol. 2014, 6, 1–5. [Google Scholar] [CrossRef]
- Elmarsafy, A.; Kadah, T. Measuring the economic impact and health risks of pesticide residues in potatoes and grapes crops. Egypt. J. Agric. Res. 2018, 96, 1203–1228. [Google Scholar] [CrossRef]
- Huong, D.; Nga, T.; Ha, D. Residue Pesticides (Pyrethroid Group) in Vegetable and Their Health Risk Assessment via Digestion on Consumers in Ha Nam Province, Vietnam. IOP Conf. Ser. Earth Environ. Sci. 2020, 505, 012052. [Google Scholar] [CrossRef]
- Prasopsuk, J.; Laohasiriwong, S.; Promkhambut, A.; Iwai, C. Food safety risk assessment of pesticide residues in Chinese kale grown in Khon Kaen province, northeast Thailand. Agric. Nat. Resour. 2020, 54, 343–350. [Google Scholar]
- Neylon, J.; Fuller, J.N.; van der Poel, C.; Church, J.E.; Dworkin, S. Organophosphate Insecticide Toxicity in Neural Development, Cognition, Behaviour and Degeneration: Insights from Zebrafish. J. Dev. Biol. 2022, 10, 49. [Google Scholar] [CrossRef]
- Delińska, K.; Yavir, K.; Kloskowski, A. Head-Space SPME for the Analysis of Organophosphorus Insecticides by Novel Silica IL-Based Fibers in Real Samples. Molecules 2022, 27, 4688. [Google Scholar] [CrossRef]
- Yousefi, M.; Shadnoush, M.; Sohrabvandi, S.; Khorshidian, N.; Mohammadi, M.; Mortazavian, A. Probiotics as potential detoxification tools for mitigation of pesticides: A mini review. Food Sci. Technol. 2021, 56, 2078–2087. [Google Scholar] [CrossRef]
- Caba, I.-C.; Ștreangă, V.; Dobrin, M.-E.; Jităreanu, C.; Jităreanu, A.; Profire, B.-Ș.; Apotrosoaei, M.; Focșa, A.-V.; Caba, B.; Agoroaei, L. Clinical Assessment of Acute Organophosphorus Pesticide Poisoning in Pediatric Patients Admitted to the Toxicology Emergency Department. Toxics 2022, 10, 582. [Google Scholar] [CrossRef]
- Li, X.; Yang, X.; Zheng, X.; Bai, M.; Hu, D. Review on Structures of Pesticide Targets. Int. J. Mol. Sci. 2020, 21, 7144. [Google Scholar] [CrossRef]
- Kolakowski, B.M.; Miller, L.; Murray, A.; Leclair, A.; Bietlot, H.; van de Riet, J.M. Analysis of Glyphosate Residues in Foods from the Canadian Retail Markets between 2015 and 2017. J. Agric. Food Chem. 2020, 68, 5201–5211. [Google Scholar] [CrossRef]
- Liang, S.; Zhao, Z.; Fan, C.; Xu, J.; Li, H.; Chang, Q.; Pang, G. Fipronil residues and risk assessment of Chinese marketed fruits and vegetables: A long-term investigation over 6 years. Food Control 2019, 106, 106734. [Google Scholar] [CrossRef]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid pesticide residues in the global environment: An overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, W.; Miao, J.; Sun, C.; Wang, Q.; Pan, L. Pyrethroids contamination and health risk assessment in seafood collected from the coast of Shandong, China. Mar. Pollut. Bull. 2023, 186, 114442. [Google Scholar] [CrossRef] [PubMed]
- Morais, S.; Correia, M.; Domingues, V.; Delerue-Matos, C. Urea Pesticides. In Pesticides—Strategies for Pesticides Analysis; Stoytcheva, M., Ed.; InTech: London, UK, 2011; pp. 241–262. [Google Scholar]
- Lee, Y.-P.; Mendoza-Huizar, L.H. Chemical Reactivity of Isoproturon, Diuron, Linuron, and Chlorotoluron Herbicides in Aqueous Phase: A Theoretical Quantum Study Employing Global and Local Reactivity Descriptors. J. Chem. 2015, 2015, 751527. [Google Scholar] [CrossRef]
- Sagratini, G.; Manes, J.; Giardina, D.; Damiani, P.; Pico, Y. Analysis of carbarnate and phenylurea pesticide residues in fruit juices by solid-phase microextraction and liquid chromatography-mass spectrometry. J. Chromatogr. A. 2007, 1147, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Mou, R.X.; Chen, M.X.; Zhi, J.L. Simultaneous determination of 15 phenylurea herbicides in rice and corn using HPLC with fluorescence detection combined with UV decomposition and post-column derivatization. J. Chromatogr. B Analyt. Technol. Biomed Life Sci. 2008, 875, 437–443. [Google Scholar] [CrossRef]
- Paiga, P.; Morais, S.; Correia, M.; Alves, A.; Delerue-Matos, C. Screening of Carbamates and Ureas in Fresh and Processed Tomato Samples using Microwave Assisted Extraction and Liquid Chromatography. Anal. Lett. 2009, 42, 265–283. [Google Scholar] [CrossRef]
- Paiga, P.; Morais, S.; Correia, M.; Delerue-Matos, C.; Alves, A. Determination of carbamate and urea pesticide residues in fresh vegetables using microwave assisted extraction and liquid chromatography. Int. J. Environ. Anal. Chem. 2009, 89, 199–210. [Google Scholar] [CrossRef]
- Moreira, S.; Silva, R.; Carrageta, D.F.; Alves, M.G.; Seco-Rovira, V.; Oliveira, P.F.; de Lourdes Pereira, M. Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System. Int. J. Mol. Sci. 2022, 23, 8206. [Google Scholar] [CrossRef]
- Ponce-Vejar, G.; Ramos de Robles, S.L.; Macias-Macias, J.O.; Petukhova, T.; Guzman-Novoa, E. Detection and Concentration of Neonicotinoids and Other Pesticides in Honey from Honey Bee Colonies Located in Regions That Differ in Agricultural Practices: Implications for Human and Bee Health. Int. J. Environ. Res. Public Health 2022, 19, 8199. [Google Scholar] [CrossRef]
- Xu, M.; Portier, L.; Bovee, T.; Zhao, Y.; Guo, Y.; Peters, J. Neonicotinoid Microsphere Immunosensing for Profiling Applications in Honeybees and Bee-Related Matrices. Biosensors 2022, 12, 792. [Google Scholar] [CrossRef]
- Mugo, S.M.; Lu, W.; Robertson, S.V. Molecularly Imprinted Polymer-Modified Microneedle Sensor for the Detection of Imidacloprid Pesticides in Food Samples. Sensors 2022, 22, 8492. [Google Scholar] [CrossRef]
- Lachat, L.; Glauser, G. Development and Validation of an Ultra-Sensitive UHPLC-MS/MS Method for Neonicotinoid Analysis in Milk. J. Agric. Food Chem. 2018, 66, 8639–8646. [Google Scholar] [CrossRef]
- Lu, C.; Chang, C.; Palmer, C.; Zhao, M.; Zhang, Q. Neonicotinoid Residues in Fruits and Vegetables: An Integrated Dietary Exposure Assessment Approach. Environ. Sci. Technol. 2018, 52, 3175–3184. [Google Scholar] [CrossRef]
- Audus, J. The Biological Detoxication of 2: 4-Dichlorophenoxyacetic. Acid in Soil. Plant Soil. 1949, 2, 31–36. [Google Scholar] [CrossRef]
- Wahla, V.; Shukla, S. Role of Microorganisms in Bioremediation of Pesticides. In Handbook of Research on Microbial Tools for Environmental Waste Management; Pathak, V., Navneet, M., Eds.; IGI Global: Hershey, PA, USA, 2018; pp. 164–189. [Google Scholar]
- Singh, B.; Walker, A. Microbial degradation of organophosphorus compounds. FEMS Microbiol. Rev. 2006, 30, 428–471. [Google Scholar] [CrossRef] [PubMed]
- John, E.M.; Shaike, J. Chlorpyrifos: Pollution and remediation. Environ Chem. Lett. 2015, 13, 269–291. [Google Scholar] [CrossRef]
- Abou-Arab, A. Effect of Ras cheese manufacturing on the stability of DDT and its metabolites. Food Chem. 1997, 59, 115–119. [Google Scholar] [CrossRef]
- Navarro, S.; Pérez, G.; Navarro, G.; Vela, N. Decline of pesticide residues from barley to malt. Food Addit. Contam. 2007, 24, 851–859. [Google Scholar] [CrossRef]
- Rajashekar, K.; Kondal, K.; Narasimha, K.; Sudhakar, K. Effect of processing of milk into products on the residue levels of certain pesticides. J. Food. Sci. Technol. 2007, 44, 551–552. [Google Scholar]
- Jung, K.; Park, Y.; Kim, H.; Kang, M.; Yang, Y.; Kang, A.; Chun, K.; Park, Y. Removal effects of Bifenthrin and Metalaxyl pesticides during preparation and fermentation of Baechu Kimchi. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1258–1264. [Google Scholar] [CrossRef]
- Bo, L.-Y.; Zhang, Y.-H.; Zhao, X.-H. Degradation kinetics of seven organophosphorus pesticides in milk during yoghurt processing. J. Serb. Chem. Soc. 2011, 76, 353–362. [Google Scholar] [CrossRef]
- Čuš, F.; Česnik, B.; Bolta, V.; Gregorčič, A. Pesticide residues in grapes and during vinification process. Food Cont. 2010, 21, 1512–1518. [Google Scholar] [CrossRef]
- Książek-Trela, P.; Szpyrka, E. The effect of natural and biological pesticides. Plant Prot. Sci. 2022, 58, 273–291. [Google Scholar] [CrossRef]
- Nandhini, A.; Harshiny, M.; Gummadi, S. Chlorpyrifos in environment and foods: A critical review of detection methods and degradation pathways. Environ. Sci. Process. Impacts 2021, 23, 1255–1277. [Google Scholar] [CrossRef] [PubMed]
- Wyszkowska, J.; Zaborowska, M.; Kucharski, J. The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities. Int. J. Mol. Sci. 2023, 24, 2892. [Google Scholar] [CrossRef]
- Bouhafs, L.; Moudilou, E.; Exbrayat, J.; Lahouel, M.; Idoui, T. Protective effects of probiotic Lactobacillus plantarum BJ0021 on liver and kidney oxidative stress and apoptosis induced by endosulfan in pregnant rats. Ren. Fail. 2015, 37, 1370–1378. [Google Scholar] [CrossRef]
- Trinder, M.; McDowell, T.; Daisley, B.; Ali, S.; Leong, H.; Sumarah, M.; Reid, G. Probiotic Lactobacillus rhamnosus reduces organophosphate pesticide absorption and toxicity to Drosophila melanogaster. Appl. Environ. Microbiol. 2016, 82, 6204–6213. [Google Scholar] [CrossRef]
- Villarini, M.; Caldini, G.; Moretti, M.; Trotta, F.; Pasquini, R.; Cenci, G. Modulatory activity of Lactobacillus casei strain on 1,2-dimethylhydrazine-induced genotoxicity in rats. Environ. Mol. Mutagen. 2008, 49, 129–199. [Google Scholar] [CrossRef]
- Kallman, B.J.; Andrews, A.K. Reductive dechlorination of DDT to DDD by yeast. Science 1963, 141, 1050. [Google Scholar] [CrossRef]
- Duan, J.; Cheng, Z.; Bi, J.; Xu, Y. Residue behavior of organochlorine pesticides during the production process of yogurt and cheese. Food Chem. 2018, 254, 119–124. [Google Scholar] [CrossRef]
- Witczak, A.; Mituniewicz-Małek, A. The comparison of probiotic monocultures influence on organochlorine pesticides changes in fermented beverages from cow and goat milk during cold storage. Mljekarstvo 2019, 69, 172–181. [Google Scholar] [CrossRef]
- Yuan, S.; Yang, F.; Yu, H.; Xie, Y.; Guo, Y.; Yao, W. Biodegradation of the organophosphate dimethoate by Lactobacillus plantarum during milk fermentation. Food Chem. 2021, 360, 130042. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhao, X. Susceptibility of nine organophosphorus pesticides in skimmed milk towards inoculated lactic acid bacteria and yogurt starters. J. Sci. Food Agric. 2015, 95, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, D.; Liu, Q.; Zhao, H. Enhanced degradation of five organophosphorus pesticides in skimmed milk by lactic acid bacteria and its potential relationship with phosphatase production. Food Chem. 2014, 164, 173–178. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J. A brief study on the degradation kinetics of seven organophosphorus pesticides in skimmed milk cultured with Lactobacillus spp. at 42 °C. Food Chem. 2012, 131, 300–304. [Google Scholar] [CrossRef]
- Nasution, L.; Bakti, D.; Agusnar, H.; Harahap, E.M. Role of Lactobacillus sakei strain pro7 to reduce dichlorodiphenyl trichloroethane level. J. Phys. 2018, 1116, 042025. [Google Scholar] [CrossRef]
- Cho, K.; Math, R.; Islam, S.; Lim, W.; Hong, S.; Kim, J.; Yum, M.; Cho, J.; Yum, H. Biodegradation of chlorpyrifos by lactic acid bacteria during kimchi fermentation. J. Agric. Food Chem. 2009, 57, 1882–1889. [Google Scholar] [CrossRef]
- Islam, S.; Math, R.; Cho, K.; Lim, W.; Hong, S.; Kim, J.; Yun, M.; Cho, J.; Yun, H. Organophosphorus Hydrolase (OpdB) of Lactobacillus brevis WCP902 from Kimchi is able to degrade organophosphorus pesticides. J. Agric. Food Chem. 2010, 58, 5380–5386. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, F.; Zhao, H. The potencies of three microorganisms to dissipate four organophosphorus pesticides in three food materials during traditional fermentation. J. Food Sci. Technol. 2015, 52, 7353–7360. [Google Scholar] [CrossRef]
- Kumral, A.Y.; Kumral, N.A.; Gurbuz, O. Chlorpyrifos and deltamethrin degradation potentials of two Lactobacillus plantarum (Orla-Jensen, 1919) (Lactobacillales: Lactobacillaceae) strains. Türk. Entomol. Derg. 2020, 44, 165–176. [Google Scholar] [CrossRef]
- Maden, B.; Kumral, A.Y. Degradation Trends of Some Insecticides and Microbial Changes during Sauerkraut Fermentation under Laboratory Conditions. J. Agric. Food Chem. 2020, 68, 14988–14995. [Google Scholar] [CrossRef]
- Kumral, A.Y.; Kumral, N.A.; Kolcu, A.; Maden, B.; Artik, B. Simulation Study for the Degradation of Some Insecticides during Different Black Table Olive Processes. ACS Omega 2020, 5, 14164–14172. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, D.; Zhao, H.; Song, Y.; Liu, L.; Li, N. Biodegradation of two organophosphorus pesticides in whole corn silage as affected by the cultured Lactobacillus plantarum. Biotech 2016, 6, 73. [Google Scholar] [CrossRef]
- Đorđević, T.; Šiler-Marinković, S.; Đurović-Pejčev, R.; Dimitrijević-Branković, S.; Gajić Umiljendić, J. Dissipation of pirimiphos-methyl during wheat fermentation by Lactobacillus plantarum. Lett. Appl. Microbiol. 2013, 57, 412–419. [Google Scholar] [CrossRef]
- Low, L.; Shaw, C.; Gerrard, A. The effect of Saccharomyces cerevisiae on the stability of the herbicide glyphosate during bread leavening. Lett. Appl. Microbiol. 2004, 40, 133–137. [Google Scholar] [CrossRef]
- Sharma, J.; Satya, S.; Kumar, V.; Tewary, D.K. Dissipation of pesticides during bread-making. Chem. Health Saf. 2005, 12, 17–22. [Google Scholar] [CrossRef]
- Dorđević, T.M.; Siler-Marinkovic, S.S.; Durovic, R.D.; Dimitrijevic-Brankovic, S.I.; Gajic Umiljendic, J.S. Stability of the pyrethroid pesticide bifenthrin in milled wheat during thermal processing, yeast and lactic acid fermentation, and storage. J. Sci. Food Agric. 2013, 93, 3377–3383. [Google Scholar] [CrossRef]
- Liu, F.; Bai, J.; Huang, W.; Li, F.; Ke, W.; Zhang, Y.; Xie, D.; Zhang, B.; Guo, X. Characterization of a novel beta-cypermethrin-degrading strain of Lactobacillus pentosus 3-27 and its effects on bioremediation and the bacterial community of contaminated alfalfa silage. J. Hazard. Mater. 2022, 423 Pt A, 127101. [Google Scholar] [CrossRef]
- Đorđević, T.M.; Đurović-Pejčev, R.D. The potency of Saccharomyces cerevisiae and Lactobacillus plantarum to dissipate organophosphorus pesticides in wheat during fermentation. J. Food Sci. Technol. 2016, 53, 4205–4215. [Google Scholar] [CrossRef]
- Deng, W.; Lin, D.; Yao, K.H.; Wang, Z.; Li, J.; Zou, L.; Han, X.; Zhou, K.; He, L.; Hu, X.; et al. Characterization of a novel β-cypermethrin-degrading Aspergillus niger YAT strain and the biochemical degradation pathway of β-cypermethrin. Appl. Microbiol. Biotechnol. 2015, 99, 8187–8198. [Google Scholar] [CrossRef]
- Gava, A.; Emer, C.; Ficagna, E.; de Andrade, S.F.; Fuentefria, A.M. Occurrence and impact of fungicides residues on fermentation during wine production—A review. Food Addit. Contam. A 2021, 38, 943–961. [Google Scholar] [CrossRef]
- Grimalt, S.; Dehouck, P. Review of analytical methods for the determination of pesticide residues in grapes. J. Chromatogr. A 2016, 1433, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhou, Y.; Zhan, R.; Zhu, L.; Chen, H.; Ma, Z.; Chen, X.; Lu, Y. Effects of Different Pesticides on the Brewing of Wine Investigated by GC-MS-Based Metabolomics. Metabolites 2022, 12, 485. [Google Scholar] [CrossRef] [PubMed]
- Ruediger, A.; Pardo, H.; Sas, N.; Godden, W.; Pollnitz, P. Fate of Pesticides during the Winemaking Process in Relation to Malolactic Fermentation. J. Agric. Food Chem. 2005, 53, 3023–3026. [Google Scholar] [CrossRef] [PubMed]
- González-Rodríguez, M.; Cancho-Grande, B.; Torrado-Agrasar, A.; Simal-Gándara, J.; Mazaira-Pérez, J. Evolution of tebuconazole residues through the winemaking process of Mencía grapes. Food Chem. 2009, 117, 529–537. [Google Scholar] [CrossRef]
- Rezaei, F.; Nejati, R.; Sayadi, M.; Nematollahi, A. Diazinon reduction in apple juice using probiotic bacteria during fermentation and storage under refrigeration. Environ. Sci. Pollut. Res. Int. 2021, 28, 61213–61224. [Google Scholar] [CrossRef] [PubMed]
- Abou-Arab, K. Degradation of organochlorine pesticides by meat starter in liquid media and fermented sausage. Food Chem. Toxicol. 2002, 40, 33–41. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, L.; Li, F.; Xiao, M.; Lin, D.; Long, X.; Wu, Z. Microbial Degradation of Pesticide Residues and an Emphasis on the Degradation of Cypermethrin and 3-phenoxy Benzoic Acid: A Review. Molecules 2018, 23, 2313. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Karpouzas, D.G.; Singh, B.K. Microbial degradation of organophosphorus xenobiotics: Metabolic pathways and molecular basis. Adv. Microb. Physiol. 2006, 51, 119–185. [Google Scholar] [CrossRef]
- Bhatt, P.; Zhou, X.; Huang, Y.; Zhang, W.; Chen, S. Characterization of the role of esterases in the biodegradation of organophosphate, carbamate, and pyrethroid pesticides. J. Hazard. Mater. 2021, 411, 125026. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, M.F.; Bellinger, D.C.; Wright, R.O.; Weisskopf, M.G. Attention-deficit/hyperactivity disorder and urinary metabolites of organophosphate pesticides. Pediatrics 2010, 125, e1270–e1277. [Google Scholar] [CrossRef]
- Bouchard, M.F.; Chevrier, J.; Harley, K.G.; Kogut, K.; Vedar, M.; Calderon, N.; Trujillo, C.; Johnson, C.; Bradman, A.; Barr, D.B.; et al. Prenatal exposure to organophosphate pesticides and IQ in 7-year-old children. Environ. Health Perspect. 2011, 119, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Gillezeau, C.; Alpert, N.; Joshi, P.; Taioli, E. Urinary Dialkylphosphate Metabolite Levels in US Adults—National Health and Nutrition Examination Survey 1999–2008. Int. J. Environ. Res. Public Health 2019, 16, 4605. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Ryan, L.; Barr, D.B.; Hauser, R. Exposure to nonpersistent insecticides and male reproductive hormones. Epidemiology 2006, 17, 61–68. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, W.; Zhu, X.; Wu, J.; Wu, X. Organophosphorus insecticides exposure and sex hormones in general U.S. population: A cross-sectional study. Environ Res. 2022, 215, 114384. [Google Scholar] [CrossRef]
- Gilani, R.A.; Rafique, M.; Rehman, A.; Munis, M.F.; Rehman, S.U.; Chaudhary, H.J. Biodegradation of chlorpyrifos by bacterial genus Pseudomonas. J. Basic. Microbiol. 2016, 56, 105–119. [Google Scholar] [CrossRef]
- Haque, M.A.; Hong, S.Y.; Hwang, C.E.; Kim, S.C.; Cho, K.M. Cloning of an organophosphorus hydrolase (opdD) gene of Lactobacillus sakei WCP904 isolated from chlorpyrifos-impregnated kimchi and hydrolysis activities of its gene product for organophosphorus pesticides. Appl. Biol. Chem. 2018, 61, 643–651. [Google Scholar] [CrossRef]
- Haque, M.A.; Hwang, C.E.; Kim, S.C.; Cho, D.Y.; Lee, H.Y.; Cho, K.M.; Lee, J.H. Biodegradation of organophosphorus insecticides by two organophosphorus hydrolase genes (opdA and opdE) from isolated Leuconostoc mesenteroides WCP307 of kimchi origin. Process Biochem. 2020, 94, 340–348. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, H.Y.; Cho, D.Y.; Kim, M.J.; Jung, J.G.; Jeong, E.H.; Haque, M.A.; Cho, K.M. Biodegradable properties of organophosphorus insecticides by the potential probiotic Lactobacillus plantarum WCP931 with a degrading gene (opdC). Appl. Biol. Chem. 2021, 64, 62. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Wu, T.-H.; Yang, Y.; Zhu, C.-L.; Ding, C.-L.; Dai, C.-C. Binding and detoxification of chlorpyrifos by lactic acid bacteria on rice straw silage fermentation. J. Environ. Sci. Health B. 2016, 51, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-H.; Yu, X.-X.; Jin, X.; Wang, Y.-T.; Zhao, D.-J.; Zhang, P.; Sun, G.-M.; Zhang, Y.-H. Purification and characterization of alkaline phosphatase from lactic acid bacteria. RSC Adv. 2019, 9, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, C.; Yu, H.; Xie, Y.; Guo, Y.; Yao, W. Screening of lactic acid bacteria for degrading organophosphorus pesticides and their potential protective effects against pesticide toxicity. LWT 2021, 147, 111672. [Google Scholar] [CrossRef]
- Pinto, G.D.A.; Castro, I.M.; Miguel, M.A.L.; Koblitz, M.G.B. Lactic acid bacteria—Promising technology for organophosphate degradation in food: A pilot study. LWT 2019, 110, 353–359. [Google Scholar] [CrossRef]
- Li, C.; Ma, Y.; Mi, Z.; Huo, R.; Zhou, T.; Hai, H.; Kwok, L.-y.; Sun, Z.; Chen, Y.; Zhang, H. Screening for Lactobacillus plantarum Strains That Possess Organophosphorus Pesticide-Degrading Activity and Metabolomic Analysis of Phorate Degradation. Front. Microbiol. 2018, 9, 2048. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Qiu, L.; Yu, L.; Zhang, S.; de Toledo, R.A.; Shim, H.; Wang, S. Microbial transformation of chiral organohalides: Distribution, microorganisms and mechanisms. J. Hazard. Mater. 2019, 368, 849–861. [Google Scholar] [CrossRef]
- Mishra, S.; Pang, S.; Zhang, W.; Lin, Z.; Bhatt, P.; Chen, S. Insights into the microbial degradation and biochemical mechanisms of carbamates. Chemosphere 2021, 279, 130500. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Lin, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Insights Into the Microbial Degradation and Biochemical Mechanisms of Neonicotinoids. Front. Microbiol. 2020, 19, 868. [Google Scholar] [CrossRef]
- Bose, S.; Kumar, P.S.; Vo, D.N. A review on the microbial degradation of chlorpyrifos and its metabolite TCP. Chemosphere 2021, 283, 131447. [Google Scholar] [CrossRef]
- Petkova, M.; Gotcheva, V.; Dimova, M.; Bartkiene, E.; Rocha, J.M.; Angelov, A. Screening of Lactiplantibacillus plantarum Strains from Sourdoughs for Biosuppression of Pseudomonas syringae pv. syringae and Botrytis cinerea in Table Grapes. Microorganisms 2022, 10, 2094. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Zokaityte, E.; Varinauskaite, I.; Pileckaite, G.; Paskeviciute, L.; Rutkauskaite, G.; Kanaporis, T.; et al. Plants and Lactic Acid Bacteria Combination for New Antimicrobial and Antioxidant Properties Product Development in a Sustainable Manner. Foods 2020, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Küley, E.; Özyurt, G.; Özogul, I.; Boga, M.; Akyol, I.; Rocha, J.M.; Özogul, F. The Role of Selected Lactic Acid Bacteria on Organic Acid Accumulation during Wet and Spray-Dried Fish-based Silages. Contributions to the Winning Combination of Microbial Food Safety and Environmental Sustainability. Microorganisms 2020, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Mahony, J.; van Sinderen, D. Broad-spectrum antifungal-producing lactic acid bacteria and their application in fruit models. Folia Microbiol. 2012, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic acid bacteria as antibacterial agents to extend the shelf life of fresh and minimally processed fruits and vegetables: Quality and safety aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Leska, A.; Nowak, A.; Miskiewicz, K.; Rosicka-Kaczmarek, J. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment—An In Vitro Study. Cells 2022, 11, 3743. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant properties of probiotic bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Palanisamy, M.; Vijila, K. Potential Protective Role of Probiotic Strains of lactobacilli Against Pesticide Toxicity. J. Curr. Crop. Sci. Technol. 2022, 109, 10–12. [Google Scholar] [CrossRef]
- Feng, P.; Ye, Z.; Kakade, A.; Virk, A.K.; Li, X.; Liu, P. A Review on Gut Remediation of Selected Environmental Contaminants: Possible Roles of Probiotics and Gut Microbiota. Nutrients 2019, 11, 22. [Google Scholar] [CrossRef]
- Huang, R.; Wu, F.; Zhou, Q.; Wei, W.; Yue, J.; Xiao, B.; Luo, Z. Lactobacillus and intestinal diseases: Mechanisms of action and clinical applications. Microbiol. Res. 2022, 260, 127019. [Google Scholar] [CrossRef]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Narbad, A.; Chen, W. A mixture of Lactobacillus species isolated from traditional fermented foods promote recovery from antibiotic-induced intestinal disruption in mice. J. Appl. Microbiol. 2018, 124, 842–854. [Google Scholar] [CrossRef] [PubMed]
- Rajawat, N.K.; Bhardwaj, K.; Mathur, N. Risk of Parkinson disease associated with pesticide exposure and protection by probiotics. Mater. Today Proc. 2022, 69, A1–A11. [Google Scholar] [CrossRef]
- Baralić, K.; Živančević, K.; Bozic, D.; Đukić-Ćosić, D. Probiotic cultures as a potential protective strategy against the toxicity of environmentally relevant chemicals: State-of-the-art knowledge. Food Chem. Toxicol. 2023, 172, 113582. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Food | Pesticide Name | Pesticide Class | Toxic Effects on Human Health | Content * (mg/kg) | Reference |
|---|---|---|---|---|---|
| Wheat | Flutriafol | Fungicide | Liver function disorders | 0.350 | [31] |
| Chlorpyrifos | Insecticide | Headache, blurred vision, coma, death | 0.338 | [32] | |
| Cypermethrin | Insecticide | Irritation to the skin, numbness, tingling, incoordination, death | 0.058 | [33] | |
| Maize | Metribuzin | Herbicide | Stomach aches, fatigue, depression, disturbance in kidneys | 1.390 | [34] |
| Thiofanox | Insecticide | Cancerogen | 0.083 | [33] | |
| Chinese cabbage | Chlorpyrifos | Insecticide Insecticide Insecticide Insecticide | Headache, blurred vision, coma, death | 0.48 | [35] |
| Bifenthrin | Endocrine disrupting effects | 1.19 | [36] | ||
| Methoxyfenozide | Liver, and thyroid systems toxicity | 1.12 | [37] | ||
| Pymetrozine | Causes diabetes | 3.02 | [38] | ||
| Tomato | Chlorates | Herbicide | Gastritis, toxic nephritis, hemolysis, methemoglobinemia, hemoglobinuria | 0.450 | [39] |
| Spinach | Methamidophos | Insecticide | Nausea, vomiting, weakness, paralysis | 2.948 | [40] |
| Peppers | Chlordecone | Insecticide | Carcinogenic, with neurological, reproductive, and developmental toxicity | 0.510 | [41] |
| Eggplant | Carbaryl | Insecticide | Sweating, nausea, vomiting, abdominal pain | 1.052 | [42] |
| Potato | Kresoxim | Fungicide | Carcinogenic | 0.180 | [43] |
| Peach | Carbofuran | Insecticide | Endocrine system disruptor | 1.660 | [44] |
| Leaf mustard | Deltamethrin | Insecticide | Headaches and dizziness | 0.661 | [45] |
| Chinese kale | Cypermethrin | Insecticide | Numbness, incoordination, death | 8.79 | [46] |
| Groundnuts | Cymoxanil | Fungicide | Lungs hyperplasia, gliosis, spongiosis | 1.46 | [34] [34] |
| Egusi seeds | Atrazine | Herbicide | Reproductive system disorders | 1.41 |
| Source | Species and Strain | Pesticide | Reduction | Reference |
|---|---|---|---|---|
| Egypt Ras cheese, milk | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, yeasts | DDT | 40.6% at 0.1 mg/kg fat 33.9% at 1.0 mg/kg fat 25.5% at 10.0 mg/kg fat | [73] |
| Yogurt, cheese | Lactobacillus acidophilus, L. delbrueckii subsp. bulgaricus, Lactiplantibacillus plantarum, Lacticaseibacillus rhamnosus, Lacticaseibacillus casei, S. thermophilus, Bifidobacterium bifidum | α-hexachlorocyclohexane (HCH), hexachlorobenzene (HCB), γ-HCH, β-chlordane, α-chlordane | 37.0–50.9% after 12 h at 20 μg/kg for all | [85] |
| Fermented beverages from cow and goat milk (bio-yogurts) | L. acidophilus LA-5, Bif. animalis subsp. lactis BB-12 | αHCH, βHCH, γHCH, 1,1-bis-(4-chlorophenyl)-2,2-dichloroethene (pp’DDE), 1-chloro-4-[2,2-dichloro-1-(4-chlorophenyl) ethyl] benzene, 1,1′-(2,2,2-trichloroethane-1,1-diyl) bis(4-chlorobenzene) | up to 48.6% (heptachlor) and 54.7% (pp’DDE) in goat milk bio-yogurts after 14 days of cold storage when both cultures were used (synergistic effect) | [86] |
| Yogurt | L. delbrueckii subsp. bulgaricus, S. thermophilus | Dimethoate, fenthion, malathion, methyl parathion, monocrotophos, phorate, trichlorphon | 9.2–17.1% after 4 h at 1.5 mg/kg for all except malathion | [77] |
| Milk | L. plantarum CICC20261 | Dimethoate | 81.28% at 50 mg/kg for 24 h | [87] |
| Skimmed milk | L. delbrueckii subsp. bulgaricus, L. acidophilus, Lc. casei, Lc. rhamnosus, S. thermophilus | Chlorpyrifos, chlorpyrifos-methyl, diazinon, dichlorvos, fenthion, malathion, phorate, pirimiphos-methyl, trichlorphon | up to 64.6% for dichlorvos with L. bulgaricus at 1 mg/kg for 24 h | [88] |
| Skimmed milk | Lp. plantarum strains 1.0317, 1.0624, 1.0315, 1.066, Levilactobacillus brevis 1.0209, Lactobacillus helveticus strains 1.0203 and 1.9204, Lactobacillus lactis 4.0611, L. delbrueckii subsp. bulgaricus L6, S. thermophilus 3.0503 | Chlorpyrifos, diazinon, fenitrothion, malathion, methyl parathion | ≈50% at 0.6 mg/kg diazinon for 24 h with Lev. brevis 1.0209 | [89] |
| Skimmed milk | L. bulgaricus, Lacticaseibacillus paracasei, Lp. plantarum | Dimethoate, fenthion, malathion, methyl parathion, monocrotophos, phorate, trichlorphon | from 20.9% (methyl parathion with Lc. paracasei) to 46.9% (malathion with Lp. plantarum) at 1.2 mg/kg for 24 h | [90] |
| Source | Species and Strain | Pesticide | Reduction | Reference |
|---|---|---|---|---|
| Kimchi | Leuconostoc mesenteroides WCP907, Lev. brevis WCP902, Lp. plantarum WCP931, La. sakei WCP904 | Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | 100% chlorpyrifos for 9 days at 200 mg/L initial concentration | [92] |
| Kimchi | Lev. brevis WCP902 | Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | ≈75% chlorpyrifos for 9 days at 100 mg/L initial concentration | [93] |
| Pickled Chinese cabbage | Lp. plantarum | Chlorpyrifos, dichlorvos, phorate, trichlorphon | 6.5–18% more compared to normal fermentation after 42 days at 1 mg/kg; <1% of all except chlorpyrifos (≈4%) in the end | [94] |
| Black olives | Lp. plantarum (LB-1 and LB-2) | Chlorpyrifos, deltamethrin | 96% for chlorpyrifos after 3 days and 86% for deltamethrin after 10 days for LB-1 at 100 mg/L | [95] |
| Sauerkraut | Lp. plantarum 112 | λ-cyhalothrin, malathion, chlorpyrifos-methyl | 13–19% for λ-cyhalothrin (2 mg/kg) after 14 days; 34–59% for the other two (2–4 mg/kg), but lower than natural fermentation (69–98%) | [96] |
| Black olives | Lp. plantarum 112, Lp. plantarum 123 | Deltamethrin, dimethoate, imidacloprid | 2–8% greater compared to natural fermentation after 60 days at 25–350 g/L initially | [97] |
| Source | Species and Strain | Pesticide | Reduction | Reference |
|---|---|---|---|---|
| Corn silage | Lp. plantarum 1.0315, Lp. plantarum 1.0624, Lp. plantarum 1.0622 | Chlorpyrifos, phorate | 33.4% for phorate (0.36 mg/kg) after 10 weeks, but very close to control fermentation (26.2%) | [98] |
| Bread wheat | Lp. plantarum | Pirimiphos, pirimiphos-methyl | 15–34% pirimiphos-methyl (5 mg/kg) for 48 h | [99] |
| Bread | Saccharomyces cerevisiae | Glyphosate | 21% for 1 h at 50 mg/kg | [100] |
| Bread | Saccharomyces cerevisiae | Endosulfan, deltamethrin, malathion, propiaconazole, chlorpyriphos, hexaconazole | 75–89% at 1 mg/kg; 47–70% at 4 mg/kg; after 12 h fermentation at 30 °C and 20 min baking at 80 °C | [101] |
| Wheat flour | Lp. plantarum | Bifenthrin | 42% at 0.5 mg/kg, only 18% at 2.5 mg/kg | [102] |
| Alfalfa silage | Lp. pentosus 3–27 | Beta-cypermethrin | 96% at 50 mg/L for 4 days | [103] |
| Wheat | Saccharomyces cerevisiae | Pirimiphos methyl | 48.8% at 5 mg/kg for 72 h | [104] |
| Wheat | Lp. plantarum | Chlorpyrifos methyl | 56.7% at 3 mg/kg for 72 h | [105] |
| Pesticide | Species/Strain | Acting Enzyme | Gene | Protein | Features | Reference |
|---|---|---|---|---|---|---|
| Chlorpyrifos, methylparathion, parathion, coumaphos, diazinon | Lev. brevis WCP902 | Organophosphate hydrolase | opdB 723 bp | 240 AA 27 kDa | ‘Gly-X-Ser-X-Gly’-motif; Ser82 * pH 6.0, 35 °C | [93] |
| La. sakei WCP904 | Organophosphate hydrolase | opdD 825 bp | 274 AA 31 kDa | ‘Gly-X-Ser-X-Gly’-motif; Ser 116 * pH 6, 30 °C | [123] | |
| Leuc. mesenteroides WCP307 | Organophosphate hydrolase | opdA 930 bp | 309 AA 35 kDa | ‘Gly-X-Ser-X-Gly’-motif; Ser128 *; pH 7, 30 °C | [124] | |
| Leuc. mesenteroides WCP307 | Organophosphate hydrolase | opdE 894 bp | 297 AA 33 kDa | ‘Gly-X-Ser-X-Gly’ -motif, Ser129 *; pH 6, 30 °C | [124] | |
| Lp. plantarum WCP931, Leuc. mesenteroides WCP907, Lev. brevis WCP902, La. sakei WCP904 | Esterase (suggested) | - | - | - | [92] | |
| Parathion, methylparathion, chlorpyrifos | Lp. plantarum WCP931 | Organophosphate hydrolase | opdC 831 bp | 276 AA 31 kDa | ‘Gly-X-Ser-X-Gly’—motif; Ser 116 *; pH 6, 35 °C | [125] |
| Chlorpyrifos | Lc. casei WYS3 | Organophosphate hydrolase | opda | - | (+) opda RNA levels; hydrolysis products detected by GC-MS | [126] |
| Dimethoate | Lc. casei 355 | Alkaline phosphatase | - | 43 kDa | pH 8.5, 37 °C (+) Mg2+, Ca2+ | [127] |
| (purified) | (−) Cu2+, Zn2+, EDTA | |||||
| Lp. plantarum CICC20261 | Phosphatase (crude activity) | - | - | Gln375 and Ser415 * | [87] | |
| Dimethoate, chlorpyrifos, trichlorphon, methylparathion | Lp. plantarum subsp. plantarum CICC20261 | Phosphatase (crude activity) | - | - | - | [128] |
| Trichlorphon, phorate, malathion, dichlorvos | L. delbrueckii subsp. bulgaricus, S. thermophilus, Lp. rhamnosus | Phosphatase activity (correlation) | - | - | - | [88] |
| Chlorpyrifos, fenitrothion, malathion | Lev. brevis 1.0209 | Phosphatase activity (correlation) | - | - | - | [89] |
| Chlorpyrifos | Pediococcus pentasaceus 4320, Leuc. mesenteroides 8293, Ent. faecium 86, Lactococcus. lactis 1454, Lc. rhamnosus GG53103, Leuc. lactis 19256 | Alkaline phosphatase (suggested) | - | - | - | [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armenova, N.; Tsigoriyna, L.; Arsov, A.; Petrov, K.; Petrova, P. Microbial Detoxification of Residual Pesticides in Fermented Foods: Current Status and Prospects. Foods 2023, 12, 1163. https://doi.org/10.3390/foods12061163
Armenova N, Tsigoriyna L, Arsov A, Petrov K, Petrova P. Microbial Detoxification of Residual Pesticides in Fermented Foods: Current Status and Prospects. Foods. 2023; 12(6):1163. https://doi.org/10.3390/foods12061163
Chicago/Turabian StyleArmenova, Nadya, Lidia Tsigoriyna, Alexander Arsov, Kaloyan Petrov, and Penka Petrova. 2023. "Microbial Detoxification of Residual Pesticides in Fermented Foods: Current Status and Prospects" Foods 12, no. 6: 1163. https://doi.org/10.3390/foods12061163
APA StyleArmenova, N., Tsigoriyna, L., Arsov, A., Petrov, K., & Petrova, P. (2023). Microbial Detoxification of Residual Pesticides in Fermented Foods: Current Status and Prospects. Foods, 12(6), 1163. https://doi.org/10.3390/foods12061163