
Investigating the Probiotic Properties and Antimicrobial Activity of Lactic Acid Bacteria Isolated from an Iranian Fermented Dairy Product, Kashk

Abstract
1. Introduction
2. Materials and Methods
2.1. Screening of LAB
2.2. Assessment of Probiotic Properties
2.2.1. Cell Survival in Simulated Digestive System
2.2.2. Bile Salts Tolerance
2.2.3. Antibiotic Susceptibility
2.2.4. Antibacterial Activity Assay
2.2.5. Cell Surface Hydrophobicity
2.2.6. Auto-Aggregation Assay
2.3. Statistical Analysis
3. Results and Discussion
3.1. Identification of Isolates
3.2. Cell Survival in Low pH Conditions, Simulated Digestive System, and Bile Salts
3.3. Antibiotic Susceptibility
3.4. Antibacterial Activity
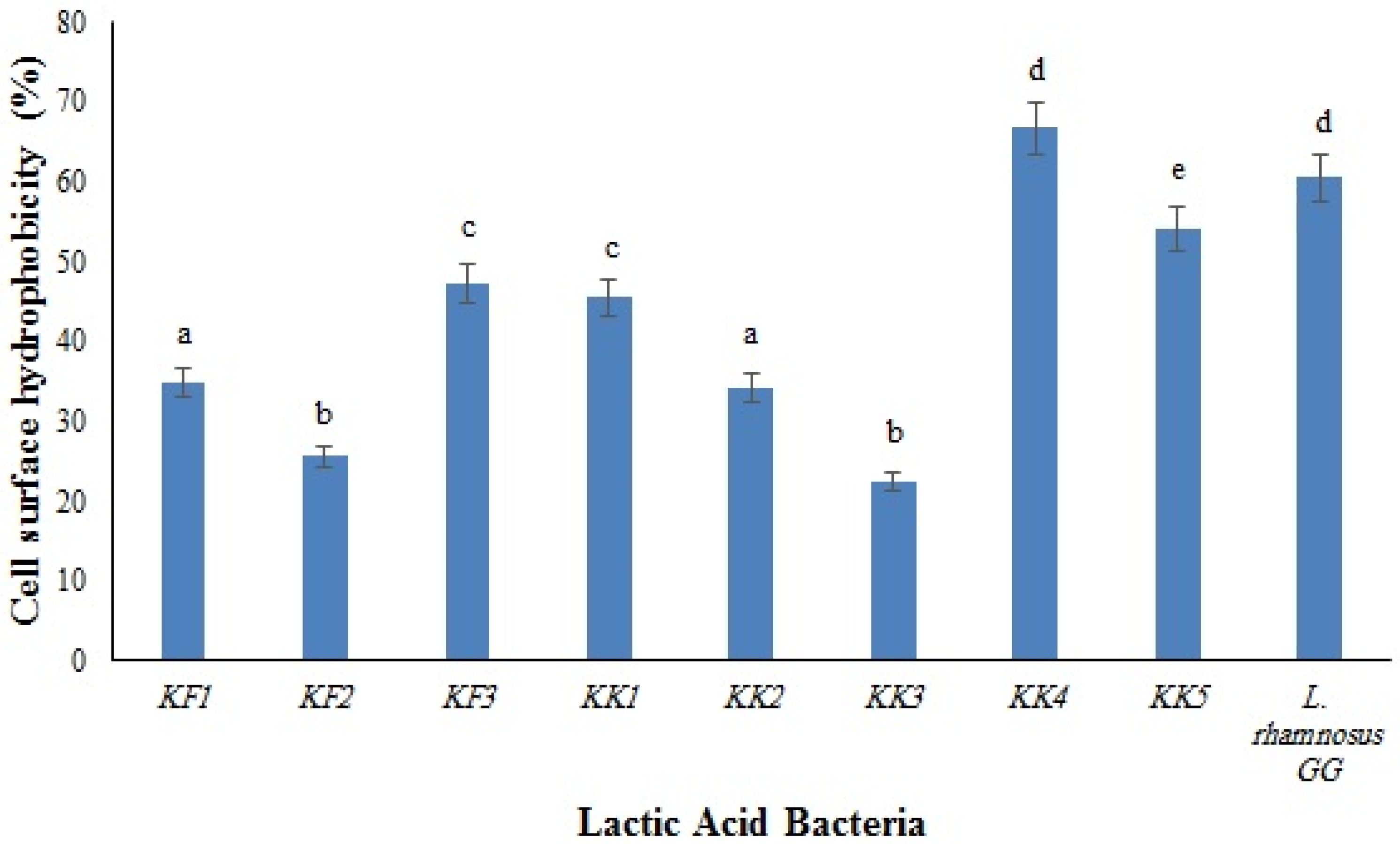
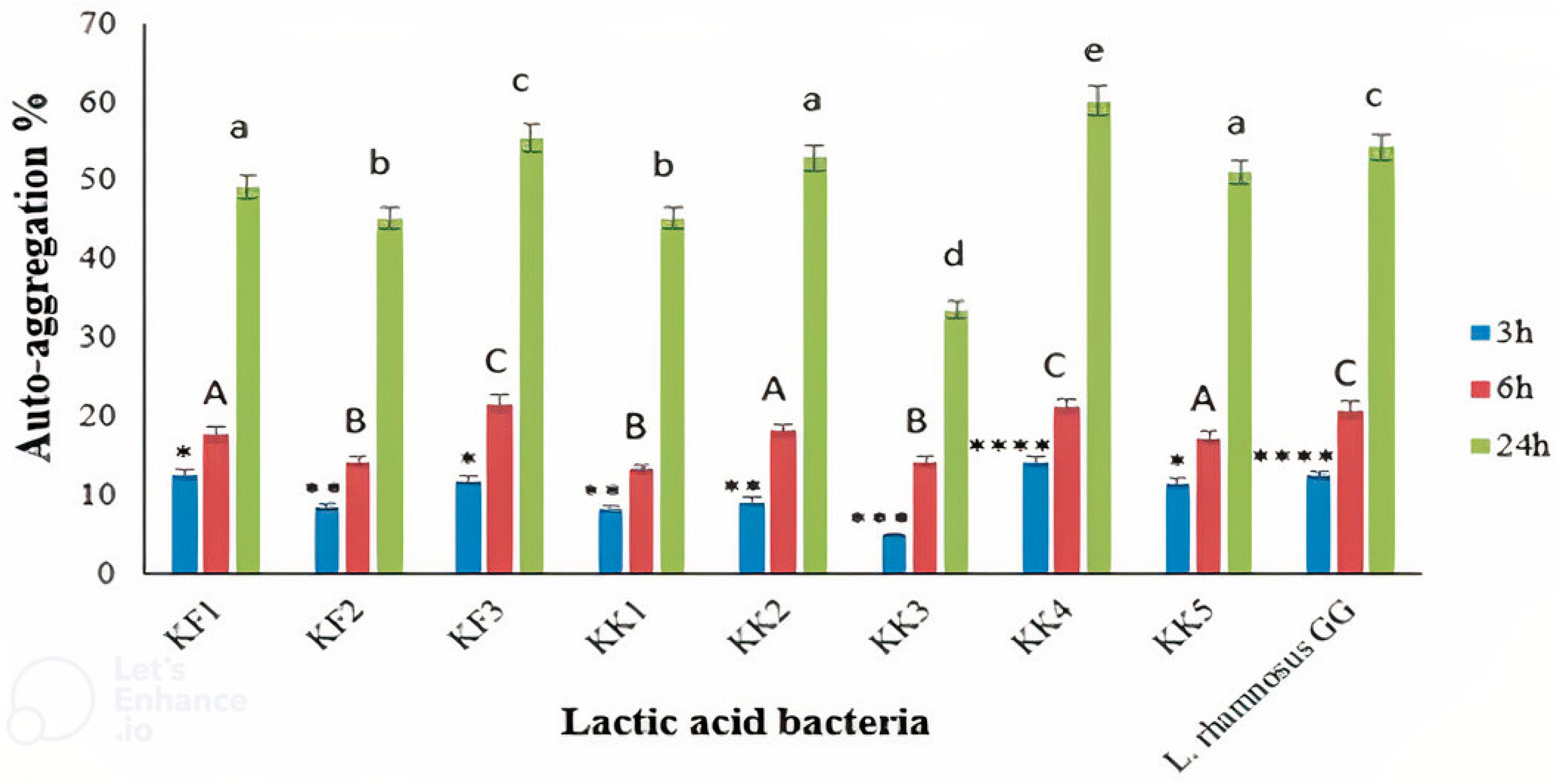
3.5. Cell Surface Hydrophobicity and Auto-Aggregation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soltani, M.; Güzeler, N. The production and quality properties of liquid kashks. Gida 2013, 38, 1–7. [Google Scholar]
- Jafari, M.; Rezaei, M.; Gheisari, H.R.; Abhari, K.; Khaniki, G.J.; Noori, N.; Khaneghah, A.M. Application of cultivable lactic acid bacteria isolated from Iranian traditional dairy products for the production of liquid and dried kashks. LWT 2019, 116, 108519. [Google Scholar] [CrossRef]
- Mohamad, N.; Manan, H.; Sallehhuddin, M.; Musa, N.; Ikhwanuddin, M. Screening of Lactic Acid Bacteria isolated from giant freshwater prawn (Macrobrachium rosenbergii) as potential probiotics. Aquac. Rep. 2020, 18, 100523. [Google Scholar] [CrossRef]
- Rahnama Vosough, P.; Habibi Najafi, M.B.; Edalatian Dovom, M.R.; Javadmanesh, A.; Mayo, B. Evaluation of antioxidant, antibacterial and cytotoxicity activities of exopolysaccharide from Enterococcus strains isolated from traditional Iranian Kishk. J. Food Meas. Charact. 2021, 15, 5221–5230. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K. Lactic acid fermentation of cereals and pseudocereals: Ancient nutritional biotechnologies with modern applications. Nutrients 2020, 12, 1118. [Google Scholar] [CrossRef]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: From starter cultures to producers of chemicals. FEMS Microbiol. Lett. 2018, 365, fny213. [Google Scholar] [CrossRef]
- Vosough, P.R.; Dovom, M.R.E.; Najafi, M.B.H.; Javadmanesh, A.; Mayo, B. Biodiversity of exopolysaccharide-producing lactic acid bacteria from Iranian traditional Kishk and optimization of EPS yield by Enterococcus spp. Food Biosci. 2022, 49, 101869. [Google Scholar] [CrossRef]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Reddy, M.S. Probiotics: Genesis, current definition, and proven therapeutic properties. Edw. Jenner FRS 2021, 1, 18. [Google Scholar]
- Vasiee, A.; Falah, F.; Sankian, M.; Tabatabaei-Yazdi, F.; Mortazavi, S.A. Oral immunotherapy using probiotic ice cream containing recombinant food-grade Lactococcus lactis which inhibited allergic responses in a BALB/c mouse model. J. Immunol. Res. 2020, 2020, 2635230. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Nam, D.G.; Kim, S.B.; Im, P.; Choe, J.S.; Choi, A.J. Enhancement of viability, acid, and bile tolerance and accelerated stability in lyophilized Weissella cibaria JW 15 with protective agents. Food Sci. Nutr. 2018, 6, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Zacarchenco, P.B.; Pimentel, T.C.; e Alves, A.T.S.; Spadoti, L.M.; Esmerino, E.A.; Silva, M.C.; da Cruz, A.G. Chapter 16—Health benefits of probiotics: An overview. In Advances in Dairy Microbial Products; Singh, J., Vyas, A., Eds.; Woodhead Publishing: Duxford, UK, 2022; pp. 235–245. ISBN 978-0-323-85793-2. [Google Scholar]
- Sanders, M.; Merenstein, D.; Merrifield, C.; Hutkins, R. Probiotics for human use. Nutr. Bull. 2018, 43, 212–225. [Google Scholar] [CrossRef]
- Vasiee, A.; Mortazavi, A.; Tabatabaei-yazdi, F.; Dovom, M. Detection, identification and phylogenetic analysis of lactic acid bacteria isolated from Tarkhineh, Iranian fermented cereal product, by amplifying the 16s rRNA gene with universal primers and differentiation using rep-PCR. Int. Food Res. J. 2018, 25, 423–432. [Google Scholar]
- Meem, F.C.; Raihan, T.; Shourove, J.H.; Azad, A.K.; Islam, G.R. Antibiotic resistance in Gram-negative bacteria isolated from street-vended foods in Bangladesh. bioRxiv 2022. [Google Scholar] [CrossRef]
- Edalatian, M.R.; Najafi, M.B.H.; Mortazavi, S.A.; Alegría, Á.; Nassiri, M.R.; Bassami, M.R.; Mayo, B. Microbial diversity of the traditional Iranian cheeses Lighvan and Koozeh, as revealed by polyphasic culturing and culture-independent approaches. Dairy Sci. Technol. 2012, 92, 75–90. [Google Scholar] [CrossRef]
- Vasiee, A.; Falah, F.; Behbahani, B.A.; Tabatabaee-Yazdi, F. Probiotic characterization of Pediococcus strains isolated from Iranian cereal-dairy fermented product: Interaction with pathogenic bacteria and the enteric cell line Caco-2. J. Biosci. Bioeng. 2020, 130, 471–479. [Google Scholar] [CrossRef]
- Vasiee, A.; Tabatabaei Yazdi, F.; Mortazavi, A.; Edalatian, M. Isolation, identification and characterization of probiotic Lactobacilli spp. from Tarkhineh. Int. Food Res. J. 2014, 21, 2487–2492. [Google Scholar]
- Nami, Y.; Vaseghi Bakhshayesh, R.; Mohammadzadeh Jalaly, H.; Lotfi, H.; Eslami, S.; Hejazi, M.A. Probiotic properties of Enterococcus isolated from artisanal dairy products. Front. Microbiol. 2019, 10, 300. [Google Scholar] [CrossRef]
- Reuben, R.; Roy, P.; Sarkar, S.; Alam, A.R.U.; Jahid, I. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Shahidi, F.; Mortazavi, S.A.; Milani, E.; Eshaghi, Z. Potentially probiotic Lactobacillus strains from traditional Kurdish cheese. Probiotics Antimicrob. Proteins 2014, 6, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Vasiee, A.; Mortazavi, S.A.; Sankian, M.; Yazdi, F.T.; Mahmoudi, M.; Shahidi, F. Antagonistic activity of recombinant Lactococcus lactis NZ1330 on the adhesion properties of Escherichia coli causing urinary tract infection. Microb. Pathog. 2019, 133, 103547. [Google Scholar] [CrossRef] [PubMed]
- Falah, F.; Vasiee, A.; Behbahani, B.A.; Yazdi, F.T.; Moradi, S.; Mortazavi, S.A.; Roshanak, S. Evaluation of adherence and anti-infective properties of probiotic Lactobacillus fermentum strain 4—17 against Escherichia coli causing urinary tract infection in humans. Microb. Pathog. 2019, 131, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Bachmann, H.; van Pelt-KleinJan, E.; Douwenga, S.; Smid, E.J.; Teusink, B.; van Mastrigt, O. Lifestyle, metabolism and environmental adaptation in Lactococcus lactis. FEMS Microbiol. Rev. 2020, 44, 804–820. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Settachaimongkon, S.; Nout, M.R.; Fernandes, E.C.A.; Hettinga, K.A.; Vervoort, J.M.; van Hooijdonk, T.C.; Zwietering, M.H.; Smid, E.J.; van Valenberg, H.J. Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp. bulgaricus on the metabolite profile of set-yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Plessas, S.; Nouska, C.; Karapetsas, A.; Kazakos, S.; Alexopoulos, A.; Mantzourani, I.; Chondrou, P.; Fournomiti, M.; Galanis, A.; Bezirtzoglou, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017, 226, 102–108. [Google Scholar] [CrossRef]
- Ding, W.; Shah, N. Acid, bile, and heat tolerance of free and microencapsulated probiotic bacteria. J. Food Sci. 2007, 72, M446–M450. [Google Scholar] [CrossRef]
- Parente, E.; Ciocia, F.; Ricciardi, A.; Zotta, T.; Felis, G.E.; Torriani, S. Diversity of stress tolerance in Lactobacillus plantarum, Lactobacillus pentosus and Lactobacillus paraplantarum: A multivariate screening study. Int. J. Food Microbiol. 2010, 144, 270–279. [Google Scholar] [CrossRef]
- Feyhl-Buska, J.; Chen, Y.; Jia, C.; Wang, J.-X.; Zhang, C.L.; Boyd, E.S. Influence of growth phase, pH, and temperature on the abundance and composition of tetraether lipids in the thermoacidophile Picrophilus torridus. Front. Microbiol. 2016, 7, 1323. [Google Scholar] [CrossRef] [PubMed]
- Azat, R.; Liu, Y.; Li, W.; Kayir, A.; Lin, D.-b.; Zhou, W.-w.; Zheng, X.-d. Probiotic properties of lactic acid bacteria isolated from traditionally fermented Xinjiang cheese. J. Zhejiang Univ. Sci. B 2016, 17, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Fani, A.; Singh, H. Biophysical insights into modulating lipid digestion in food emulsions. Prog. Lipid Res. 2022, 85, 101129. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, I.; Chondrou, P.; Bontsidis, C.; Karolidou, K.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Galanis, A.; Plessas, S. Assessment of the probiotic potential of lactic acid bacteria isolated from kefir grains: Evaluation of adhesion and antiproliferative properties in in vitro experimental systems. Ann. Microbiol. 2019, 69, 751–763. [Google Scholar] [CrossRef]
- Lashani, E.; Davoodabadi, A.; Dallal, M.M.S. Some probiotic properties of Lactobacillus species isolated from honey and their antimicrobial activity against foodborne pathogens. Vet. Res. Forum. 2020, 11, 121–126. [Google Scholar]
- EFSA. Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 2007, 5, 587. [Google Scholar]
- Gueimonde, M.; Sánchez, B.; de los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Gupta, A.; Sharma, N. Characterization of potential probiotic lactic acid bacteria-Pediococcus acidilactici Ch-2 isolated from Chuli-A traditional apricot product of Himalayan region for the production of novel bioactive compounds with special therapeutic properties. J. Food Microbiol. Saf. Hyg 2017, 2, 119. [Google Scholar] [CrossRef]
- Aditya, A.; Peng, M.; Young, A.; Biswas, D. Antagonistic mechanism of metabolites produced by Lactobacillus casei on lysis of enterohemorrhagic Escherichia coli. Front. Microbiol. 2020, 11, 574422. [Google Scholar] [CrossRef]
- Chaudhary, A.; Saharan, B.S. Probiotic Properties of Lactobacillus plantarum. J. Pure Appl. Microbiol. 2019, 13, 933–949. [Google Scholar] [CrossRef]
- Saboktakin-Rizi, M.; Alizadeh Behbahani, B.; Hojjati, M.; Noshad, M. Identification of Lactobacillus plantarum TW29-1 isolated from Iranian fermented cereal-dairy product (Yellow Zabol Kashk): Probiotic characteristics, antimicrobial activity and safety evaluation. J. Food Meas. Charact. 2021, 15, 2615–2624. [Google Scholar] [CrossRef]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum sensing mediated pathogenicity in Pseudomonas aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Gharbi, Y.; Fhoula, I.; Ruas-Madiedo, P.; Afef, N.; Boudabous, A.; Gueimonde, M.; Ouzari, H.-I. In-vitro characterization of potentially probiotic Lactobacillus strains isolated from human microbiota: Interaction with pathogenic bacteria and the enteric cell line HT29. Ann. Microbiol. 2019, 69, 61–72. [Google Scholar] [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Raeisi, M.; Nematollahi, Z. Biological control of foodborne pathogens and aflatoxins by selected probiotic LAB isolated from rice bran sourdough. Biol. Control 2019, 130, 70–79. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group Number | Region | Number of Isolates | Growth at 10 °C | Growth at 45 °C | Growth at pH 4.4 | Growth at pH 9.6 | Growth at 6.5% NaCl | Arginine Hydrolysis | CO2 Gas Production |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Abadeh | 1 | + | - | + | ± | + | - | - |
| 2 | Abadeh | 2 | + | ± | ± | - | + | - | - |
| 3 | Kalat | 1 | + | - | - | - | - | + | - |
| 4 | Kalat | 4 | + | + | + | + | + | + | - |
| Sampling Location | Isolate Code | Highest Similarity in NCBI Database | Similarity Percentage | Accession Number |
|---|---|---|---|---|
| Abadeh | KF1 | Enterococcus faecium KKP 3772 | 99 | OL454909 |
| Abadeh | KF2 | Enterococcus faecium C1 | 100 | EU428011 |
| Abadeh | KF3 | Pediococcus pentosaceus H11 | 100 | OM388463 |
| Kalat | KK1 | Lactococcus lactis RSg | 97 | KJ660075 |
| Kalat | KK2 | Enterococcus faecalis P190052 | 98 | MN749533 |
| Kalat | KK3 | Enterococcus mundtii CECT972T | 100 | AJ420806 |
| Kalat | KK4 | Pediococcus pentosaceus VNK-1 | 99 | ON810480 |
| Kalat | KK5 | Lactiplantibacillus plantarum PM411 | 98 | JX440377 |
| pH 2 | pH 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain Code | 0 h | 1 h | 2 h | 3 h | 0 h | 1 h | 2 h | 3 h | Survival in GIT |
| Enterococcus faecium KKP 3772 (KF1) | 9.05 ± 0.11 | 7.06 ± 0.20 | 6.15 ± 0.14 | 5.15 ± 0.05 | 9.11 ± 0.08 | 8.90 ± 0.17 | 9.25 ± 0.60 | 9.31 ± 0.22 | 6.28 ± 0.25 |
| Enterococcus faecium C1 (KF2) | 9.14 ± 0.23 | 6.95 ± 0.70 | 6.40 ± 0.15 | 4.65 ± 0.18 | 9.03 ± 0.10 | 9.45 ± 0.28 | 9.89 ± 0.26 | 9.46 ± 0.11 | 6.55 ± 0.13 |
| Pediococcus pentosaceus H11 (KF3) | 9.02 ± 0.14 | 7.22 ± 0.12 | 7.35 ± 0.41 | 6.15 ± 0.23 | 9.12 ± 0.16 | 9.32 ± 0.14 | 9.50 ± 0.13 | 9.54 ± 0.18 | 7.35 ± 0.11 |
| Lactococcus lactis RSg (KK1) | 9.12 ± 0.07 | 6.95 ± 0.32 | 5.65 ± 0.23 | 3.95 ± 0.12 | 9.09 ± 0.18 | 8.75 ± 0.16 | 8.15 ± 0.10 | 7.70 ± 0.39 | 5.45 ± 0.37 |
| Enterococcus faecalis P190052 (KK2) | 9.06 ± 0.19 | 7.02 ± 0.33 | 6.65 ± 0.12 | 6.42 ± 0.14 | 9.10 ± 0.13 | 9.51 ± 0.37 | 9.36 ± 0.10 | 9.30 ± 0.41 | 6.80 ± 0.16 |
| Enterococcus mundtii CECT972T (KK3) | 9.13 ± 0.21 | 7.89 ± 0.22 | 6.28 ± 0.15 | 5.67 ± 0.19 | 9.07 ± 0.12 | 9.20 ± 0.13 | 9.28 ± 0.34 | 9.18 ± 0.15 | 6.45 ± 0.36 |
| Pediococcus pentosaceus VNK-1 (KK4) | 9.03 ± 0.12 | 8.26 ± 0.19 | 7.75 ± 0.16 | 6.56 ± 0.35 | 9.05 ± 0.14 | 9.11 ± 0.23 | 9.48 ± 0.36 | 9.67 ± 0.41 | 7.52 ± 0.15 |
| Lactiplantibacillus plantarum PM411 (KK5) | 8.98 ± 0.31 | 7.23 ± 0.16 | 6.35 ± 0.20 | 5.71 ± 0.25 | 9.03 ± 0.15 | 9.10 ± 0.42 | 8.95 ± 0.35 | 8.80 ± 0.09 | 5.95 ± 0.20 |
| Strain Code | Bile Acid Degradation Ability | Growth in the Presence of Bile (%, Compared to the Control Sample) | |||||
|---|---|---|---|---|---|---|---|
| GDC * | GC | TDC | TC | 1 | 0.5 | 0.3 | |
| KF1 | g | g | g | g | 7.60 ± 0.30 | 11.2 ± 0.42 | 28.50 ± 0.25 |
| KF2 | - ** | - | - | wg | 0 | 0 | 8.1 ± 0.47 |
| KF3 | - | g | + | + | 65.04 ± 0.21 | 81.0 ± 0.27 | 84.60 ± 0.33 |
| KK1 | g | g | g | g | 0 | 9.10 ± 0.34 | 17.20 ± 0.18 |
| KK2 | + | g | + | g | 25.50 ± 0.30 | 41.40 ± 0.32 | 50.40 ± 0.47 |
| KK3 | - | wg | - | - | 0 | 0 | 7.90 ± 0.14 |
| KK4 | wg | g | + | + | 70.70 ± 0.19 | 82.40 ± 0.30 | 91.10 ± 0.42 |
| KK5 | - | g | g | g | 30.20 ± 0.46 | 41.08 ± 0.36 | 63.02 ± 0.25 |
| Strain Code | CN * | C | CRO | CC | GM | AM | E | TE | CP | K | SXT | CFM | FEB | S | CL | V |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KF1 | R | S | S | R | S | S | S | R | R | S | S | S | R | R | R | S |
| KF2 | R | S | I | R | S | R | S | R | R | S | R | R | S | R | R | S |
| KF3 | I | S | S | I | S | S | S | R | I | S | S | I | R | S | S | S |
| KK1 | S | S | S | R | S | I | S | R | I | S | S | S | I | S | I | S |
| KK2 | R | S | R | R | S | R | I | R | R | I | S | S | R | R | R | S |
| KK3 | S | S | S | S | S | S | S | R | S | S | S | S | S | S | R | S |
| KK4 | I | S | I | R | S | I | S | R | S | I | S | S | R | I | S | I |
| KK5 | R | S | R | S | S | R | I | R | R | S | I | I | R | R | R | I |
| Treatment | Indicator Pathogens | LAB | |||||||
|---|---|---|---|---|---|---|---|---|---|
| KF1 | KF2 | KF3 | KK1 | KK2 | KK3 | KK4 | KK5 | ||
| Escherichia coli ATCC 25922 | + * | ++ | ++ | ++ | ++ | ++ | +++ | +++ | |
| Not treated | Staphylococcus aureus ATCC 25923 | +++ | +++ | +++ | ++ | +++ | ++ | +++ | +++ |
| Pseudomonas aeruginosa PTCC 1707 | + | ++ | ++ | + | ++ | + | +++ | ++ | |
| Salmonella typhimurium ATCC 14028 | ++ | ++ | +++ | ++ | ++ | ++ | +++ | ++ | |
| Escherichia coli ATCC 25922 | - | - | + | - | - | - | + | + | |
| Neutralized | Staphylococcus aureus ATCC 25923 | - | - | + | - | - | - | + | + |
| Pseudomonas aeruginosa PTCC 1707 | - | - | - | - | - | - | + | - | |
| Salmonella typhimurium ATCC 14028 | + | - | + | - | - | - | + | + | |
| Escherichia coli ATCC 25922 | ++ | + | ++ | + | ++ | + | +++ | +++ | |
| Staphylococcus aureus ATCC 25923 | +++ | +++ | +++ | ++ | +++ | ++ | +++ | +++ | |
| Heat treatment | Pseudomonas aeruginosa PTCC 1707 | + | ++ | ++ | + | ++ | - | +++ | ++ |
| Salmonella typhimurium ATCC 14028 | ++ | ++ | +++ | ++ | ++ | ++ | +++ | ++ | |
| Escherichia coli ATCC 25922 | ++ | ++ | ++ | ++ | ++ | ++ | +++ | + | |
| Staphylococcus aureus ATCC 25923 | +++ | +++ | +++ | ++ | +++ | ++ | +++ | ++ | |
| Catalase treatment | Pseudomonas aeruginosa PTCC 1707 | + | ++ | ++ | + | ++ | + | +++ | - |
| Salmonella typhimurium ATCC 14028 | ++ | ++ | +++ | ++ | ++ | ++ | +++ | +++ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saboori, B.; Shahidi, F.; Hedayati, S.; Javadmanesh, A. Investigating the Probiotic Properties and Antimicrobial Activity of Lactic Acid Bacteria Isolated from an Iranian Fermented Dairy Product, Kashk. Foods 2022, 11, 3904. https://doi.org/10.3390/foods11233904
Saboori B, Shahidi F, Hedayati S, Javadmanesh A. Investigating the Probiotic Properties and Antimicrobial Activity of Lactic Acid Bacteria Isolated from an Iranian Fermented Dairy Product, Kashk. Foods. 2022; 11(23):3904. https://doi.org/10.3390/foods11233904
Chicago/Turabian StyleSaboori, Bahareh, Fakhri Shahidi, Sara Hedayati, and Ali Javadmanesh. 2022. "Investigating the Probiotic Properties and Antimicrobial Activity of Lactic Acid Bacteria Isolated from an Iranian Fermented Dairy Product, Kashk" Foods 11, no. 23: 3904. https://doi.org/10.3390/foods11233904
APA StyleSaboori, B., Shahidi, F., Hedayati, S., & Javadmanesh, A. (2022). Investigating the Probiotic Properties and Antimicrobial Activity of Lactic Acid Bacteria Isolated from an Iranian Fermented Dairy Product, Kashk. Foods, 11(23), 3904. https://doi.org/10.3390/foods11233904