

Effects of Cow’s Milk Processing on MicroRNA Levels
 ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk Sample Collection and Treatments
2.2. Total RNA Extraction and Spike in
2.3. Real Time–Quantitative PCR (RT-qPCR)
2.4. Prediction of miRNA Structure
2.5. Statistical Analyses
3. Results
3.1. Validation of Milk Treatments
3.2. miRNA Losses Due to Milk Processing
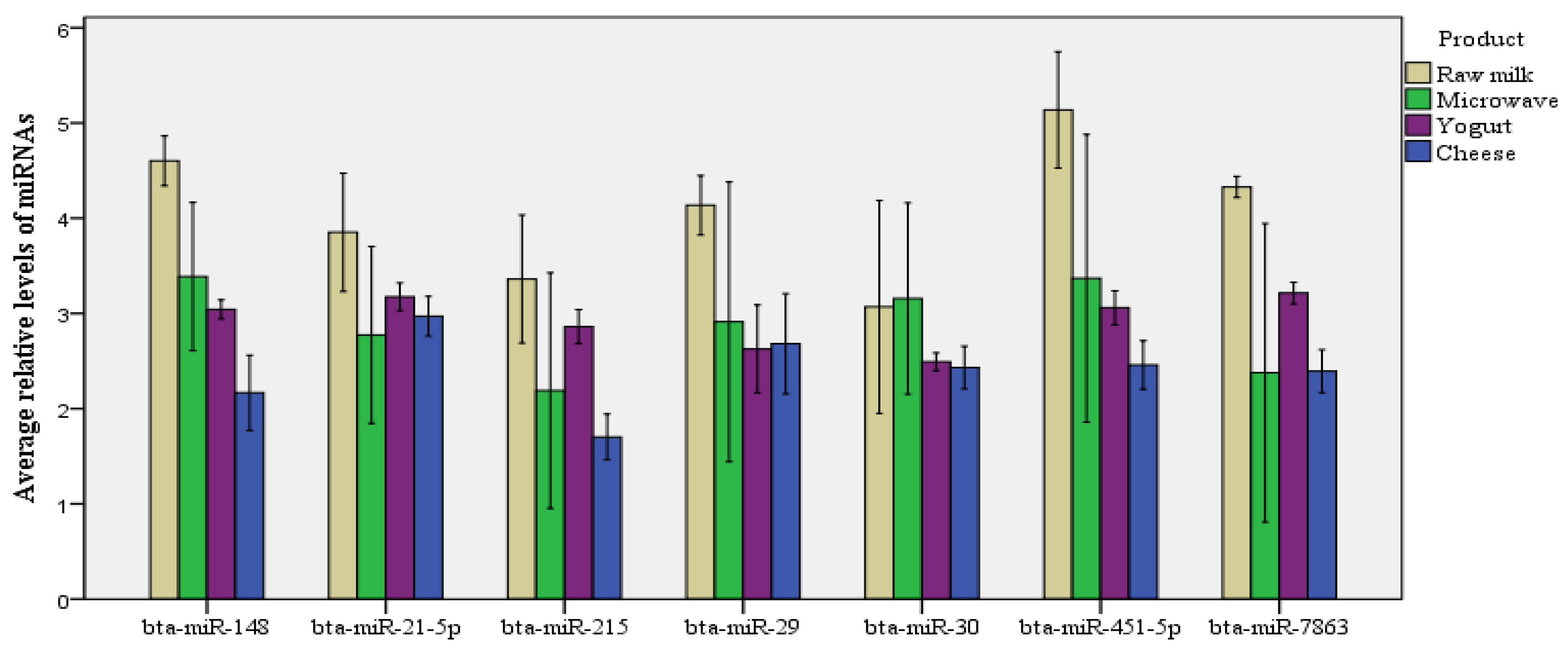
3.3. miRNA Concentrations in Milk and Cheese
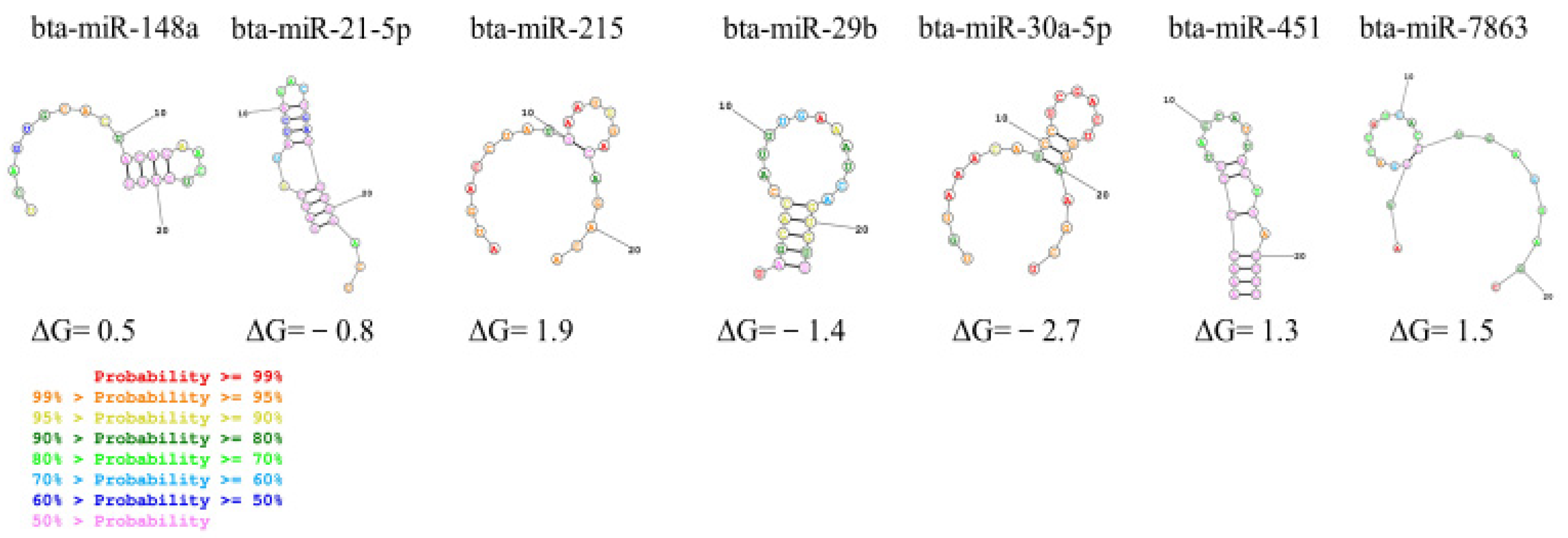
3.4. Determination of Secondary Structure of miRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, L.; Hannon, G.J. MicroRNAs: Small rnas with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microrna function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.Y. Horizontal transfer of micrornas: Molecular mechanisms and clinical applications. Protein Cell 2012, 3, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Lozano, E.; Sebastián-Valles, F.; Knott-Torcal, C. Circulating microRNAs in breast milk and their potential impact on the infant. Nutrients 2020, 12, 3066. [Google Scholar] [CrossRef]
- Alsaweed, M.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. Human Milk MiRNAs Primarily originate from the mammary gland resulting in unique mirna profiles of fractionated milk. Sci. Rep. 2016, 6, 20680. [Google Scholar] [CrossRef]
- Hata, T.; Murakami, K.; Nakatani, H.; Yamamoto, Y.; Matsuda, T.; Aoki, N. Isolation of bovine milk-derived microvesicles carrying mRNAs and microRNAs. Biochem. Biophys. Res. Commun. 2010, 396, 528–533. [Google Scholar] [CrossRef]
- Munch, E.M.; Harris, R.A.; Mohammad, M.; Benham, A.L.; Pejerrey, S.M.; Showalter, L.; Hu, M.; Shope, C.D.; Maningat, P.D.; Gunaratne, P.H.; et al. Transcriptome profiling of microrna by next-gen deep sequencing reveals known and novel miRNA species in the lipid fraction of human breast milk. PLoS ONE 2013, 8, e50564. [Google Scholar] [CrossRef]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Time-dependent expression profiles of microRNAs and mRNAs in rat milk Whey. PLoS ONE 2014, 9, e88843. [Google Scholar] [CrossRef]
- Wolf, T.; Baier, S.R.; Zempleni, J. The Intestinal Transport of Bovine Milk Exosomes is mediated by endocytosis in human colon carcinoma caco-2 cells and rat small intestinal iec-6 cells1-3. J. Nutr. 2015, 145, 2201–2206. [Google Scholar] [CrossRef]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, hek-293 kidney cell cultures, and mouse livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- López de Las Hazas, M.C.; Del Pozo-Acebo, L.; Hansen, M.S.; Gil-Zamorano, J.; Mantilla-Escalante, D.C.; Gómez-Coronado, D.; Marín, F.; Garcia-Ruiz, A.; Rasmussen, J.T.; Dávalos, A. Dietary Bovine Milk MiRNAs transported in extracellular vesicles are partially stable during gi digestion, are bioavailable and reach target tissues but need a minimum dose to impact on gene expression. Eur. J. Nutr. 2022, 61, 1043–1056. [Google Scholar] [CrossRef]
- Zempleni, J.; Baier, S.R.; Howard, K.M.; Cui, J. Gene regulation by dietary microRNAs. Can. J. Physiol. Pharmacol. 2015, 93, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo-Acebo, L.; de las Hazas, M.C.L.; Tomé-Carneiro, J.; Gil-Cabrerizo, P.; San-Cristobal, R.; Busto, R.; García-Ruiz, A.; Dávalos, A. Bovine milk-derived exosomes as a drug delivery vehicle for mirna-based therapy. Int. J. Mol. Sci. 2021, 22, 1105. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Fernández-Alonso, N.; Tomás-Zapico, C.; Visioli, F.; Iglesias-Gutierrez, E.; Dávalos, A. Breast milk microRNASs harsh journey towards potential effects in infant development and maturation. Lipid encapsulation can help. Pharmacol. Res. 2018, 132, 21–32. [Google Scholar] [CrossRef]
- Cammaerts, S.; Strazisar, M.; De Rijk, P.; Del Favero, J. Genetic variants in microRNA genes: Impact on microRNA expression, function, and disease. Front. Genet. 2015, 6, 186. [Google Scholar] [CrossRef]
- Colitti, M.; Sgorlon, S.; Licastro, D.; Stefanon, B. Differential expression of miRNAs in milk exosomes of cows subjected to group relocation. Res. Vet. Sci. 2019, 122, 148–155. [Google Scholar] [CrossRef]
- Li, R.; Beaudoin, F.; Ammah, A.A.; Bissonnette, N.; Benchaar, C.; Zhao, X.; Lei, C.; Ibeagha-Awemu, E.M. Deep sequencing shows microRNA involvement in bovine mammary gland adaptation to diets supplemented with linseed oil or safflower oil. BMC Genom. 2015, 16, 884. [Google Scholar] [CrossRef]
- Muroya, S.; Shibata, M.; Hayashi, M.; Oe, M.; Ojima, K. Differences in circulating microRNAs between grazing and grain-fed wagyu cattle are associated with altered expression of intramuscular microRNA, the potential target PTEN, and lipogenic genes. PLoS ONE 2016, 11, e0162496. [Google Scholar] [CrossRef]
- Alsaweed, M.; Hepworth, A.R.; Lefèvre, C.; Hartmann, P.E.; Geddes, D.T.; Hassiotou, F. Human Milk microRNA and total RNA differ depending on milk fractionation. J. Cell. Biochem. 2015, 116, 2397–2407. [Google Scholar] [CrossRef]
- Li, R.; Dudemaine, P.L.; Zhao, X.; Lei, C.; Ibeagha-Awemu, E.M. Comparative analysis of the miRNome of bovine milk fat, whey and cells. PLoS ONE 2016, 11, e0154129. [Google Scholar] [CrossRef]
- Abou El Qassim, L.; Le Guillou, S.; Royo, L.J. Variation of miRNA content in cow raw milk depending on the dairy production system. Int. J. Mol. Sci. 2022, 23, 11681. [Google Scholar] [CrossRef] [PubMed]
- Abou El Qassim, L.; Alonso, J.; Zhao, K.; Guillou, S.L.; Diez, J.; Vicente, F. Differences in the microRNAs levels of raw milk from dairy cattle raised under extensive or intensive production systems. Vet. Sci. 2022, 9, 661. [Google Scholar] [CrossRef]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J. Dairy Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef]
- Abou El Qassim, L.; Royo, L.J. The effect of pasteurization in the expression of bovine milk microRNA. In Book of Abstracts of the 72nd Annual Meeting of the European Federation of Animal Science, Davos, Switzerland, 30 August–3 September 2021; European Federation of Animal Science: Rome, Italy, 2021; Volume 27, p. 637. [Google Scholar]
- Buschmann, D.; Haberberger, A.; Kirchner, B.; Spornraft, M.; Riedmaier, I.; Schelling, G.; Pfaffl, M.W. Toward reliable biomarker signatures in the age of liquid biopsies—How to standardize the small RNA-seq workflow. Nucleic Acids Res. 2016, 44, 5995–6018. [Google Scholar] [CrossRef] [PubMed]
- Hynes, E.; Ogier, J.C.; Delacroix-Buchet, A. Protocol for the Manufacture of miniature washed-curd cheeses under controlled microbiological conditions. Int. Dairy J. 2000, 10, 733–737. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2008, 8, R19. [Google Scholar] [CrossRef]
- Rani, P.; Yenuganti, V.R.; Shandilya, S.; Onteru, S.K.; Singh, D. MiRNAs: The hidden bioactive component of milk. Trends Food Sci. Technol. 2017, 65, 94–102. [Google Scholar] [CrossRef]
- Musto, M.; Faraone, D.; Cellini, F.; Musto, E. Changes of DNA quality and meat physicochemical properties in bovine supraspinatus muscle during microwave heating. J. Sci. Food Agric. 2014, 94, 785–791. [Google Scholar] [CrossRef]
- Dehghan, A.; Jamalian, J.; Farahnaky, A.; Mesbahi, G.; Moosavi-Nasab, M. The effect of microwave pasteurization on some physical and chemical characteristics of milk. Int. J. Food Eng. 2012, 8, 1–12. [Google Scholar] [CrossRef]
- Zhao, Z.; Yu, S.; Xu, M.; Li, P. Effects of microwave on extracellular vesicles and microRNA in milk. J. Dairy Sci. 2018, 101, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Howard, K.M.; Jati Kusuma, R.; Baier, S.R.; Friemel, T.; Markham, L.; Vanamala, J.; Zempleni, J. Loss of miRNAs during processing and storage of cow’s (Bos taurus) Milk. J. Agric. Food Chem. 2015, 63, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Smyczynska, U.; Bartlomiejczyk, M.A.; Stanczak, M.M.; Sztromwasser, P.; Wesolowska, A.; Barbarska, O.; Pawlikowska, E.; Fendler, W. Impact of processing method on donated human breast milk microRNA content. PLoS ONE 2020, 15, e0236126. [Google Scholar] [CrossRef]
- Sohel, M.H. Extracellular/Circulating microRNAs: Release mechanisms, functions and challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef]
- Le, T.T.; Van de Wiele, T.; Do, T.N.H.; Debyser, G.; Struijs, K.; Devreese, B.; Dewettinck, K.; Van Camp, J. Stability of milk fat globule membrane proteins toward human enzymatic gastrointestinal digestion. J. Dairy Sci. 2012, 95, 2307–2318. [Google Scholar] [CrossRef]
- Rodríguez-Alcalá, L.M.; Alonso, L.; Fontecha, J. Stability of fatty acid composition after thermal, high pressure, and microwave processing of cow milk as affected by polyunsaturated fatty acid concentration. J. Dairy Sci. 2014, 97, 7307–7315. [Google Scholar] [CrossRef]
- Hansen, M.S.; Gregersen, S.B.; Rasmussen, J.T. Bovine milk processing impacts characteristics of extracellular vesicle isolates obtained by size-exclusion chromatography. Int. Dairy J. 2022, 127, 105212. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Z.; Sun, L.; Li, P. Fermentation results in quantitative changes in milk-derived exosomes and different effects on cell growth and survival. J. Agric. Food Chem. 2017, 65, 1220–1228. [Google Scholar] [CrossRef]
- Settachaimongkon, S.; Nout, M.J.R.; Antunes Fernandes, E.C.; Hettinga, K.A.; Vervoort, J.M.; van Hooijdonk, T.C.M.; Zwietering, M.H.; Smid, E.J.; Van Valenberg, H.J.F. Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp. bulgaricus on the metabolite profile of set-yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef]
- Sfakianakis, P.; Tzia, C. Conventional and innovative processing of milk for yogurt manufacture; development of texture and flavor: A review. Foods 2014, 3, 176–193. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.; Ressutte, J.; Bánkuti, S.; Bánkuti, F.; Pozza, M.; Madrona, G.S. Características tecnológicas, de qualidade e potencialidades da cadeia produtiva de queijo colonial na região sul do brasil: Uma Revisão. FTT J. Eng. Bus. 2017, 1, 50–64. [Google Scholar]
- Ressutte, J.B.; Rodrigues, T.S.; dos Santos Pozza, M.S.; Madrona, G.S. Application of Lactococcus Lactis Subsp. Lactis and Cremoris as starter culture in the colonial cheese production. J. Agric. Stud. 2020, 8, 561. [Google Scholar] [CrossRef][Green Version]
- Oh, S.; Park, M.R.; Ryu, S.; Maburutse, B.E.; Kim, J.U.; Kim, Y. Quantitative analysis of milk-derived microRNAs and microbiota during the manufacturing and ripening of soft cheese. J. Microbiol. Biotechnol. 2017, 27, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Yang, J.; Elbaz-Younes, I.; Primo, C.; Murungi, D.; Hirschi, K.D. Intestinal permeability, digestive stability and oral bioavailability of dietary small RNAs. Sci. Rep. 2018, 8, 10253. [Google Scholar] [CrossRef]
- Billa, P.A.; Faulconnier, Y.; Ye, T.; Chervet, M.; LeProvost, F.; Pires, J.A.A.; Leroux, C. Deep RNA-Seq reveals miRNome differences in mammary tissue of lactating Holstein and Montbéliarde cows. BMC Genom. 2019, 20, 621. [Google Scholar] [CrossRef]
- LeGuillou, S.; Leduc, A.; Laubier, J.; Barbey, S.; Rossignol, M.N.; Lefebvre, R.; Marthey, S.; Laloë, D.; LeProvost, F. Characterization of Holstein and Normande whole milk miRNomes highlights breed specificities. Sci. Rep. 2019, 9, 20345. [Google Scholar] [CrossRef]
- Wang, L.; Sadri, M.; Giraud, D.; Zempleni, J. RNase H2-Dependent Polymerase Chain Reaction and elimination of confounders in sample collection, storage, and analysis strengthen evidence that microRNAs in bovine milk are bioavailable in humans. J. Nutr. 2018, 148, 153–159. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| MiRNA | Mature Sequence |
|---|---|
| bta-mir-148a | UCAGUGCACUACAGAACUUUGU |
| bta-mir-21-5p | UAGCUUAUCAGACUGAUGUUGA |
| bta-mir-215 | AUGACCUAUGAAUUGACAGACA |
| bta-mir-29b | UAGCACCAUUUGAAAUCAGUGUU |
| bta-mir-30a-5p | UGUAAACAUCCUCGACUGGAAGCU |
| bta-mir-451 | AAACCGUUACCAUUACUGAGUU |
| bta-mir-7863 | AUGGACUGUCACCUGAGGAGC |
| cel-mir-238 | UUUGUACUCCGAUGCCAUUCAGA |
| cel-mir-39 | UCACCGGGUGUAAAUCAGCUUG |
| MiRNA | Raw Milk | Microwave | Yogurt | Cheese | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| bta-miR-148a | 4.62 | 0.72 | 3.44 | 0.87 | 3.04 | 0.16 | 2.17 | 0.63 |
| bta-miR-21-5p | 4.04 | 0.62 | 2.86 | 1.08 | 3.18 | 0.23 | 2.97 | 0.33 |
| bta-miR-215 | 3.90 | 0.85 | 2.60 | 1.24 | 2.86 | 0.28 | 1.70 | 0.38 |
| bta-miR-29b | 4.20 | 0.34 | 2.98 | 1.64 | 2.63 | 0.73 | 2.68 | 0.83 |
| bta-miR-30a-5p | 3.61 | 0.91 | 2.99 | 0.94 | 2.49 | 0.15 | 2.43 | 0.35 |
| bta-miR-451 | 5.24 | 0.42 | 3.17 | 1.99 | 3.06 | 0.28 | 2.46 | 0.40 |
| bta-miR-7863 | 4.28 | 0.39 | 2.33 | 1.75 | 3.21 | 0.18 | 2.39 | 0.36 |
| miRNA | Microwaved Milk | Yogurt | Cheese | |||
|---|---|---|---|---|---|---|
| Decrease (%) | p-Value | Decrease (%) | p-Value | Decrease (%) | p-Value | |
| bta-miR-148a | 25.53 | 0.043 | 34.22 | 0.005 | 53.15 | 0.005 |
| bta-miR-21-5p | 29.18 | 0.012 | 21.45 | 0.013 | 26.51 | 0.005 |
| bta-miR-215 | 33.20 | 0.012 | 26.58 | 0.013 | 56.32 | 0.005 |
| bta-miR-29b | 29.07 | 0.080 | 37.46 | 0.028 | 36.15 | 0.028 |
| bta-miR-30a-5p | 17.20 | 0.063 | 31.04 | 0.013 | 32.73 | 0.013 |
| bta-miR-451 | 39.42 | 0.018 | 41.62 | 0.005 | 53.09 | 0.005 |
| bta-miR-7863 | 45.45 | 0.123 | 24.84 | 0.005 | 44.01 | 0.005 |
| Mean | 31.29 | 31.03 | 43.14 | |||
| MiRNA | Raw Milk | Cheese | p-Value (Wilcoxon Test) | ||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| bta-miR-148a | 2.32 | 1.35 | 2.10 | 0.30 | 0.76 |
| bta-miR-21-5p | 2.30 | 2.01 | 2.97 | 1.64 | 0.33 |
| bta-miR-215 | 1.68 | 2.06 | 1.60 | 0.17 | >0.99 |
| bta-miR-29b | 1.72 | 2.26 | 3.16 | 1.63 | 0.11 |
| bta-miR-30a-5p | 1.77 | 1.95 | 2.89 | 0.33 | 0.13 |
| bta-miR-451 | 2.24 | 2.58 | 1.50 | 0.11 | 0.31 |
| bta-miR-7863 | 1.99 | 1.46 | 2.43 | 1.21 | 0.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou el qassim, L.; Martínez, B.; Rodríguez, A.; Dávalos, A.; López de las Hazas, M.-C.; Menéndez Miranda, M.; Royo, L.J. Effects of Cow’s Milk Processing on MicroRNA Levels. Foods 2023, 12, 2950. https://doi.org/10.3390/foods12152950
Abou el qassim L, Martínez B, Rodríguez A, Dávalos A, López de las Hazas M-C, Menéndez Miranda M, Royo LJ. Effects of Cow’s Milk Processing on MicroRNA Levels. Foods. 2023; 12(15):2950. https://doi.org/10.3390/foods12152950
Chicago/Turabian StyleAbou el qassim, Loubna, Beatriz Martínez, Ana Rodríguez, Alberto Dávalos, María-Carmen López de las Hazas, Mario Menéndez Miranda, and Luis J. Royo. 2023. "Effects of Cow’s Milk Processing on MicroRNA Levels" Foods 12, no. 15: 2950. https://doi.org/10.3390/foods12152950
APA StyleAbou el qassim, L., Martínez, B., Rodríguez, A., Dávalos, A., López de las Hazas, M.-C., Menéndez Miranda, M., & Royo, L. J. (2023). Effects of Cow’s Milk Processing on MicroRNA Levels. Foods, 12(15), 2950. https://doi.org/10.3390/foods12152950