

Effect of 2,5-Dicarbonyl-3-Isobutyl-Piperazine on 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Wine Grape

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Solutions
2.2. Incorporation Treatments on Clusters In Situ
2.2.1. Vineyard Site
2.2.2. Experiment Treatments
2.2.3. Sample Collection and Pretreatment
2.3. IBMP Quantitation
2.4. RNA Extractions, cDNA Synthesis, and Real-Time Polymerase Chain Reaction
2.5. Statistics
3. Results
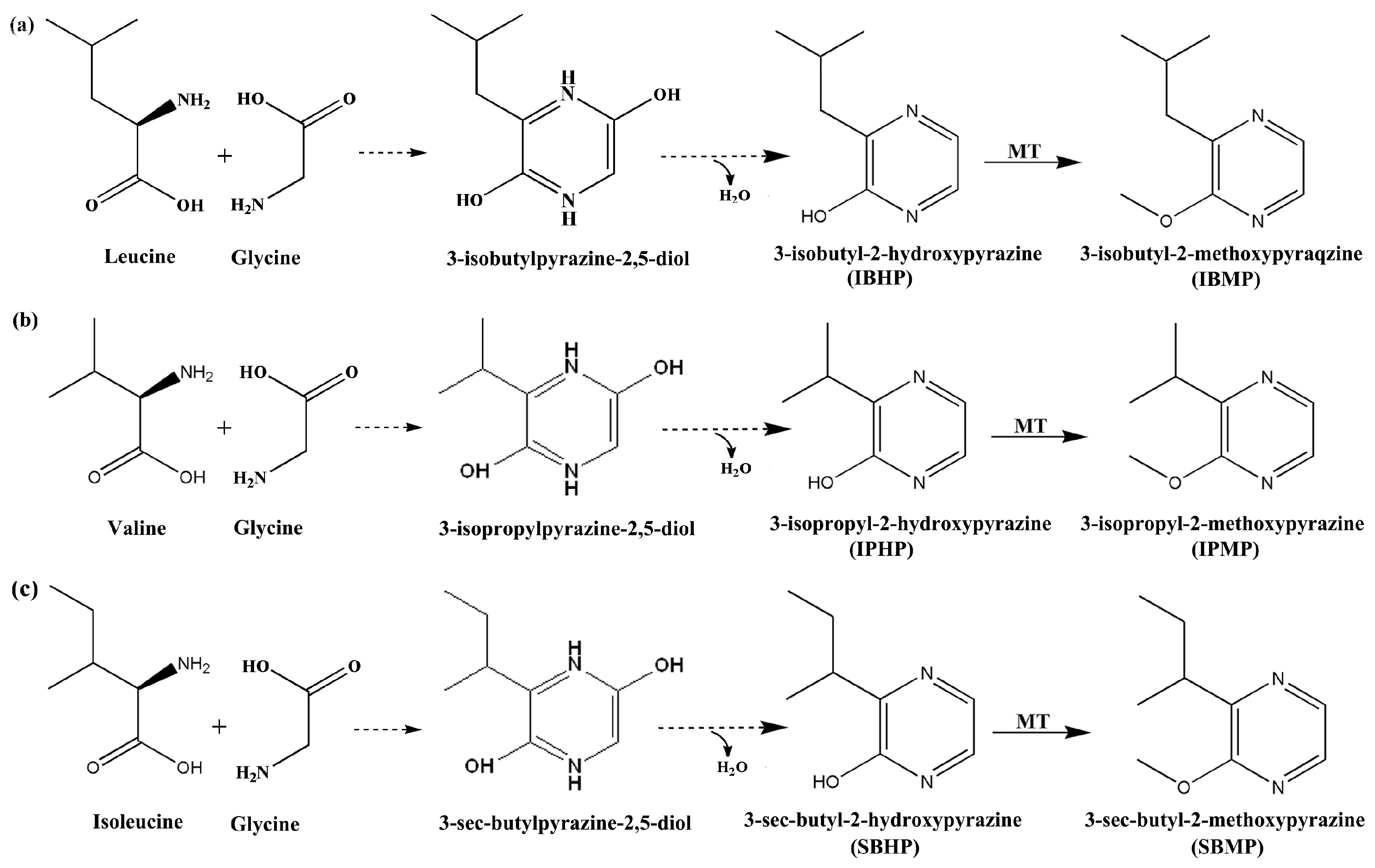
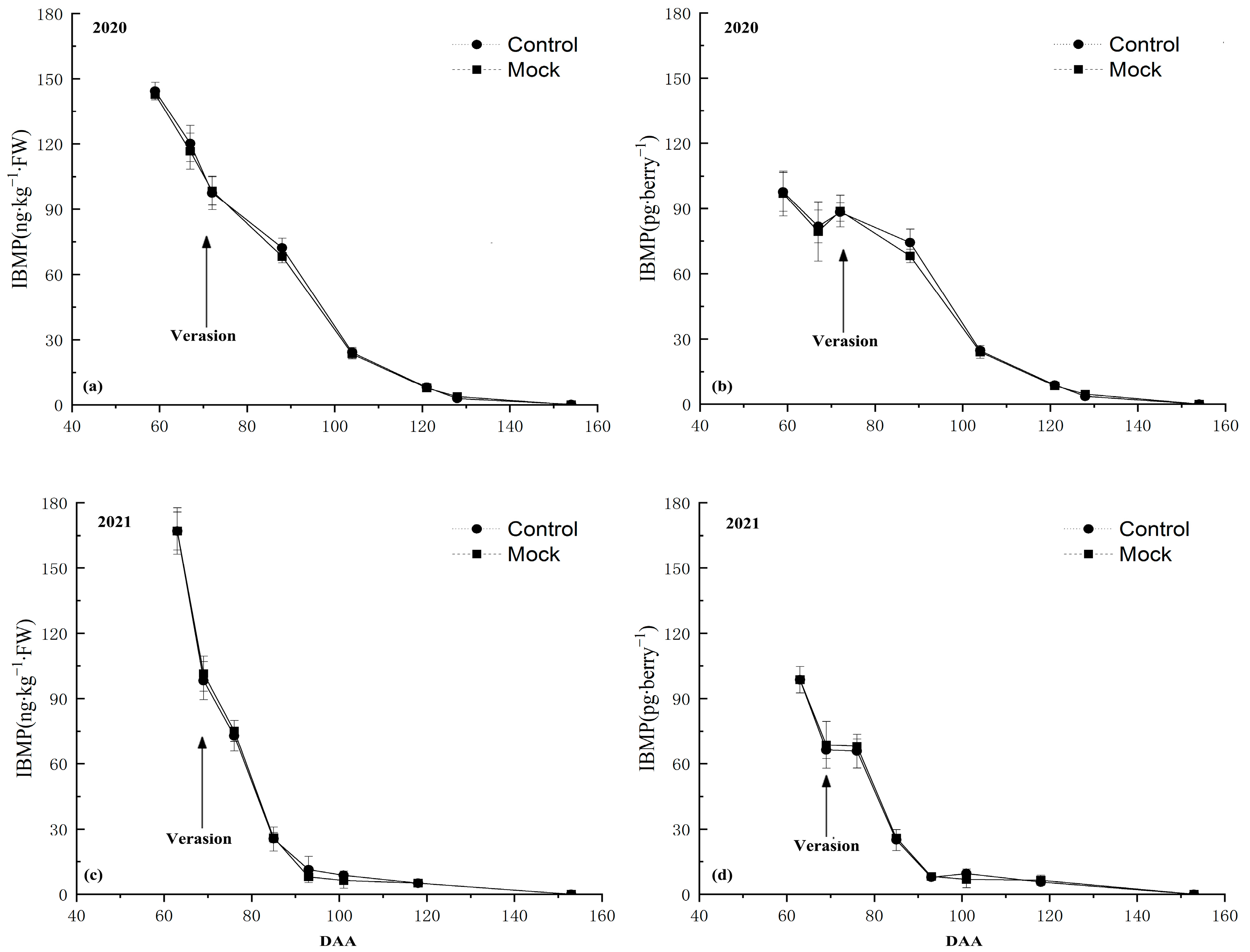
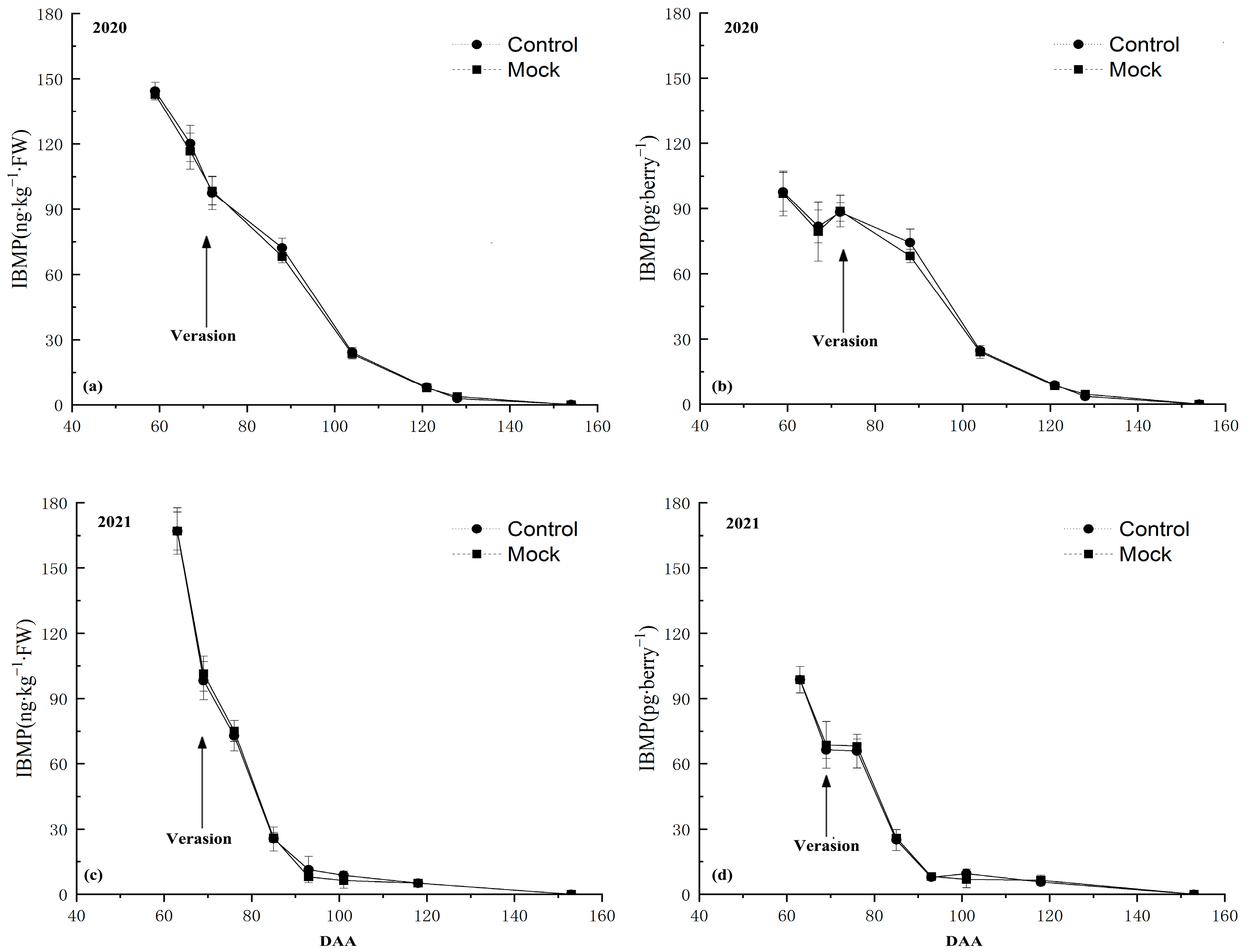
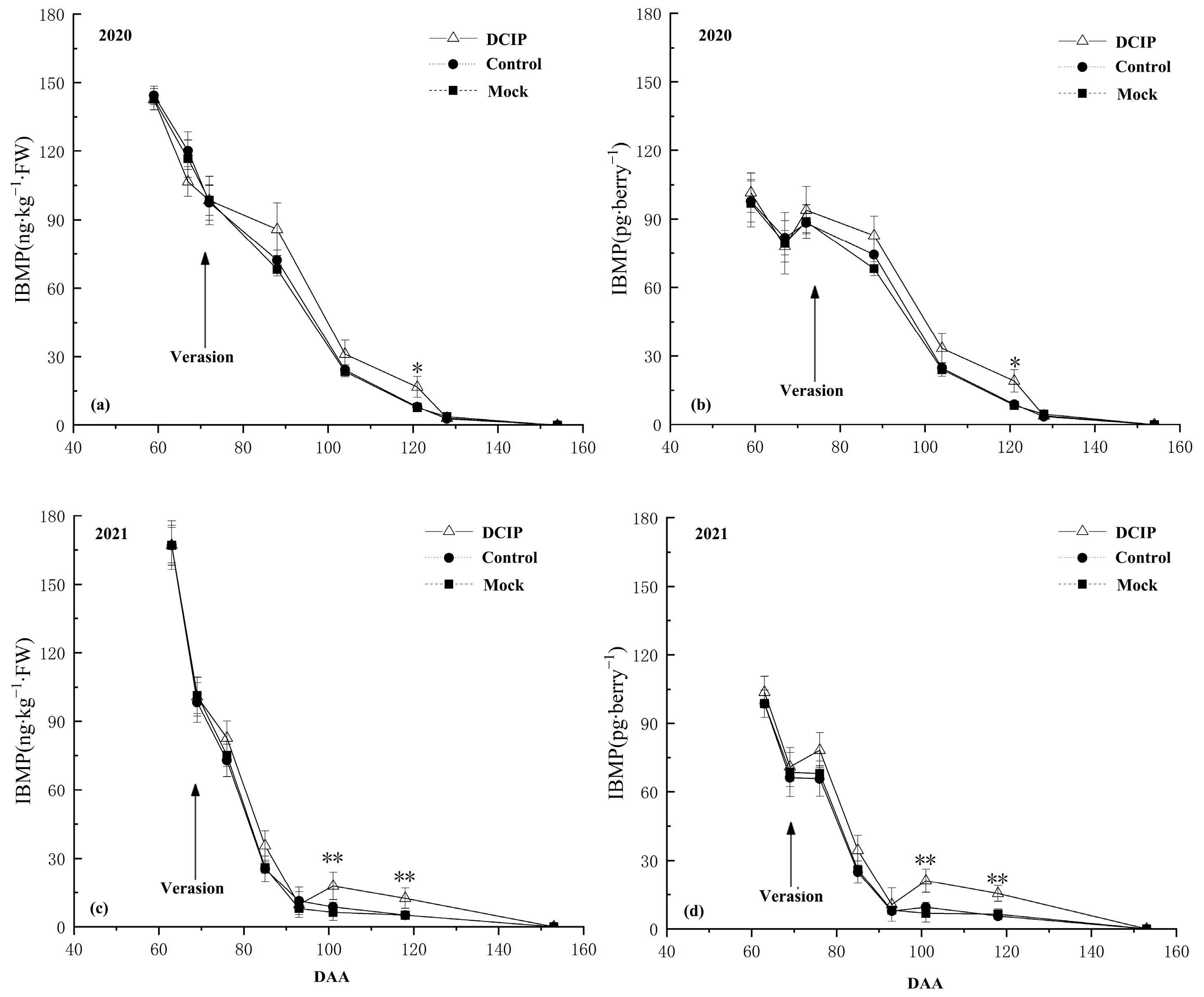
3.1. IBMP Synthesis and Accumulation in Grape Berry
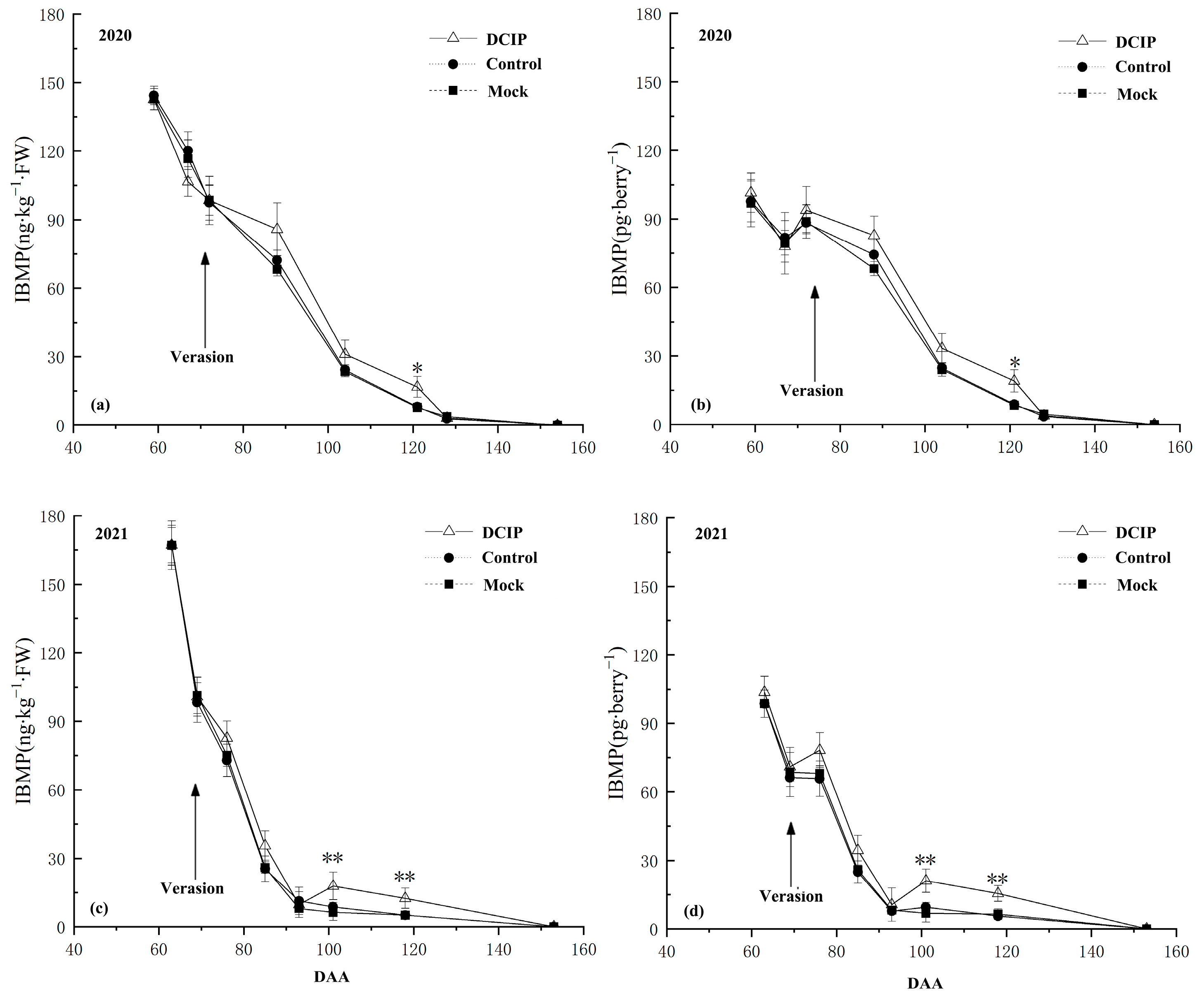
3.2. IBMP Accumulation in Grapes after DCIP Incorporation
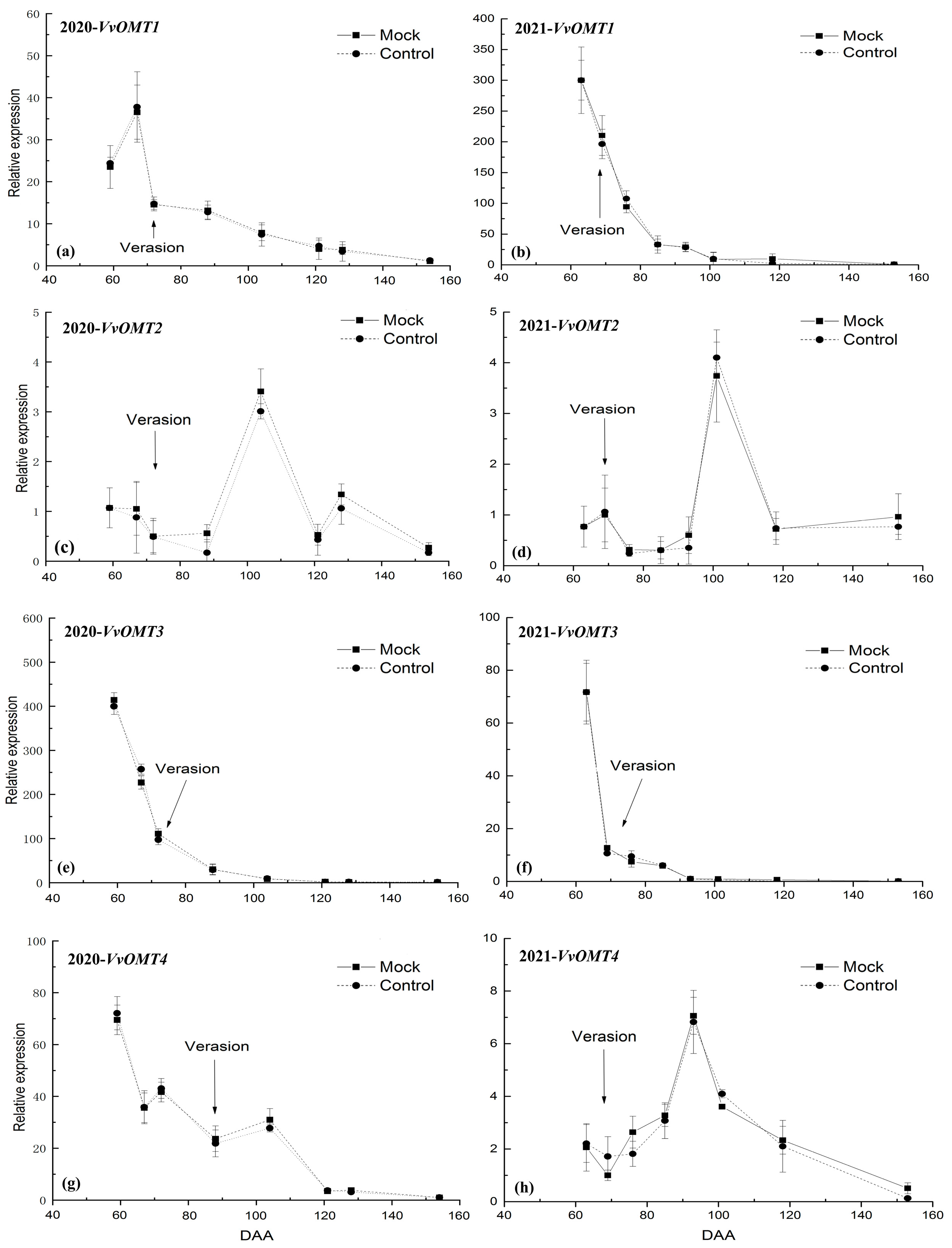
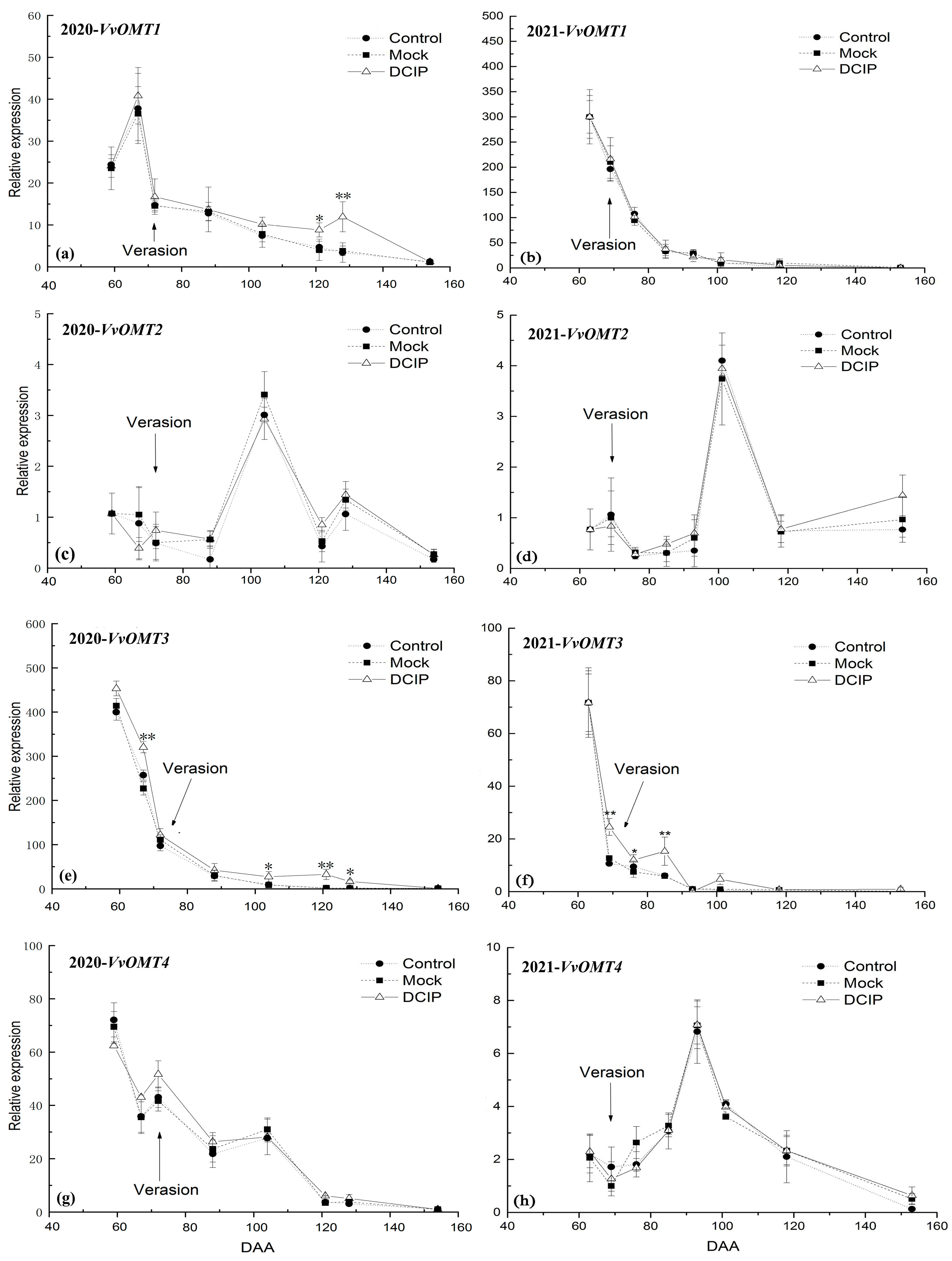
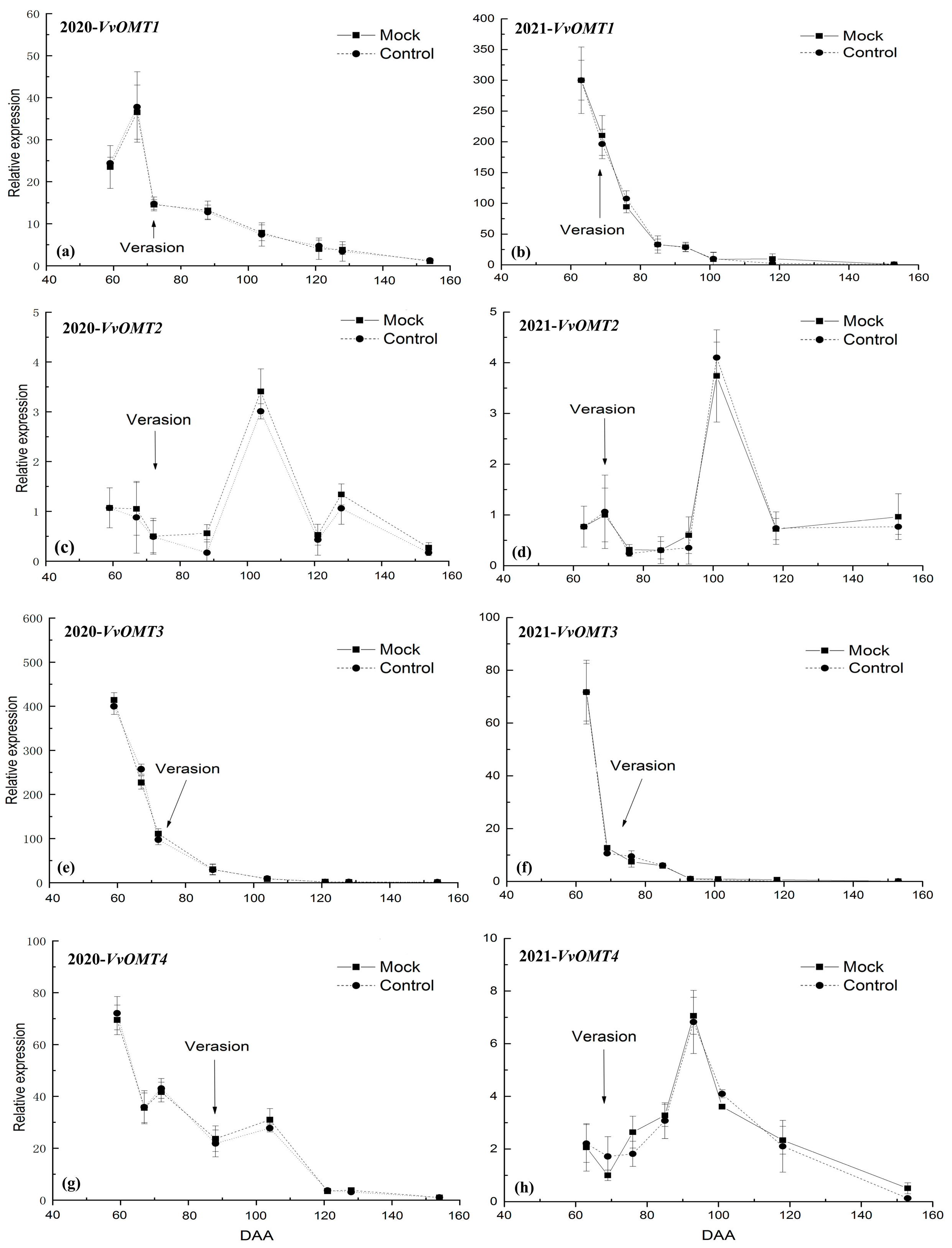
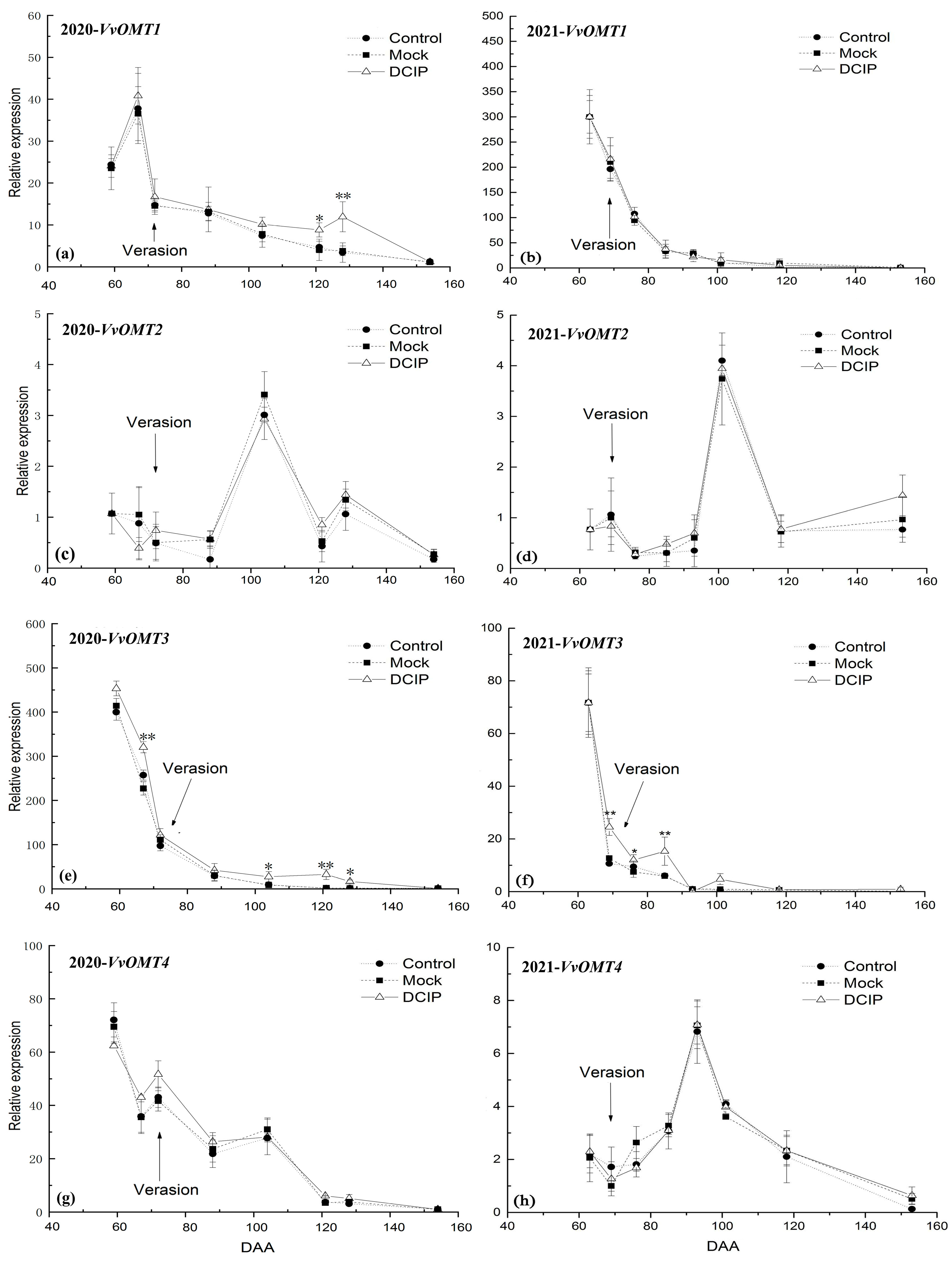
3.3. The Relative Expression Level of VvOMTs in Berry Skins after DCIP Incorporation
4. Discussion
4.1. The Relationship between the DCIP and IBMP Accumulation
4.2. Correlation of the Relative Expression Level of VvOMTs in Berry Skins with IBMP Biosynthesis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Leeuwen, C.; Roby, J.P.; Alonso-Villaverde, V.; Gindro, K. Impact of clonal variability in Vitis vinifera Cabernet franc on Grape composition, wine quality, leaf blade stilbene content, and downy mildew resistance. J. Agric. Food Chem. 2013, 61, 19–24. [Google Scholar] [CrossRef]
- Ward, S.C.; Petrie, P.R.; Johnson, T.E.; Boss, P.K.; Bastian, S.E.P. Unripe berries and petioles in Vitis vinifera cv. Cabernet Sauvignon fermentations affect sensory and chemical profiles. Am. J. Enol. Vitic. 2015, 66, 435–443. [Google Scholar] [CrossRef]
- Mozzon, M.; Savini, S.; Boselli, E.; Thorngate, J.H. The herbaceous character of wines. Ital. J. Food Sci. 2016, 28, 190–207. [Google Scholar]
- Lei, Y.J.; Xie, S.; Guan, X.Q.; Song, C.Z.; Zhang, Z.W.; Meng, J.F. Methoxypyrazines biosynthesis and metabolism in grape: A review. Food Chem. 2018, 245, 1141–1147. [Google Scholar] [CrossRef]
- Allen, M.S.; Lacey, M.J.; Brown, W.V.; Harris, R.L.N. Occurrence of methoxypyrazine in grapes of Vitis vinifera cv. Cabernet-sauvignon and sauvignon blanc. In Actualites Enologiques 89, Compte Rendu du IVe Symposium d’ Enologie de Bordeaux; Dunod: Paris, France, 1989; pp. 25–30. [Google Scholar]
- Kotseridis, Y.; Anocibar, B.A.; Bertrand, A.; Doazan, J.P. An analytical method for studying the volatile compounds of merlot noir clone wines. Am. J. Enol. Vitic. 1998, 49, 44–47. [Google Scholar] [CrossRef]
- Roujou-de-Boubée, D.; Van Leeuwen, C.; Dubordieu, D. Organoleptic impact of 2-methoxy-3-isobutylpyrazine on red Bordeaux and Loire wines. Effect of environmental conditions on concentrations in grapes during ripening. J. Agric. Food Chem. 2000, 48, 4830–4835. [Google Scholar] [CrossRef]
- Maga, J.A. Sensory and stability properties of added methoxypyrazines to model and authentic wine. Dev. Food Sci. 1990, 24, 61–70. [Google Scholar]
- Allen, M.S.; Lacey, M.J.; Boyd, S. Determination of methoxypyrazines in red wines by stable isotope dilution gas chromatography-mass spectrometry. J. Agric. Food Chem. 1994, 42, 1734–1738. [Google Scholar] [CrossRef]
- Lacey, M.J.; Allen, M.S.; Harris, R.L.N.; Brown, W.V. Methoxypyrazines in Sauvignon blanc grapes and wines. Am. J. Enol. Vitic. 1991, 42, 103–108. [Google Scholar] [CrossRef]
- Harris, R.L.N.; Lacey, M.J.; Brown, W.V.; Allen, M.S. Determination of 2-methoxy-3-alkylpyrazines in wine by gas chromatography/mass spectrometry. Vitis 1987, 26, 201–207. [Google Scholar]
- Chapman, D.M.; Thorngate, J.H.; Matthews, M.A.; Guinard, J.X.; Ebeler, S.E. Yield effects on 2-methoxy-3-isobutylpyrazine concentration in cabernet sauvignon using a solid phase micro-extraction gas chromatography/mass spectrometry method. J. Agric. Food Chem. 2004, 52, 5431–5435. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, D.; Lund, J.; Kotseridis, Y.; Saucier, C. Methoxypyrazine analysis and influence of viticultural and enological procedures on their levels in grapes, musts, and wines. Crit. Rev. Food Sci. Nutr. 2015, 55, 485–502. [Google Scholar] [CrossRef]
- Botezatu, A.; Kemp, B.S.; Pickering, G.J. Chemical and sensory evaluation of silicone and polylactic acid-based remedial treatments for elevated methoxypyrazine levels in wine. Molecules 2016, 21, 1238. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.L.; Sacks, G.L. Rapid headspace solid-phase microextraction sheets with direct analysis in real time mass spectrometry (SPMESH-DART-MS) of derivatized volatile phenols in grape juices and wines. Anal. Chim. Acta 2023, 1275, 341577. [Google Scholar] [CrossRef]
- Jastrzembski, J.A.; Sacks, G.L. Solid phase mesh enhanced sorption from headspace (SPMESH) coupled to DART-MS for rapid quantification of trace-level volatiles. Anal. Chem. 2016, 88, 8617–8623. [Google Scholar] [CrossRef]
- de Villiers, A.; Alberts, P.; Tredoux, A.G.; Nieuwoudt, H.H. Analytical techniques for wine analysis: An African perspective; a review. Anal. Chim. Acta 2012, 730, 2–23. [Google Scholar] [CrossRef]
- Nolvachai, Y.; Amaral, M.S.; Marriott, P.J. Foods and contaminants analysis using multidimensional gas chromatography: An update of recent studies, technology, and applications. Anal. Chem. 2023, 95, 238–263. [Google Scholar] [CrossRef]
- Hashizume, K.; Umeda, N. Methoxypyrazine content of Japanese red wines. Biosci. Biotechnol. Biochem. 1996, 60, 802–805. [Google Scholar] [CrossRef]
- Belancic, A.; Agosin, E. Methoxypyrazines in grapes and wines of Vitis vinifera cv. Carmenere. Am. J. Enol. Vitic. 2007, 58, 462–469. [Google Scholar] [CrossRef]
- Roujou-de-Boubée, D.; Cumsille, A.M.; Pons, D.; Dubordieu, D. Location of 2-methoxy-3-isobutylpyrazine in Cabernet sauvignon bunches and its extractability during vinification. Am. J. Enol. Vitic. 2002, 53, 1–5. [Google Scholar] [CrossRef]
- Murray, K.E.; Shipton, J.; Whitfield, F.B. 2-Methoxypyrazines and the flavor of green peas (Pisum sativum). Chem. Ind. 1970, 7, 897–898. [Google Scholar]
- Cheng, T.B.; Reineccius, G.A.; Bjorklund, J.A.; Leete, E. Biosynthesis of 2-methoxy-3-isopropylpyrazine in Pseudomonas perolens. J. Agric. Food Chem. 1991, 39, 1009–1012. [Google Scholar] [CrossRef]
- Hashizume, K.; Tozawa, K.; Endo, M.; Aramaki, I. S-AdenosylL-methionine-dependent O-methylation of 2-Hydroxy-3-alkylpyrazine in Wine Grapes: A putative final step of methoxypyrazine biosynthesis. Biosci. Biotechnol. Biochem. 2001, 65, 795–801. [Google Scholar] [CrossRef]
- Hashizume, K.; Tozawa, K.; Hiraga, Y.; Aramaki, I. Purification and characterization of a O-methyltransferase capable of methylating 2-hydroxy-3-alkylpyrazine from Vitis vinifera L. (cv. Cabernet Sauvignon). Biosci. Biotechnol. Biochem. 2001, 65, 2213–2219. [Google Scholar] [CrossRef]
- Ryona, I.; Leclerc, S.; Sacks, G.L. Correlation of 3-Isobutyl-2-methoxypyrazine to 3-Isobutyl-2-hydroxypyrazine during Maturation of Bell Pepper (Capsicum annuum) and Wine Grapes (Vitis vinifera). J. Agric. Food Chem. 2010, 58, 9723–9730. [Google Scholar] [CrossRef]
- Dunlevy, J.D.; Soole, K.L.; Perkins, V.M.; Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Boss, P.K. Two O-methyltransferases involved in the biosynthesis of methoxypyrazines: Grape-derived aroma compounds important to wine flavor. Plant Mol. Biol. 2010, 74, 77–89. [Google Scholar] [CrossRef]
- Vallarino, J.G.; Lopez-Cortes, X.A.; Dunlevy, J.D.; Boss, P.K.; Gonzalez-Nilo, F.D.; Moreno, Y.M. Biosynthesis of methoxypyrazines: Elucidating the structural/functional relationship of two Vitisvinifera O-methyltransferases capable of catalyzing the putative final step of the biosynthesis of 3-alkyl-2-methoxypyrazine. J. Agric. Food Chem. 2011, 59, 7310–7316. [Google Scholar] [CrossRef]
- Dunlevy, J.D.; Dennis, E.G.; Soole, K.L.; Perkins, M.V.; Davies, C.; Boss, P.K. A methyltransferase essential for the methoxypyrazine-derived flavour of wine. Plant J. 2013, 75, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Heymann, H.; Noble, A.C.; Boulton, R.B. Analysis of methoxypyrazines in wines. 1. Development of a quantitative procedure. J. Agric. Food Chem. 1986, 34, 268–271. [Google Scholar] [CrossRef]
- Allen, M.S.; Lacey, M.J.; Harris, R.L.N.; Brown, W.V. Contribution of methoxypyrazines to Sauvignon blanc wine aroma. Am. J. Enol. Vitic. 1991, 42, 109–112. [Google Scholar] [CrossRef]
- Harris, S.A.; Ryona, I.; Sacks, G.L. Behavior of 3-Isobutyl-2-hydroxypyrazine (IBHP), a Key Intermediate in 3-Isobutyl-2-methoxypyrazine (IBMP) Metabolism, in Ripening Wine Grapes. J. Agric. Food Chem. 2012, 60, 11901–11908. [Google Scholar] [CrossRef]
- Hayasaka, Y.; Baldock, G.A.; Pardon, K.H.; Jeffery, D.W.; Herderich, M.J. Investigation into the Formation of Guaiacol Conjugates in Berries and Leaves of Grapevine Vitis vinifera L. cv. Cabernet Sauvignon Using Stable Isotope Tracers Combined with HPLC-MS and MS/MS Analysis. J. Agric. Food Chem. 2010, 58, 2076–2081. [Google Scholar] [CrossRef] [PubMed]
- Chassy, A.W.; Adams, D.O.; Laurie, V.F.; Waterhouse, A.L. Tracing phenolic biosynthesis in Vitis vinifera via in situ C-13 labeling and liquid chromatography diode-array detector-mass spectrometer/mass spectrometer detection. Anal. Chim. Acta 2012, 747, 51–57. [Google Scholar] [CrossRef]
- Lei, Y.J.; Xie, S.; Chen, H.Z.; Guan, X.Q.; Zhang, Z.W. Behavior of 3-isobutyl-2-methoxypyrazine biosynthesis related to proposed precursor and intermediate in wine grape. Food Chem. 2019, 217, 609–616. [Google Scholar] [CrossRef]
- Koch, A.; Ebeler, S.E.; Williams, L.E.; Matthews, M.A. Fruit ripening in Vitis vinifera: Light intensity before and not during ripening determines the concentration of 2-methoxy-3-isobutylpyrazine in Cabernet Sauvignon berries. Physiol. Plant. 2012, 145, 275–285. [Google Scholar] [CrossRef]
- Hashizume, K.; Samuta, T. Grape maturity and light exposure affect berry methoxypyrazine concentration. Am. J. Enol. Vitic. 1999, 50, 194–198. [Google Scholar] [CrossRef]
- Gregan, S.M.; Wargent, J.J.; Liu, L.; Shinkle, J.; Hofmann, R.; Winefield, C.; Trought, M.; Jordan, B. Effects of solar ultraviolet radiation and canopy manipulation on the biochemical composition of Sauvignon Blanc grapes. Aust. J. Grape Wine Res. 2012, 18, 227–238. [Google Scholar] [CrossRef]
- Scheiner, J.J.; Sacks, G.L.; Pan, B.; Ennahli, S.; Tarlton, L.; Wise, A.; Lerch, S.D.; Vanden Heuvel, J.E. Impact of severity and timing of basal leaf removal on 3-isobutyl-2-methoxypyrazine concentrations in red wine grapes. Am. J. Enol. Vitic. 2010, 61, 358–364. [Google Scholar] [CrossRef]
- Gregan, S.M.; Jordan, B. Methoxypyrazine accumulation and O-methyltransferase gene expression in Sauvignon blanc grapes: The role of leaf removal, light exposure, and berry development. J. Agric. Food Chem. 2016, 64, 2200–2208. [Google Scholar] [CrossRef]
- Guillaumie, S.; Ilg, A.; Rety, S.; Brette, M.; Trossat-Magnin, C.; Decroocq, S. Genetic analysis of the biosynthesis of 2-methoxy-3-isobutilpyrazine, a major grape derived aroma compound impacting wine quality. Plant Physiol. 2013, 162, 604–615. [Google Scholar] [CrossRef]
- Roujou-de-Boubée, D.; Botella, T. Research on 2-Methoxy-3-Isobutylpyrazine in Grapes and Wines; Academie Amorim: Bordeaux, France, 2003. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Y.; Ma, Z.; Wang, P.; Qin, X.; Guan, X.; Zhang, Z. Effect of 2,5-Dicarbonyl-3-Isobutyl-Piperazine on 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Wine Grape. Foods 2023, 12, 3258. https://doi.org/10.3390/foods12173258
Lei Y, Ma Z, Wang P, Qin X, Guan X, Zhang Z. Effect of 2,5-Dicarbonyl-3-Isobutyl-Piperazine on 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Wine Grape. Foods. 2023; 12(17):3258. https://doi.org/10.3390/foods12173258
Chicago/Turabian StyleLei, Yujuan, Zhansheng Ma, Ping Wang, Xuchen Qin, Xueqiang Guan, and Zhenwen Zhang. 2023. "Effect of 2,5-Dicarbonyl-3-Isobutyl-Piperazine on 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Wine Grape" Foods 12, no. 17: 3258. https://doi.org/10.3390/foods12173258
APA StyleLei, Y., Ma, Z., Wang, P., Qin, X., Guan, X., & Zhang, Z. (2023). Effect of 2,5-Dicarbonyl-3-Isobutyl-Piperazine on 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Wine Grape. Foods, 12(17), 3258. https://doi.org/10.3390/foods12173258