
Fermentation Efficiency of Genetically Modified Yeasts in Grapes Must


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Yeast Strains, Media, and Growth Conditions
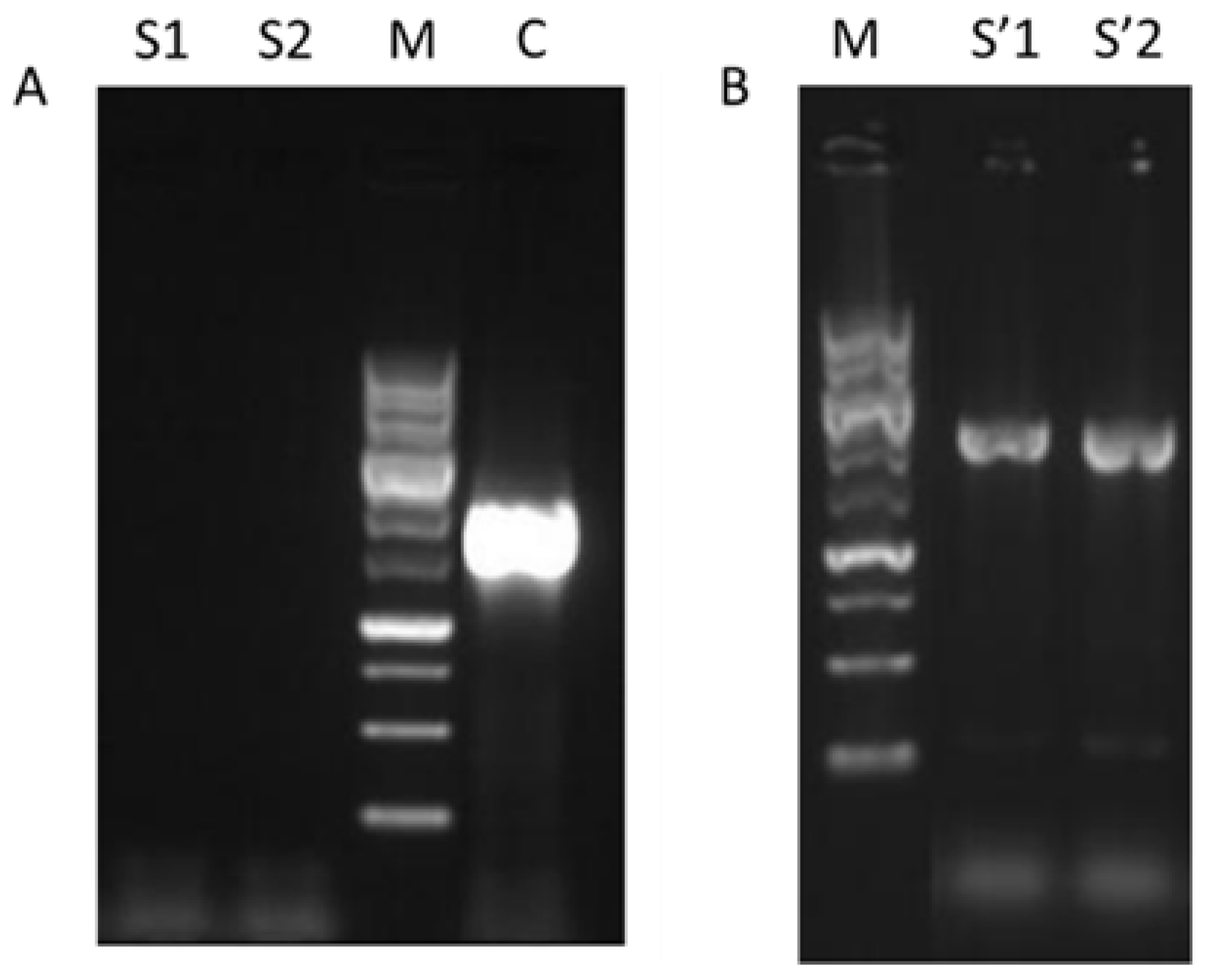
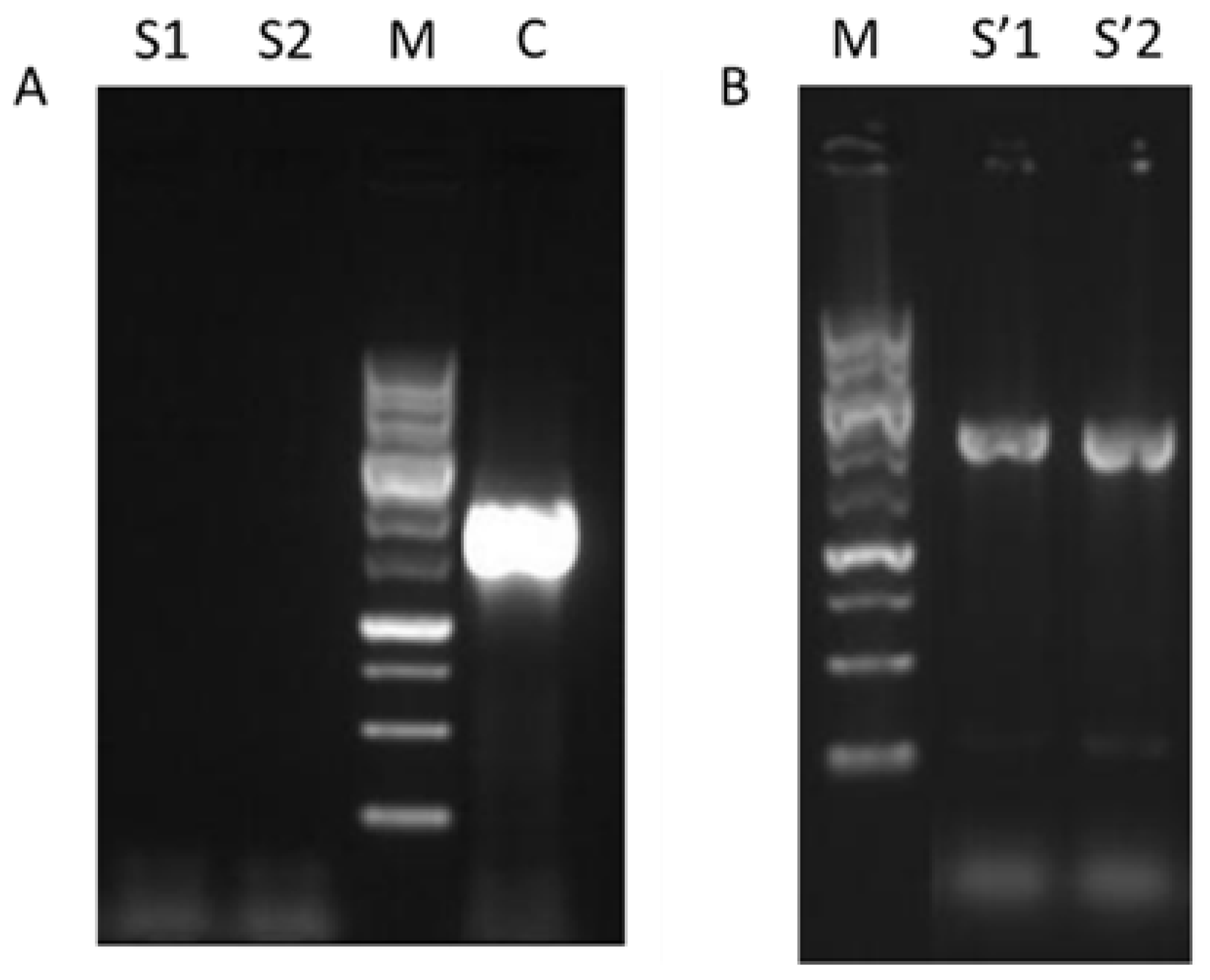
2.3. W_M4_533 Mutant Generation
2.4. Must Preparation, Inoculation and Sampling
2.5. Reverse Flow Gas Chromatography (RFGC)
2.6. Activation Energy Determination
2.7. HPLC Analysis
2.8. Statistical Analysis
- The standard error of calculated slope was determined.
- t-value was calculated
- The students t-distribution (p) was used to test the hypothesis of equal slopes.
- Statistical analyses were performed using Microcal Origin v.9.0 (Microcal Software Inc., Northampton, MA, USA).
3. Results
3.1. Generation of W_M4_533
3.2. Determination of Ethanol Production Using RFGC
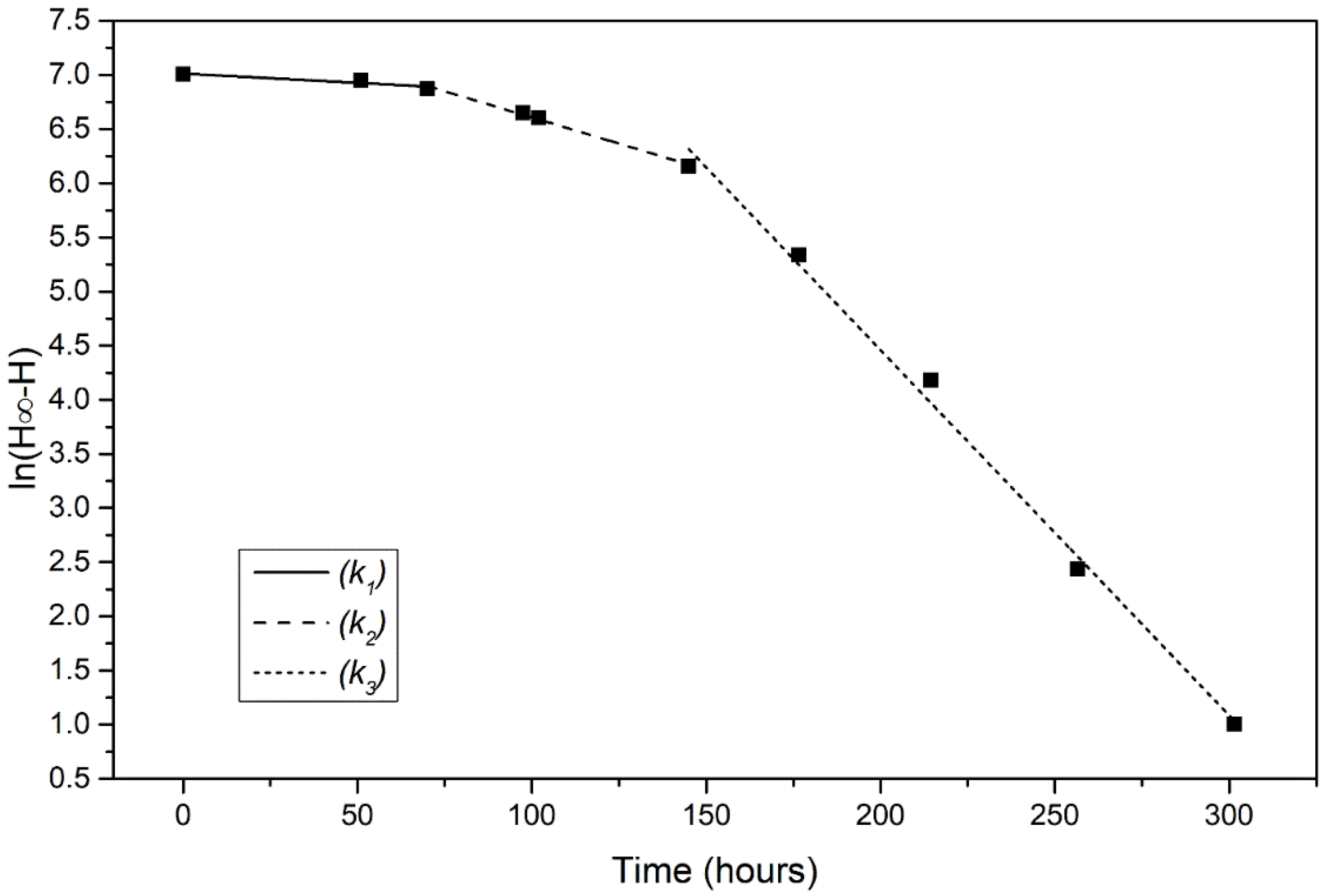
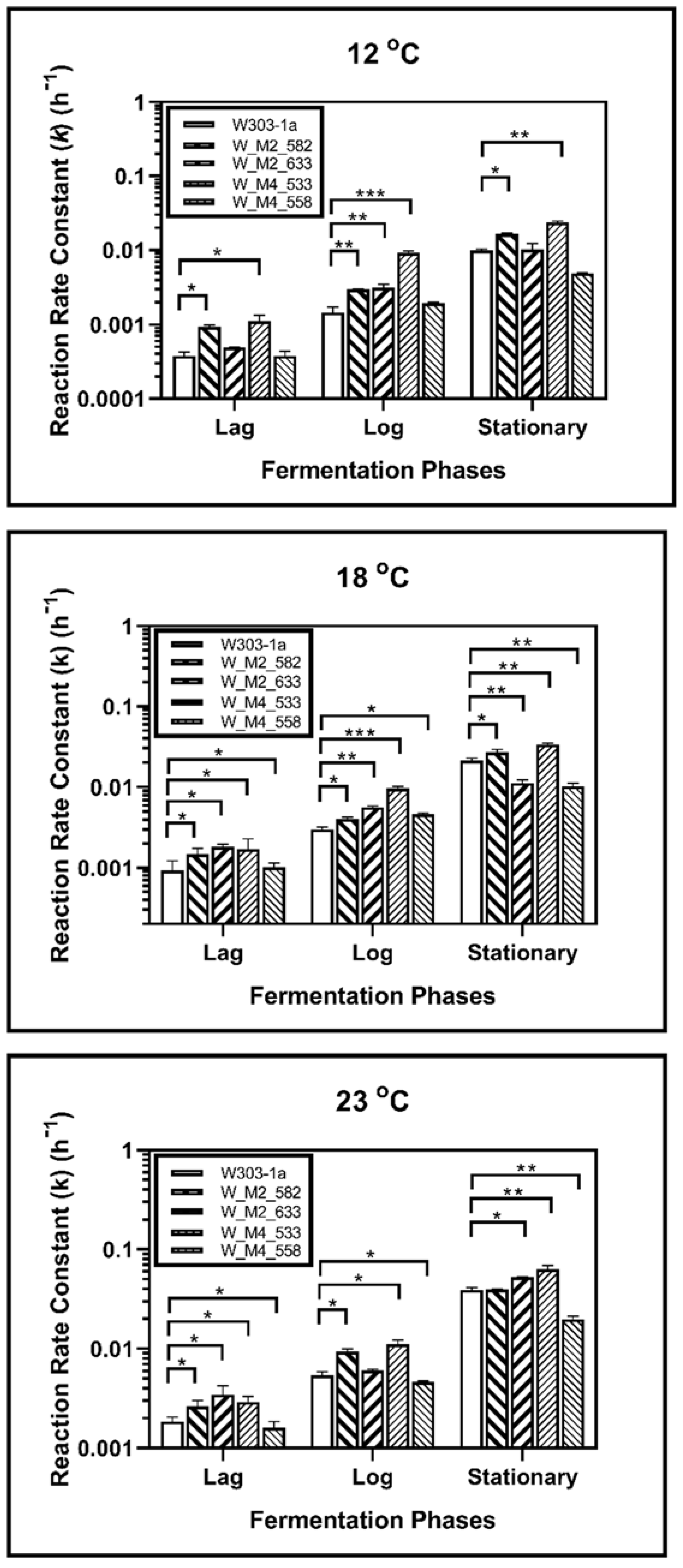
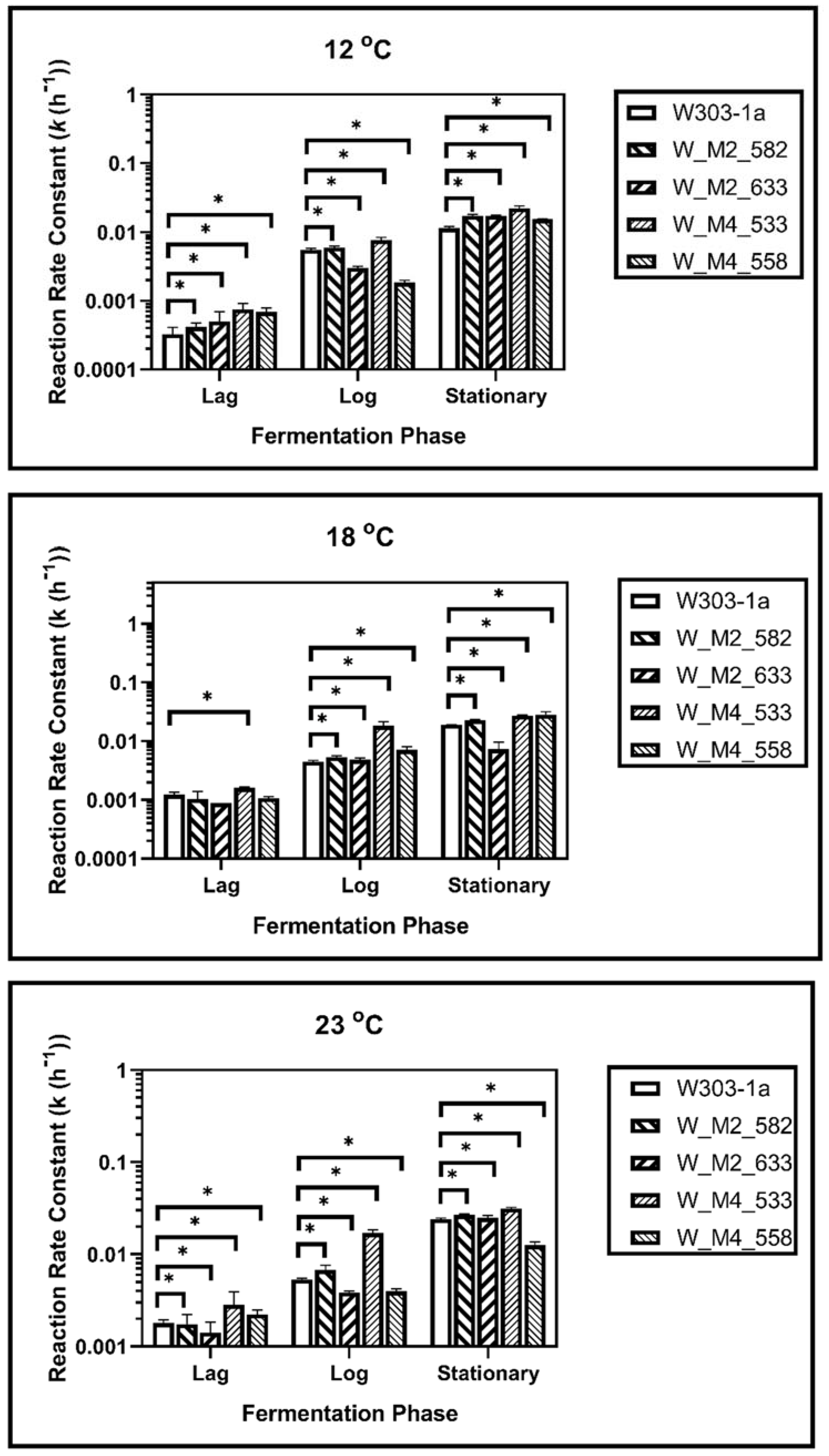
3.3. Kinetic Analysis
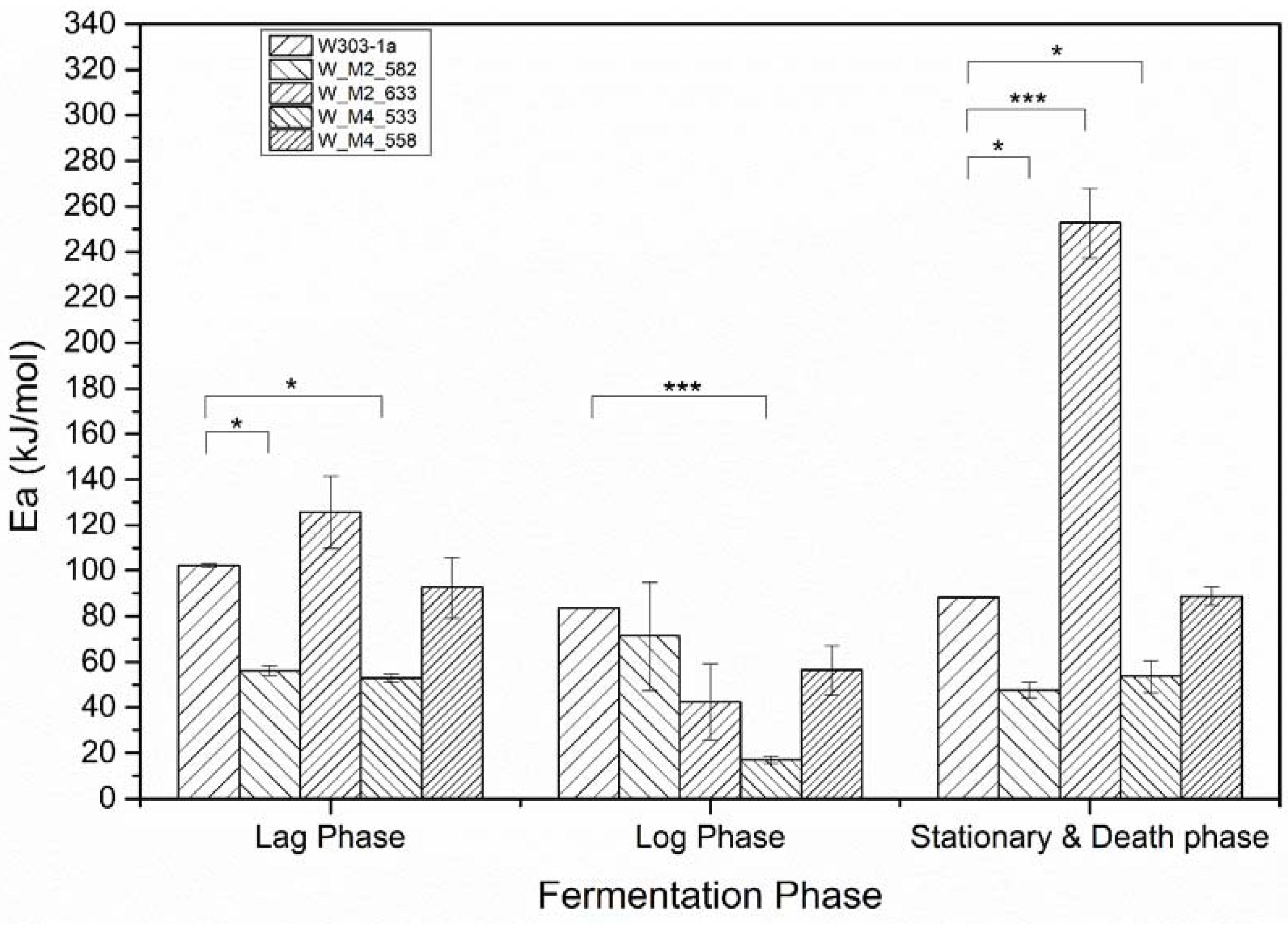
3.4. Activation Energy
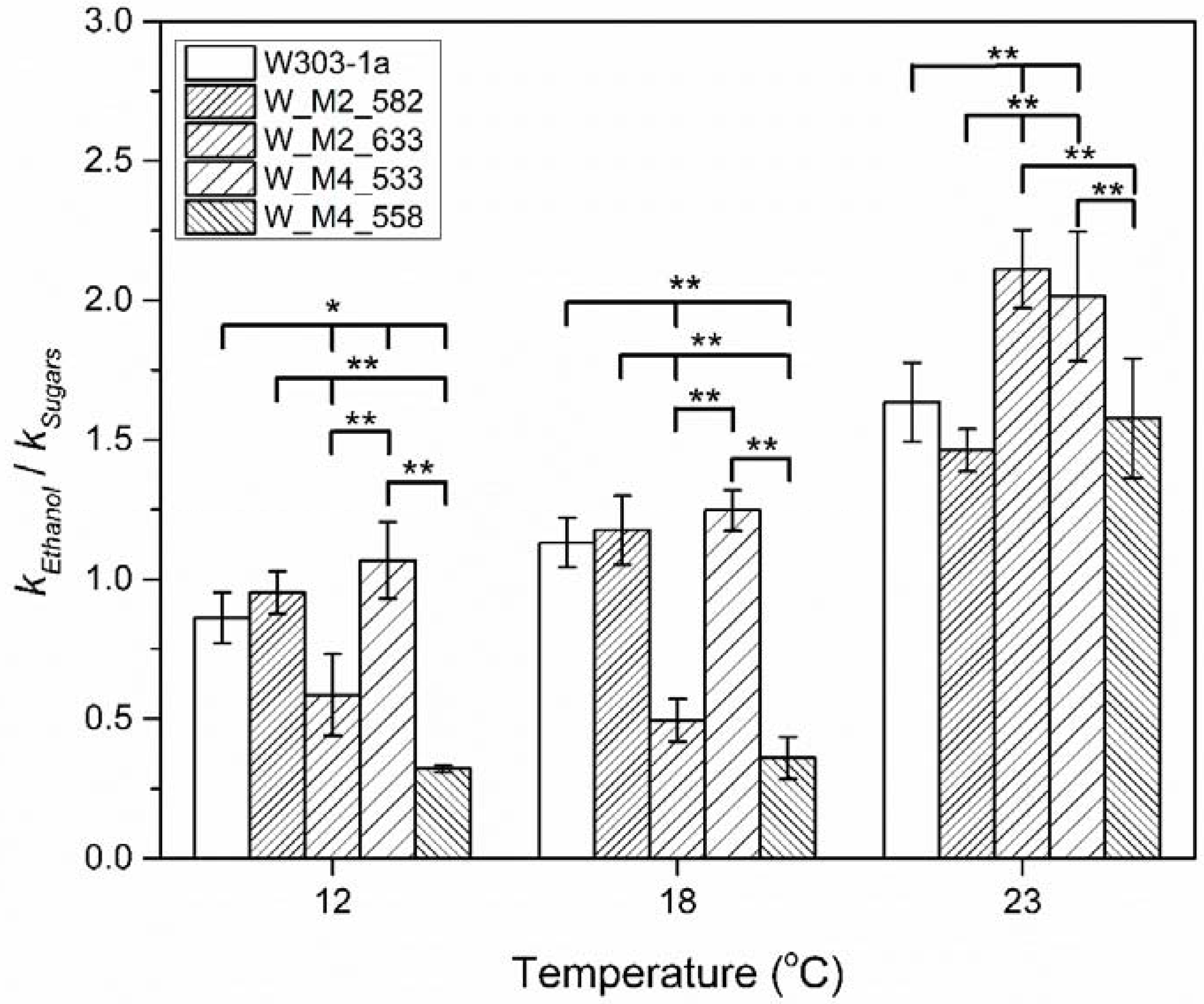
3.5. Sugar Consumption
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Garcia, M.A.; Oliva, J.; Barba, A.; Camara, M.A.; Pardo, F.; Diaz-Plaza, E.M. Effect of fungicide residues on the aromatic composition of white wine inoculated with three Saccharomyces cerevisiae strains. J. Agric. Food Chem. 2004, 52, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Estruch, F.; Carlson, M. Two homologous zinc finger genes identified by multicopy suppression in a SNF1 protein kinase mutant of Saccharomyces cerevisiae. Mol. Cell Biol. 1993, 13, 3872–3881. [Google Scholar] [CrossRef] [PubMed]
- Gorner, W.; Durchschlag, E.; Martinez-Pastor, M.T.; Estruch, F.; Ammerer, G.; Hamilton, B.; Ruis, H.; Schuller, C. Nuclear localization of the C2H2 zinc finger protein Msn2p is regulated by stress and protein kinase A activity. Genes Dev. 1998, 12, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Ward, M.P.; Garrett, S. Yeast PKA represses Msn2p/Msn4p-dependent gene expression to regulate growth, stress response and glycogen accumulation. EMBO J. 1998, 17, 3556–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, F. Stress-controlled transcription factors, stress-induced genes and stress tolerance in budding yeast. FEMS Microbiol. Rev. 2000, 24, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Pfanzagl, V.; Gorner, W.; Radolf, M.; Parich, A.; Schuhmacher, R.; Strauss, J.; Reiter, W.; Schuller, C. A constitutive active allele of the transcription factor Msn2 mimicking low PKA activity dictates metabolic remodeling in yeast. Mol. Biol. Cell 2018, 29, 2848–2862. [Google Scholar] [CrossRef] [PubMed]
- Rajvanshi, P.K.; Arya, M.; Rajasekharan, R. The stress-regulatory transcription factors Msn2 and Msn4 regulate fatty acid oxidation in budding yeast. J. Biol. Chem. 2017, 292, 18628–18643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, Z.; Pinglay, S.; Ji, H.; Boeke, J.D. Msn2/4 regulate expression of glycolytic enzymes and control transition from quiescence to growth. Elife 2017, 6, e29938. [Google Scholar] [CrossRef] [Green Version]
- Vamvakas, S.S.; Kapolos, J.; Farmakis, L.; Genneos, F.; Damianaki, M.E.; Chouli, X.; Vardakou, A.; Liosi, S.; Stavropoulou, E.; Leivaditi, E.; et al. Specific serine residues of Msn2/4 are responsible for regulation of alcohol fermentation rates and ethanol resistance. Biotechnol. Prog. 2019, 35, e2759. [Google Scholar] [CrossRef]
- Vamvakas, S.S.; Kapolos, J.; Farmakis, L.; Koskorellou, G.; Genneos, F. Ser625 of msn2 transcription factor is indispensable for ethanol tolerance and alcoholic fermentation process. Biotechnol. Prog. 2019, 35, e2837. [Google Scholar] [CrossRef] [PubMed]
- Al-sahlany, S.T.G.; Altemimi, A.B.; Al-Manhel, A.J.A.; Niamah, A.K.; Lakhssassi, N.; Ibrahim, S.A. Purification of bioactive peptide with antimicrobial properties produced by Saccharomyces cerevisiae. Foods 2020, 9, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, C.L.; Pei, G.K. Modification of a PCR-based site-directed mutagenesis method. Biotechniques 1997, 23, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Fukuda, Y.; Murata, K.; Kimura, A. Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 1983, 153, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, J.L. Introduction to Wine Laboratory Practices and Procedures; Springer: Berlin/Heidelberg, Germany, 2006; p. 258. [Google Scholar]
- Lainioti, G.; Kapolos, J.; Koliadima, A.; Karaiskakis, G. New separation methodologies for the distinction of the growth phases of Saccharomyces cerevisiae cell cycle. J. Chromatogr. A 2010, 1217, 1813–1820. [Google Scholar] [CrossRef]
- Kapolos, J.; Katsanos, N.A.; Niotis, A. Physicochemical quantities in catalytic reactions measured simultaneously by gas chromatography. Chromatographia 1989, 27. [Google Scholar] [CrossRef]
- Economopoulos, N.; Athanassopoulos, N.; Katsanos, N.A.; Karaiskakis, G.; Agathonos, P.; Vassilakos, C. A Plant Kinetic Study of Alcoholic Fermentation Using Reversed-Flow Gas Chromatography. Sep. Sci. Technol. 1992, 27, 2055–2070. [Google Scholar] [CrossRef]
- Roels, J.A. Energetics and Kinetics in Biotechnology; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1983; Volume 330, pp. 333–339. [Google Scholar]
- Converti, A.; Bargagliotti, C.; Cavanna, C.; Nicolella, C.; Borghi, M.D.; Nicomla, C.; Del Borghi, M. Evaluation of kinetic parameters and thermodynamic quantities of starch hydrolysate alcohol fermentation by Saccharomyces cerevisiae. Bioprocess. Eng. 1996, 15, 63–69. [Google Scholar] [CrossRef]
- Thevelein, J.M.; Cauwenberg, L.; Colombo, S.; De Winde, J.H.; Donation, M.; Dumortier, F.; Kraakman, L.; Lemaire, K.; Ma, P.; Nauwelaers, D.; et al. Nutrient-induced signal transduction through the protein kinase A pathway and its role in the control of metabolism, stress resistance, and growth in yeast. Enzyme. Microb. Technol. 2000, 26, 819–825. [Google Scholar] [CrossRef]
- Endoh, R.; Horiyama, M.; Ohkuma, M. D-Fructose Assimilation and Fermentation by Yeasts Belonging to Saccharomycetes: Rediscovery of Universal Phenotypes and Elucidation of Fructophilic Behaviors in Ambrosiozyma platypodis and Cyberlindnera americana. Microorganisms 2021, 9, 758. [Google Scholar] [CrossRef]
- Berthels, N.J.; Cordero Otero, R.R.; Bauer, F.F.; Thevelein, J.M.; Pretorius, I.S. Discrepancy in glucose and fructose utilisation during fermentation by Saccharomyces cerevisiae wine yeast strains. FEMS Yeast. Res. 2004, 4, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillaume, C.; Delobel, P.; Sablayrolles, J.M.; Blondin, B. Molecular basis of fructose utilization by the wine yeast Saccharomyces cerevisiae: A mutated HXT3 allele enhances fructose fermentation. Appl. Environ. Microbiol. 2007, 73, 2432–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.K. The chemistry and biochemistry of the sweetness of sugars. Adv. Carbohydr. Chem. Biochem. 1987, 45, 199–351. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Ref. |
|---|---|---|
| W303-1a | MAT a, ura3–1, leu2–3, his3–11, trp1–1, ade2–1, can1–1000 | [10] |
| W_M2_582 | MAT a, ura3–1, leu2–3, his3–11, trp1–1, ade2–1, can1–1000, msn2-S582A | [10] |
| W_M2_633 | MAT a, ura3–1, leu2–3, his3–11, trp1–1, ade2–1, can1–1000, msn2-S633A | [10] |
| W_M4_533 | MAT a, ura3–1, leu2–3, his3–11, trp1–1, ade2–1, can1–1000, msn4-S531A | This Study |
| W_M4_558 | MAT a, ura3–1, leu2–3, his3–11, trp1–1, ade2–1, can1–1000, msn4-S558A | [10] |
| Primer | Sequence |
|---|---|
| MSN4_URA3_F | 5′CAGTTCGGCTTTTTTTTCTTTTCTTCTTATTAAAAACAATATAATGTCGAAAGCTACATATAAGG 3′ |
| MSN4_URA3_R | 5′CCGTAGCTTGTCTTGCTTTTATTTGCTTTTGACCTTATTTTTTTTAGTTTTGCTGGCCGCATC 3′ |
| MSN4_533_F | 5′ GAAGAAGAAAGTCGTCAGCTGTTACTTTAAGTCCAAC 3′ |
| MSN4_533_R | 5′ GTTGGACTTAAAGTAACAGCTGACGACTTTCTTCTTC 3′ |
| MSN4_F | 5′CAGTTCGGCTTTTTTTTCTTTTCTTCTTATTAAAAACAATATAATGCTAGTCTTCGGACCTAA 3′ |
| MSN4_R | 5′CCGTAGCTTGTCTTGCTTTTATTTGCTTTTGACCTTATTTTTTTCAAAAATCACCGTGCTTTTTGTG 3′ |
| W303-1a | W_M2_582 | W_M2_633 | W_M4_533 | W_M4_558 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp (°C) | Phase | k (h−1) × 10−3 | SEM* | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM |
| 12 | Lag | 0.327 | 0.085 | 0.416 * | 0.057 | 0.504 * | 0.198 | 0.754 * | 0.164 | 0.688 * | 0.099 |
| Log | 5.550 | 0.269 | 6.020 * | 0.223 | 3.030 * | 0.167 | 7.660 * | 0.768 | 1.840 * | 0.146 | |
| Stationary | 11.46 | 0.678 | 17.26 * | 0.856 | 17.32 * | 0.504 | 22.24 * | 1.990 | 15.40 * | 0.292 | |
| 18 | Lag | 1.230 | 0.133 | 1.040 | 0.353 | 0.884 | 0.001 | 1.600 * | 0.075 | 1.080 | 0.068 |
| Log | 4.480 | 0.173 | 5.340 * | 0.231 | 5.690 * | 0.286 | 18.26 * | 3.230 | 7.050 * | 0.921 | |
| Stationary | 18.85 | 0.208 | 22.86 * | 0.352 | 22.57 * | 1.510 | 27.08 * | 0.453 | 28.25 * | 3.44 | |
| 23 | Lag | 1.790 | 0.167 | 1.750 * | 0.488 | 1.420 * | 0.416 | 2.840 * | 1.070 | 2.230 * | 0.279 |
| Log | 5.370 | 0.102 | 6.780 * | 0.866 | 3.850 * | 0.163 | 17.03 * | 1.420 | 3.960 * | 0.268 | |
| Stationary | 24.05 | 0.649 | 26.90 * | 0.784 | 24.91 * | 1.520 | 31.42 * | 0.867 | 12.65 * | 0.940 | |
| W303-1a | W_M2_582 | W_M2_633 | W_M4_533 | W_M4_558 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp (°C) | Phase | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM |
| 12 | Lag | 0.337 | 0.126 | 0.623 * | 0.087 | 1.090 * | 0.297 | 0.894 * | 0.034 | 1.130 * | 0.126 |
| Log | 2.340 | 0.355 | 9.780 * | 0.460 | 3.610 * | 0.559 | 6.760 * | 0.803 | 2.860 * | 0.190 | |
| Stationary | 13.72 | 0.537 | 20.08 * | 1.700 | 13.05 | 0.595 | 21.92 * | 0.480 | 16.90 * | 0.275 | |
| 18 | Lag | 1.960 | 0.068 | 2.600 * | 0.749 | 1.510 * | 0.093 | 2.090 | 0.191 | 1.670 | 0.112 |
| Log | 6.580 | 0.152 | 10.80 * | 0.791 | 9.830 | 0.411 | 17.80 * | 0.120 | 10.00 * | 1.050 | |
| Stationary | 25.76 | 0.428 | 20.63 * | 0.539 | 23.00 * | 0.686 | 22.75 * | 0.307 | 33.90 * | 1.050 | |
| 23 | Lag | 2.710 | 0.294 | 2.600 | 0.749 | 1.670 * | 0.151 | 1.440 * | 0.032 | 4.66 * | 0.260 |
| Log | 9.000 | 1.001 | 10.78 * | 0.791 | 5.600 * | 0.155 | 14.20 * | 2.550 | 15.65 * | 2.840 | |
| Stationary | 30.92 | 1.310 | 24.62 * | 0.894 | 23.89 * | 1.880 | 40.70 * | 0.950 | 47.45 * | 5.180 | |
| W303-1a | W_M2_582 | W_M2_633 | W_M4_533 | W_M4_558 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp (°C) | Phase | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM | k (h−1) × 10−3 | SEM |
| 12 | Lag | 0.100 | 0.006 | 0.285 * | 0.082 | 0.102 | 0.038 | 0.848 * | 0.098 | 0.634 * | 0.084 |
| Log | 2.790 | 0.174 | 2.960 | 0.121 | 1.470 * | 0.197 | 4.960 * | 0.588 | 4.150 * | 0.282 | |
| Stationary | 13.22 | 0.947 | 17.07 * | 0.789 | 15.35 * | 1.250 | 22.45 * | 2.200 | 15.29 * | 0.312 | |
| 18 | Lag | 0.610 | 0.185 | 1.040 * | 0.281 | 0.725 | 0.071 | 1.390 * | 0.149 | 1.570 * | 0.327 |
| Log | 3.380 | 0.120 | 4.440 * | 1.090 | 3.530 * | 0.426 | 7.420 * | 0.004 | 8.340 * | 0.934 | |
| Stationary | 17.34 | 0.352 | 24.58 * | 1.650 | 21.38 * | 0.130 | 26.91 * | 0.355 | 28.10 * | 3.600 | |
| 23 | Lag | 1.08 | 0.089 | 1.040 * | 0.281 | 0.763 * | 0.184 | 2.030 * | 0.006 | 2.150 * | 0.220 |
| Log | 3.520 | 0.216 | 4.440 * | 1.090 | 4.010 | 0.404 | 11.59 * | 1.350 | 12.59 * | 1.320 | |
| Stationary | 20.08 | 1.620 | 24.58 * | 1.650 | 24.49 * | 1.660 | 31.17 * | 0.413 | 38.86 * | 2.580 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kassoumi, K.; Kousoulou, P.; Sevastos, D.; Vamvakas, S.-S.; Papadimitriou, K.; Kapolos, J.; Koliadima, A. Fermentation Efficiency of Genetically Modified Yeasts in Grapes Must. Foods 2022, 11, 413. https://doi.org/10.3390/foods11030413
Kassoumi K, Kousoulou P, Sevastos D, Vamvakas S-S, Papadimitriou K, Kapolos J, Koliadima A. Fermentation Efficiency of Genetically Modified Yeasts in Grapes Must. Foods. 2022; 11(3):413. https://doi.org/10.3390/foods11030413
Chicago/Turabian StyleKassoumi, Konstantina, Penny Kousoulou, Dimitrios Sevastos, Sotirios-Spyridon Vamvakas, Konstantinos Papadimitriou, John Kapolos, and Athanasia Koliadima. 2022. "Fermentation Efficiency of Genetically Modified Yeasts in Grapes Must" Foods 11, no. 3: 413. https://doi.org/10.3390/foods11030413
APA StyleKassoumi, K., Kousoulou, P., Sevastos, D., Vamvakas, S.-S., Papadimitriou, K., Kapolos, J., & Koliadima, A. (2022). Fermentation Efficiency of Genetically Modified Yeasts in Grapes Must. Foods, 11(3), 413. https://doi.org/10.3390/foods11030413