
Green/Roasted Coffee and Silverskin Extracts Inhibit Sugar Absorption by Human Intestinal Epithelial (Caco-2) Cells by Decreasing GLUT2 Gene Expression

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Standards
2.2. Samples and Sample Preparation
2.3. Ultrasound-Assisted Extraction
2.4. RP-HPLC-DAD Analysis
2.5. Cellular Assays
2.5.1. Caco-2 Cell Culture
2.5.2. Cell Treatments
2.5.3. Evaluation of Uptake of 3H-DG and 14C-FRU by Caco-2 Cells
2.5.4. Determination of Cell Viability
2.5.5. Determination of Culture Mass
2.5.6. Quantitative Reverse Transcription Real-Time PCR
2.5.7. Total Protein Determination
2.6. Statistical Analysis
3. Results and Discussion
3.1. CGA Profile and Caffeine Contents of the Different Extracts
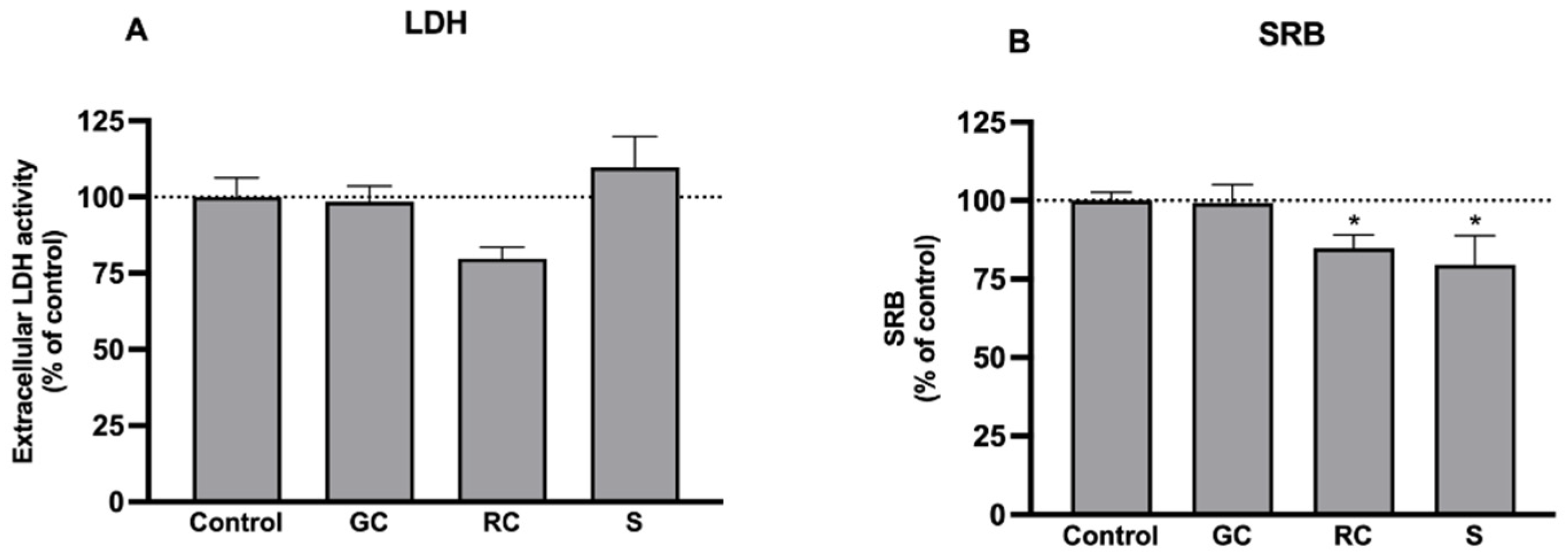
3.2. Effects on Cell Viability and Culture Mass
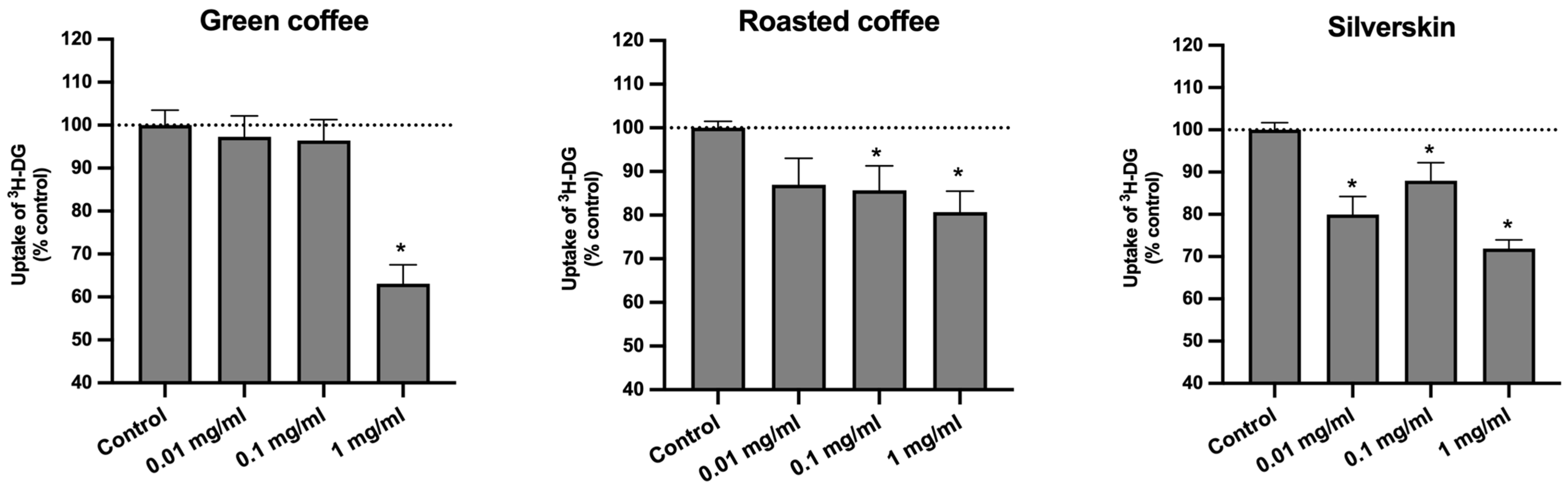
3.3. Effects on the Uptake of 3H-DG
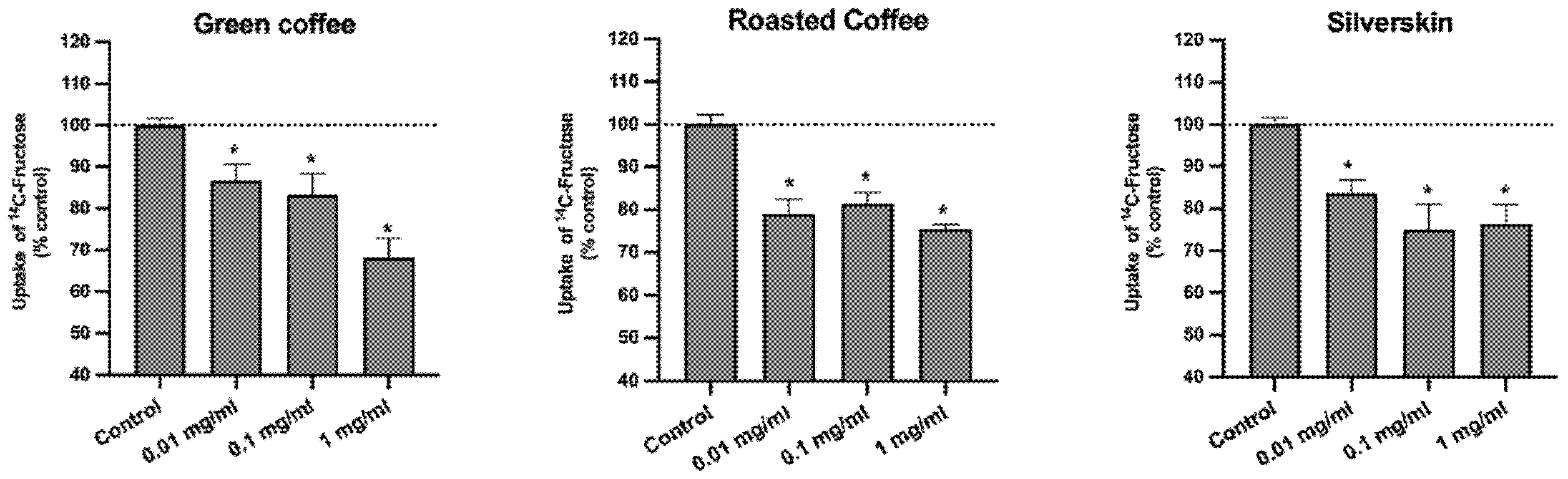
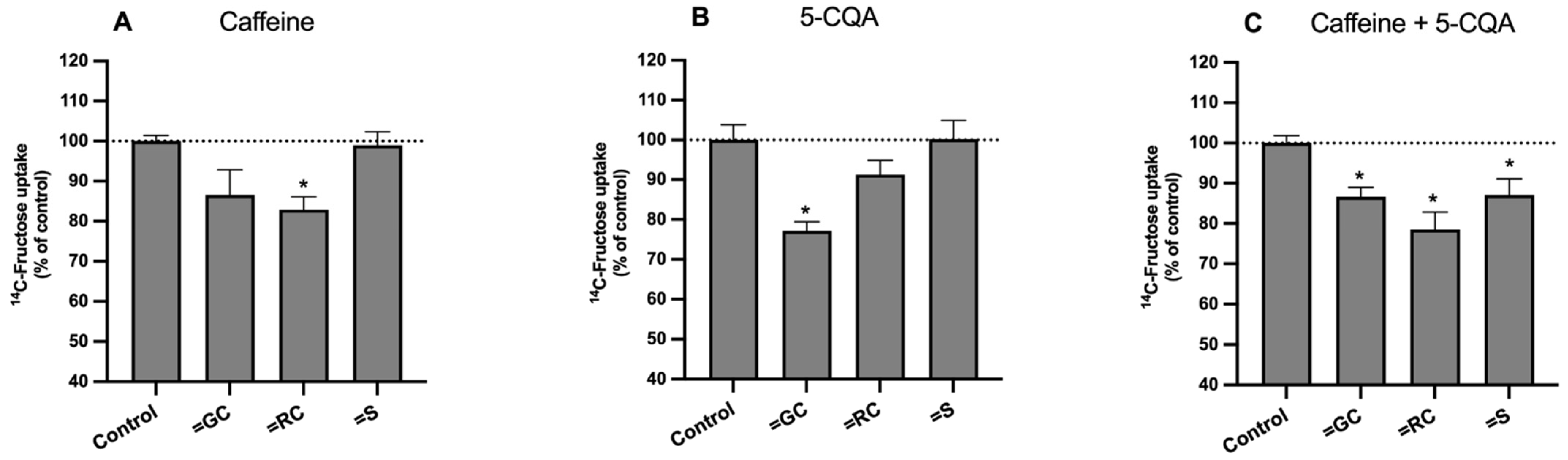
3.4. Effect on the Uptake of 14C-FRU
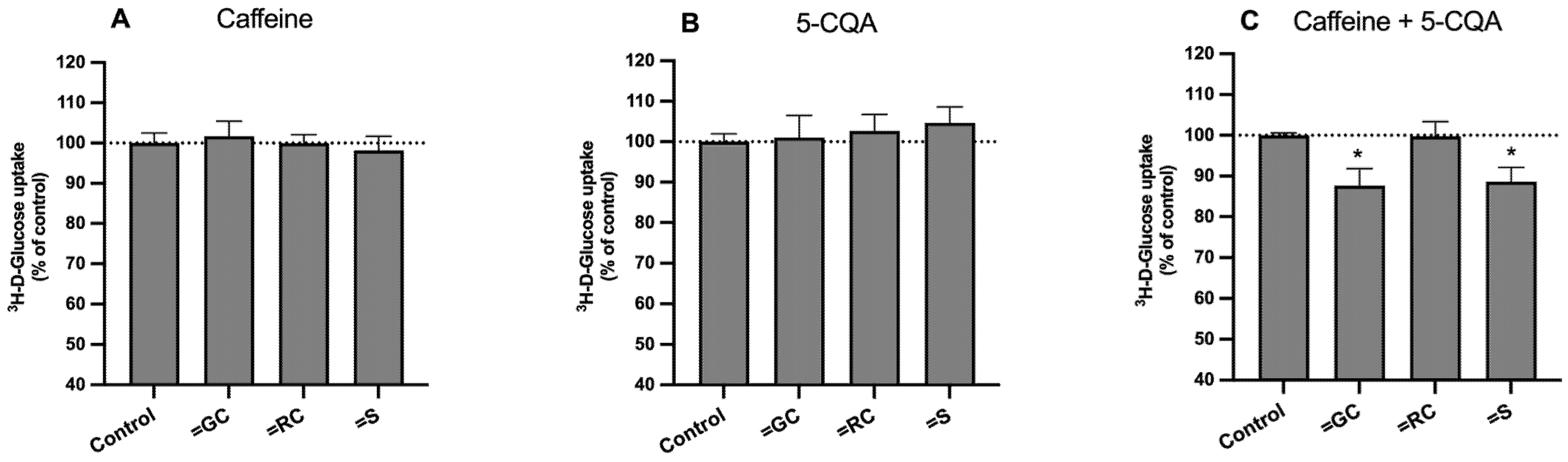
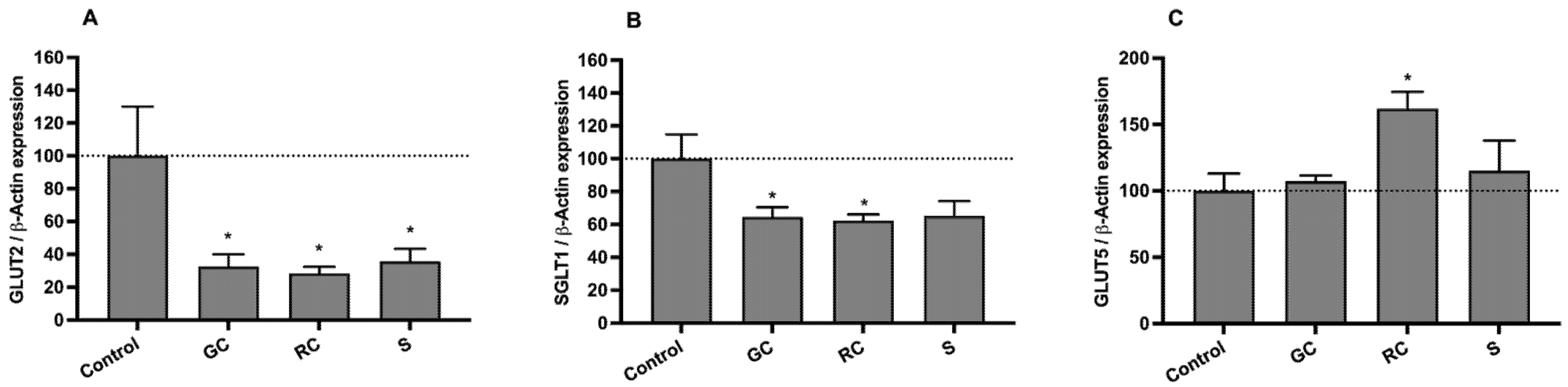
3.5. Effect on on SGLT1, GLUT2, and GLUT5 mRNA Levels
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Magalhães Júnior, A.I.; do Prado, F.G.; Pagnoncelli, M.G.B.; Karp, S.G.; Soccol, C.R. Chemical composition and health properties of coffee and coffee by-products. Adv. Food Nutr. Res. 2020, 91, 65–96. [Google Scholar] [PubMed]
- Farah, A.; de Paula Lima, J. Consumption of chlorogenic acids through coffee and health implications. Beverages 2019, 5, 11. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.J.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funct. 2014, 5, 1695–1717. [Google Scholar] [CrossRef]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez de Meija, E.; Ramirez-Mares, M.V. Impact of caffeine and coffee on our health. Trends Endocrinol. Metab. 2014, 25, 489–492. [Google Scholar]
- Alves, R.C.; Costa, A.S.G.; Jerez, M.; Casal, S.; Sineiro, J.; Núñez, M.J.; Oliveira, M.B.P.P. Antiradical activity, phenolics profile, and hydroxymethylfurfural in espresso coffee: Influence of technological factors. J. Agric. Food Chem. 2010, 58, 12221–12229. [Google Scholar] [CrossRef] [PubMed]
- International Coffee Organization. World Coffee Consumption. Available online: http://www.ico.org/prices/new-consumption-table.pdf (accessed on 28 September 2022).
- Alves, R.C.; Rodrigues, F.; Nunes, M.A.; Vinha, A.F.; Oliveira, M.B.P.P. State of the art in coffee processing by-products. In Handbook of Coffee Processing By-Products; Galanakis, C.M., Ed.; Academic Press: London, UK, 2017; pp. 1–26. [Google Scholar]
- Costa, A.S.G.; Alves, R.C.; Vinha, A.F.; Costa, E.; Costa, C.S.G.; Nunes, M.A.; Almeida, A.A.; Santos-Silva, A.; Oliveira, M.B.P.P. Nutritional, chemical and antioxidant/pro-oxidant profiles of silverskin, a coffee roasting by-product. Food Chem. 2018, 267, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Bessada, S.M.F.; Alves, R.C.; Oliveira, M.B.P.P. Coffee silverskin: A review on potential cosmetic applications. Cosmetics 2018, 5, 5. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Alves, R.C.; Costa, A.S.G.; Nunes, M.A.; Oliveira, M.B.P.P. Coffea canephora silverskin from different geographical origins: A comparative study. Sci. Total Environ. 2018, 645, 1021–1028. [Google Scholar] [CrossRef]
- Loureiro, G.; Martel, F. The effect of dietary polyphenols on intestinal absorption of glucose and fructose: Relation with obesity and type 2 diabetes. Food Rev. Int. 2019, 35, 390–406. [Google Scholar] [CrossRef]
- Andrade, N.; Peixoto, J.A.B.; Oliveira, M.B.P.P.; Martel, F.; Alves, R.C. Can coffee silverskin be a useful tool to fight metabolic syndrome? Front. Nutr. 2022, 9, 966734. [Google Scholar] [CrossRef] [PubMed]
- Kelishadi, R.; Mansourian, M.; Heidari-Beni, M. Association of fructose consumption and components of metabolic syndrome in human studies: A systematic review and meta-analysis. Nutrition 2014, 30, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.-R.; Packard, C.J.; Borén, J. Dietary fructose and the metabolic syndrome. Nutrients 2019, 11, 1987. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Sánchez-Lozada, L.G.; Andrews, P.; Lanaspa, M.A. Perspective: A historical and scientific perspective of sugar and its relation with obesity and diabetes. Adv. Nutr. 2017, 8, 412–422. [Google Scholar] [CrossRef]
- Gromova, L.V.; Fetissov, S.O.; Gruzdkov, A.A. Mechanisms of glucose absorption in the small intestine in health and metabolic diseases and their role in appetite regulation. Nutrients 2021, 13, 2474. [Google Scholar] [CrossRef]
- Bassoli, B.K.; Cassolla, P.; Borba-Murad, G.R.; Constantin, J.; Salgueiro-Pagadigorria, C.L.; Bazotte, R.B.; da Silva, R.S.S.; de Souza, H.M. Chlorogenic acid reduces the plasma glucose peak in the oral glucose tolerance test: Effects on hepatic glucose release and glycaemia. Cell Biochem. Funct. 2008, 26, 320–328. [Google Scholar] [CrossRef]
- Johnston, K.L.; Clifford, M.N.; Morgan, L.M. Coffee acutely modifies gastrointestinal hormone secretion and glucose tolerance in humans: Glycemic effects of chlorogenic acid and caffeine. Am. J. Clin. Nutr. 2003, 78, 728–733. [Google Scholar] [CrossRef]
- Ontawong, A.; Duangjai, A.; Srimaroeng, C. Coffea arabica bean extract inhibits glucose transport and disaccharidase activity in Caco-2 cells. Biomed. Rep. 2021, 15, 73. [Google Scholar] [CrossRef]
- Thom, E. The effect of chlorogenic acid enriched coffee on glucose absorption in healthy volunteers and its effects on body mass when used long-term in overweight and obese people. J. Int. Med. Res. 2007, 35, 900–908. [Google Scholar] [CrossRef]
- Tunnicliffe, J.M.; Eller, L.K.; Reimer, R.A.; Hittel, D.S.; Shearer, J. Chlorogenic acid differentially affects postpandrial glucose and glucose-dependent insulinotropic polypeptide response in rats. Appl. Physiol. Nutr. Metab. 2011, 36, 650–659. [Google Scholar] [CrossRef]
- Welsch, C.A.; Lachance, P.A.; Wasserman, B.P. Dietary phenolic compounds: Inhibition of Na+-dependent D-glucose uptake in rat intestinal brush border membrane vesicles. J. Nutr. 1989, 119, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Lea, T. Caco-2 cell line. In The Impact of Food Bioactives on Gut Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Berlin, Germany, 2015; pp. 103–111. [Google Scholar]
- O’Brien, P.; Corpe, C.P. Acute effects of sugars and artificial sweeteners on small intestinal sugar transport: A study using Caco-2 cells as an in vitro model of the human enterocyte. PLoS ONE 2016, 11, e0167785. [Google Scholar] [CrossRef] [PubMed]
- Puga, H.; Alves, R.C.; Costa, A.S.; Vinha, A.F.; Oliveira, M.B.P.P. Multi-frequency multimode modulated technology as a clean, fast, and sustainable process to recover antioxidants from a coffee by-product. J. Clean. Prod. 2017, 168, 14–21. [Google Scholar] [CrossRef]
- Andrade, N.; Silva, C.; Martel, F. The effect of oxidative stress upon intestinal sugar transport: An in vitro study using human intestinal epithelial (Caco-2) cells. Toxicol. Res. 2018, 7, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, J.A.B.; Andrade, N.; Machado, S.; Costa, A.S.G.; Puga, H.; Oliveira, M.B.P.P.; Martel, F.; Alves, R.C. Valorizing coffee silverskin based on its phytochemicals and antidiabetic potential: From lab to a pilot scale. Foods 2022, 11, 1671. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Prot. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Farah, A.; de Paulis, T.; Moreira, D.P.; Trugo, L.C.; Martin, P.R. Chlorogenic acids and lactones in regular and water-decaffeinated arabica coffees. J. Agric. Food Chem. 2006, 54, 374–381. [Google Scholar] [CrossRef]
- Habtemariam, S. Chemical and pharmacological evidences for coffee as a modulator of type 2 diabetes and metabolic syndrome. In Medicinal Foods as Potential Therapies for Type-2 Diabetes and Associated Diseases—The Chemical and Pharmacological Basis of Their Action; Habtemariam, S., Ed.; Academic Press: London, UK, 2019; pp. 793–838. [Google Scholar]
- Narita, Y.; Inouye, K. High antioxidant activity of coffee silverskin extracts obtained by the treatment of coffee silverskin with subcritical water. Food Chem. 2012, 135, 943–949. [Google Scholar] [CrossRef]
- Tunnicliffe, J.M.; Cowan, T.; Shearer, J. Chlorogenic acid in whole body and tissue-specific glucose regulation. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: London, UK; Elsevier Inc.: Waltham, MA, USA, 2015; pp. 777–785. [Google Scholar]
- Budryn, G.; Nebesny, E.; Podsędek, A.; Żyżelewicz, D.; Materska, M.; Jankowski, S.; Janda, B. Effect of different extraction methods on the recovery of chlorogenic acids, caffeine and Maillard reaction products in coffee beans. Eur. Food Res. Technol. 2009, 228, 913–922. [Google Scholar] [CrossRef]
- Pilipczuk, T.; Kusznierewicz, B.; Zielińska, D.; Bartoszek, A. The influence of roasting and additional processing on the content of bioactive components in special purpose coffees. J. Food Sci. Technol. 2014, 52, 5736–5744. [Google Scholar] [CrossRef] [PubMed]
- Tfouni, S.A.V.; Serrate, C.S.; Carreiro, L.B.; Camargo, M.C.R.; Teles, C.R.A.; Cipolli, K.M.V.A.B.; Furlani, R.P.Z. Effect of roasting on chlorogenic acids, caffeine and polycyclic aromatic hydrocarbons levels in two Coffea cultivars: Coffea arabica cv. Catuaí Amarelo IAC-62 and Coffea canephora cv. Apoatã IAC-2258. Int. J. Food Sci. Technol. 2012, 47, 406–415. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Sentkowska, A.; Pyrzyńska, K.; De Peña, M.P. Chlorogenic acids, caffeine content and antioxidant properties of green coffee extracts: Influence of green coffee bean preparation. Eur. Food Res. Technol. 2016, 242, 1403–1409. [Google Scholar] [CrossRef]
- Rostagno, M.A.; Celeghini, R.M.S.; Debien, I.C.N.; Nogueira, G.C.; Meireles, M.A.A. Phenolic compounds in coffee compared to other beverages. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: London, UK, 2015; pp. 137–142. [Google Scholar]
- Zamanipoor, M.H.; Yakufu, B.; Tse, E.; Rezaeimotlagh, A.; Hook, J.M.; Bucknall, M.P.; Thomas, D.S.; Trujillo, F.J. Brewing coffee?–ultra-sonication has clear beneficial effects on the extraction of key volatile aroma components and triglycerides. Ultrason. Sonochem. 2020, 60, 104796. [Google Scholar] [CrossRef] [PubMed]
- Panusa, A.; Petrucci, R.; Lavecchia, R.; Zuorro, A. UHPLC-PDA-ESI-TOF/MS metabolic profiling and antioxidant capacity of arabica and robusta coffee silverskin: Antioxidants vs phytotoxins. Food Res. Int. 2017, 99, 155–165. [Google Scholar] [CrossRef]
- Martinez-Saez, N.; Ullate, M.; Martin-Cabrejas, M.A.; Martorell, P.; Genovés, S.; Ramon, D.; del Castillo, M.D. A novel antioxidant beverage for body weight control based on coffee silverskin. Food Chem. 2014, 150, 227–234. [Google Scholar] [CrossRef]
- Bresciani, L.; Calani, L.; Bruni, R.; Brighenti, F.; Del Rio, D. Phenolic composition, caffeine content and antioxidant capacity of coffee silverskin. Food Res. Int. 2014, 61, 196–201. [Google Scholar] [CrossRef]
- Guarino, M.P.; Sacramento, J.; Ribeiro, M.J.; Conde, S.V. Caffeine, insulin resistance, and hypertension. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: London, UK, 2015; pp. 747–755. [Google Scholar]
- Pimentel, G.D.; Micheletti, T.O.; Fernandes, R.C.; Nehlig, A. Coffee intake and obesity. In Nutrition in the Prevention and Treatment of Abdominal Obesity, 2nd ed.; Watson, R., Ed.; Academic Press: London, UK; Elsevier Inc.: Waltham, MA, USA, 2019; pp. 329–351. [Google Scholar]
- Schuster, J.; Mitchell, E.S. More than just caffeine: Psychopharmacology of methylxanthine interactions with plant-derived phytochemicals. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 263–274. [Google Scholar] [CrossRef]
- Alves, R.C.; Soares, C.M.D.; Oliveira, M.B.P.P. Isoflavones in coffee. In Coffee in Health and Disease Prevention; Preedy, V.R., Ed.; Academic Press: London, UK; Elsevier Inc.: Waltham, MA, USA, 2015; pp. 143–148. [Google Scholar]
- Andrade, N.; Araújo, J.R.; Correia-Branco, A.; Carletti, J.V.; Martel, F. Effect of dietary polyphenols on fructose uptake by human intestinal epithelial (Caco-2) cells. J. Funct. Foods 2017, 36, 429–439. [Google Scholar] [CrossRef]
- Nikpayam, O.; Najafi, M.; Ghaffari, S.; Jafarabadi, M.A.; Sohrab, G.; Roshanravan, N. Effects of green coffee extract on fasting blood glucose, insulin concentration and homeostatic model assessment of insulin resistance (HOMA-IR): A systematic review and meta-analysis of interventional studies. Diabetol. Metab. Syndr. 2019, 11, 91. [Google Scholar] [CrossRef]
- Xu, M.; Yang, L.; Zhu, Y.; Liao, M.; Chu, L.; Li, X.; Lin, L.; Zheng, G. Collaborative effects of chlorogenic acid and caffeine on lipid metabolism via the AMPKα-LXRα/SREBP-1c pathway in high-fat diet-induced obese mice. Food Funct. 2019, 10, 7489–7497. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Green Coffee | Roasted Coffee | Coffee Silverskin | |
|---|---|---|---|
| Caffeine | 51.66 ± 0.92 b | 60.22 ± 0.47 a | 32.81 ± 1.21 c |
| 5-CQA | 129.82 ± 5.02 a | 26.57 ± 0.05 b | 3.45 ± 0.13 c |
| 4-CQA | 38.90 ± 2.09 a | 18.00 ± 0.11b b | 0.74 ± 0.03 c |
| 3-CQA | 24.53 ± 1.50 a | 13.78 ± 0.55 b | 0.70 ± 0.06 c |
| 5-FQA | 52.58 ± 2.77 a | 15.75 ± 0.06 b | 2.56 ± 0.10 c |
| 4-FQA | 7.24 ± 0.38 a | 5.39 ± 0.09 b | 0.64 ± 0.07 c |
| Ʃ CGA | 253.07 | 79.49 | 8.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, J.A.B.; Andrade, N.; Machado, S.; Costa, A.S.G.; Oliveira, M.B.P.P.; Martel, F.; Alves, R.C. Green/Roasted Coffee and Silverskin Extracts Inhibit Sugar Absorption by Human Intestinal Epithelial (Caco-2) Cells by Decreasing GLUT2 Gene Expression. Foods 2022, 11, 3902. https://doi.org/10.3390/foods11233902
Peixoto JAB, Andrade N, Machado S, Costa ASG, Oliveira MBPP, Martel F, Alves RC. Green/Roasted Coffee and Silverskin Extracts Inhibit Sugar Absorption by Human Intestinal Epithelial (Caco-2) Cells by Decreasing GLUT2 Gene Expression. Foods. 2022; 11(23):3902. https://doi.org/10.3390/foods11233902
Chicago/Turabian StylePeixoto, Juliana A. Barreto, Nelson Andrade, Susana Machado, Anabela S. G. Costa, Maria Beatriz P. P. Oliveira, Fátima Martel, and Rita C. Alves. 2022. "Green/Roasted Coffee and Silverskin Extracts Inhibit Sugar Absorption by Human Intestinal Epithelial (Caco-2) Cells by Decreasing GLUT2 Gene Expression" Foods 11, no. 23: 3902. https://doi.org/10.3390/foods11233902
APA StylePeixoto, J. A. B., Andrade, N., Machado, S., Costa, A. S. G., Oliveira, M. B. P. P., Martel, F., & Alves, R. C. (2022). Green/Roasted Coffee and Silverskin Extracts Inhibit Sugar Absorption by Human Intestinal Epithelial (Caco-2) Cells by Decreasing GLUT2 Gene Expression. Foods, 11(23), 3902. https://doi.org/10.3390/foods11233902