
Fermentation of cv. Kalamata Natural Black Olives with Potential Multifunctional Yeast Starters



Abstract
1. Introduction
2. Materials and Methods
2.1. Olive Samples and Fermentation Procedures
2.2. Inoculum Preparation
2.3. Microbiological Analyses
2.4. Yeast Inoculation and DNA Amplification
2.5. Physicochemical Analyses
2.6. Data Analysis
3. Results and Discussion
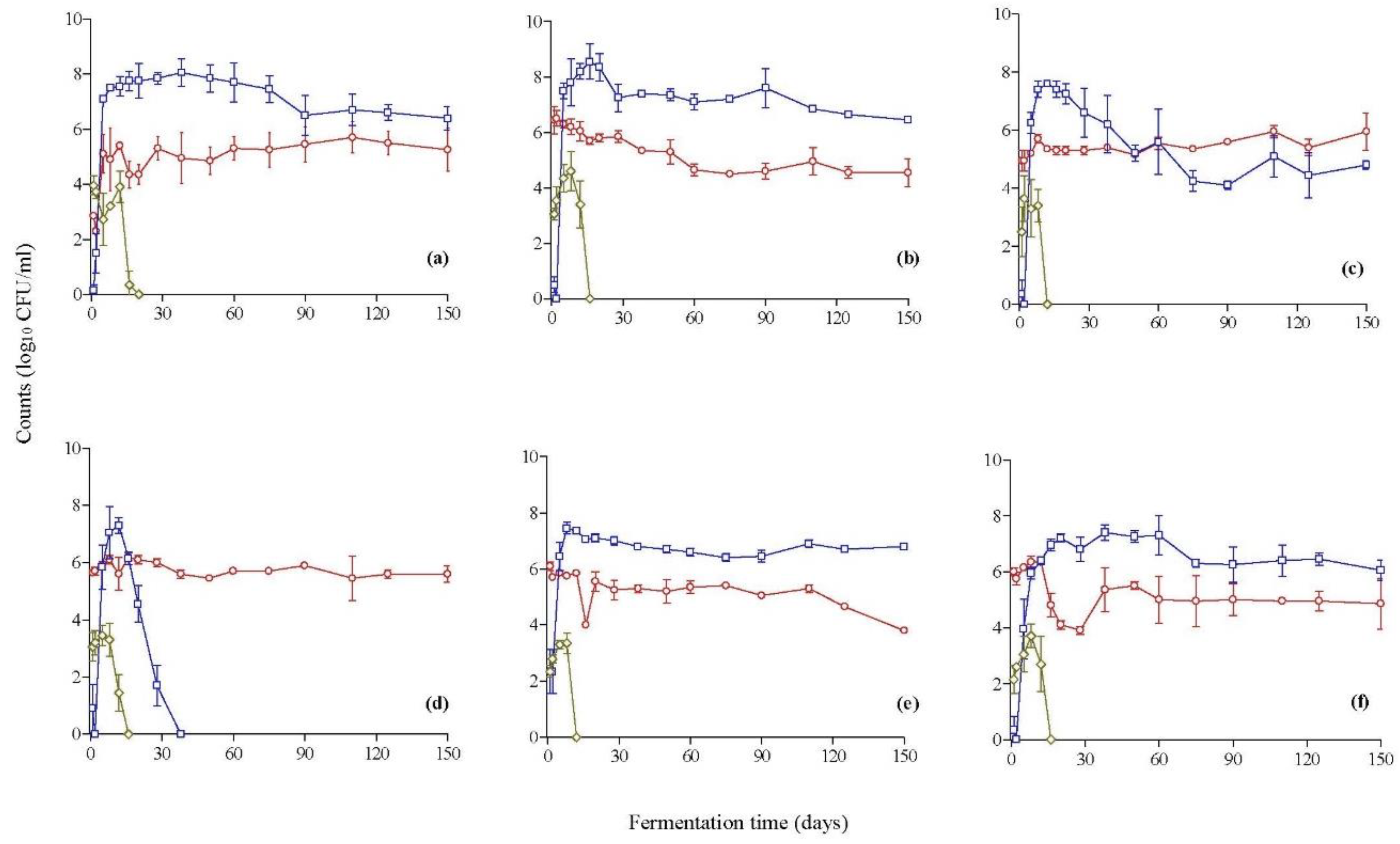
3.1. Population Dynamics during Fermentation
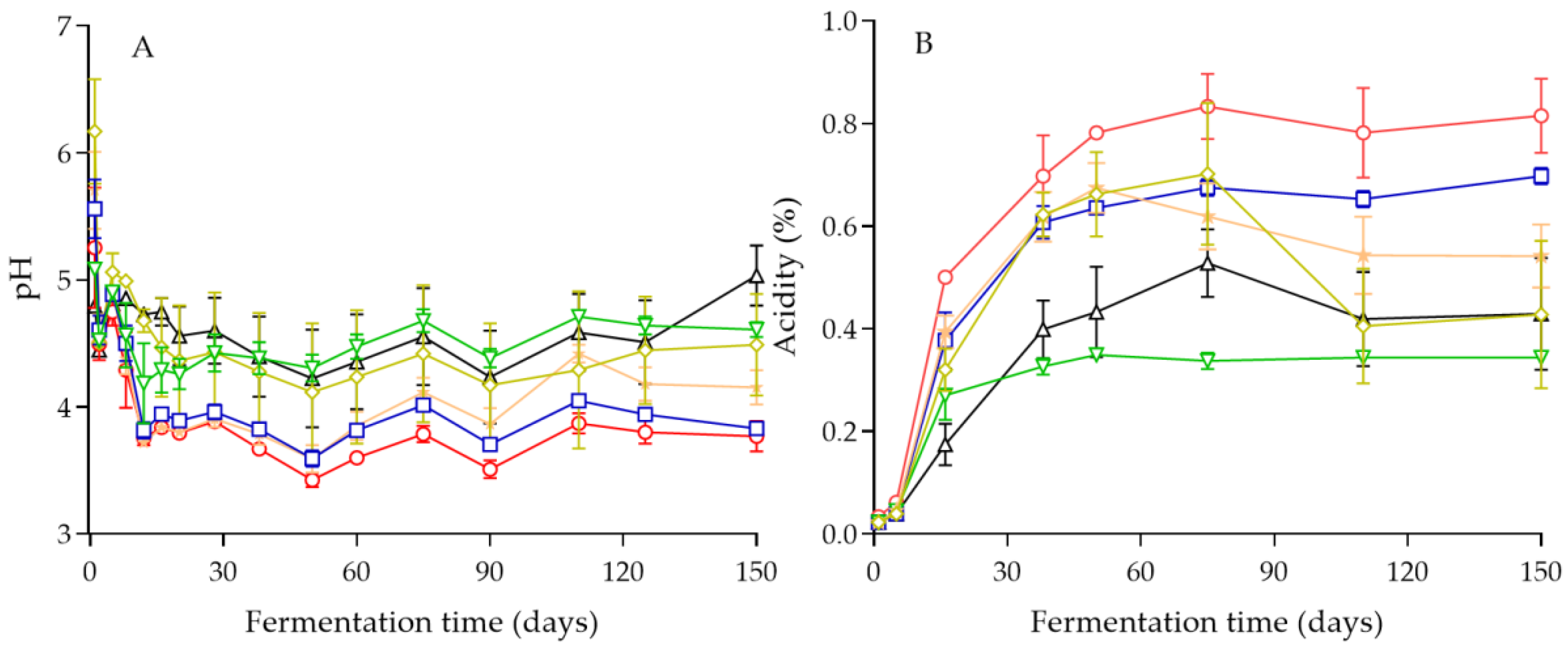
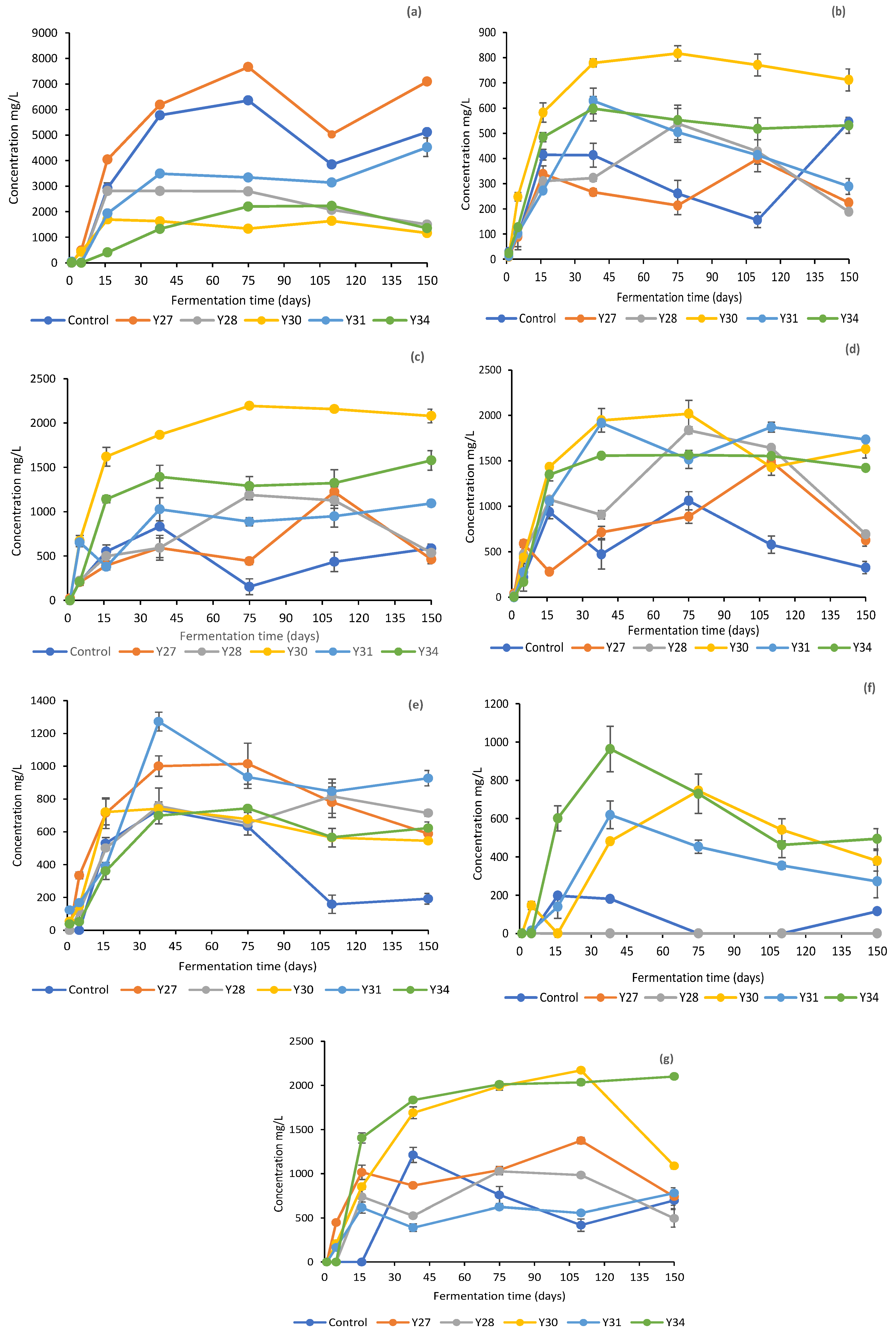
3.2. Physicochemical Changes in the Brines during Fermentation
3.3. Survival of the Inoculated Yeast Strains
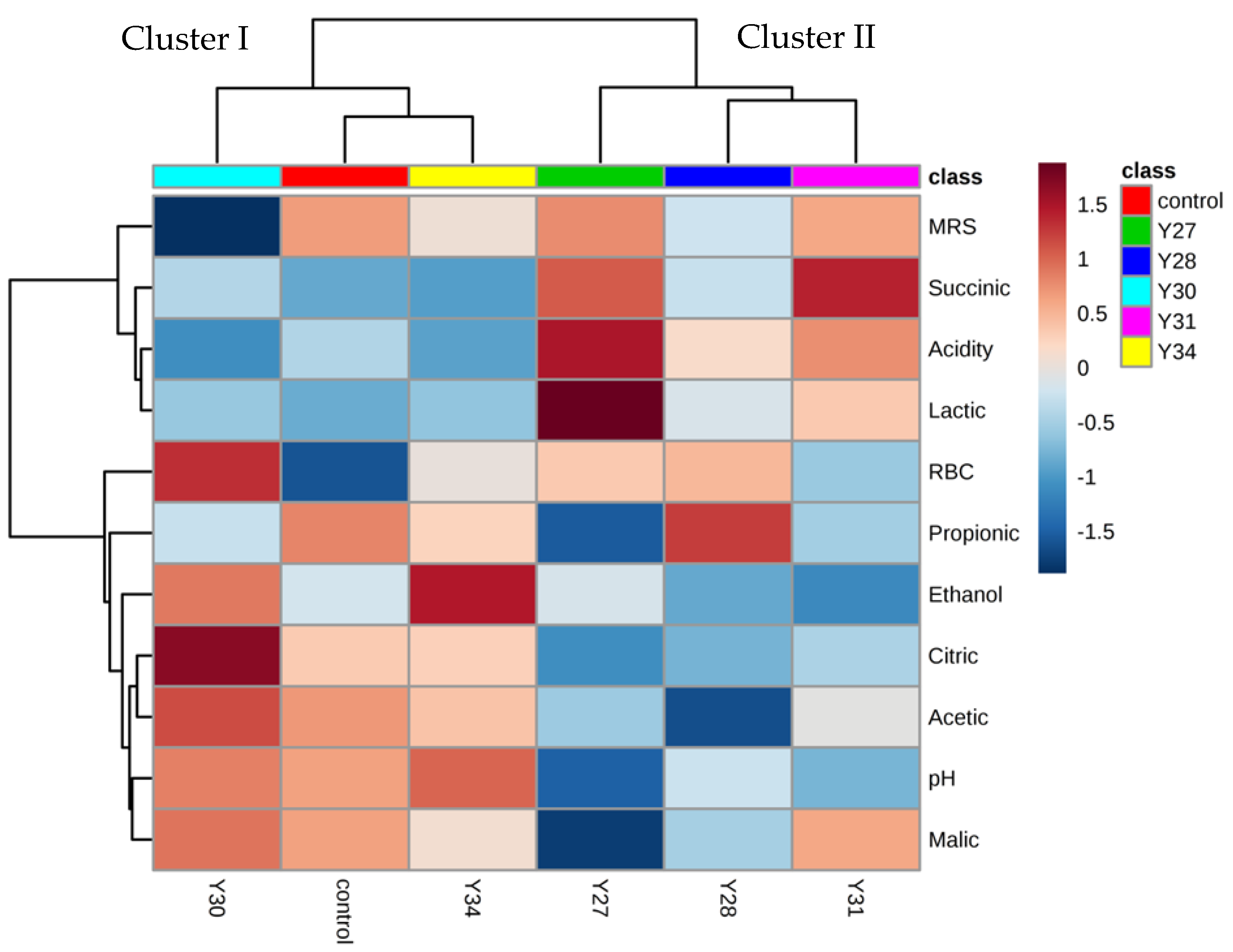
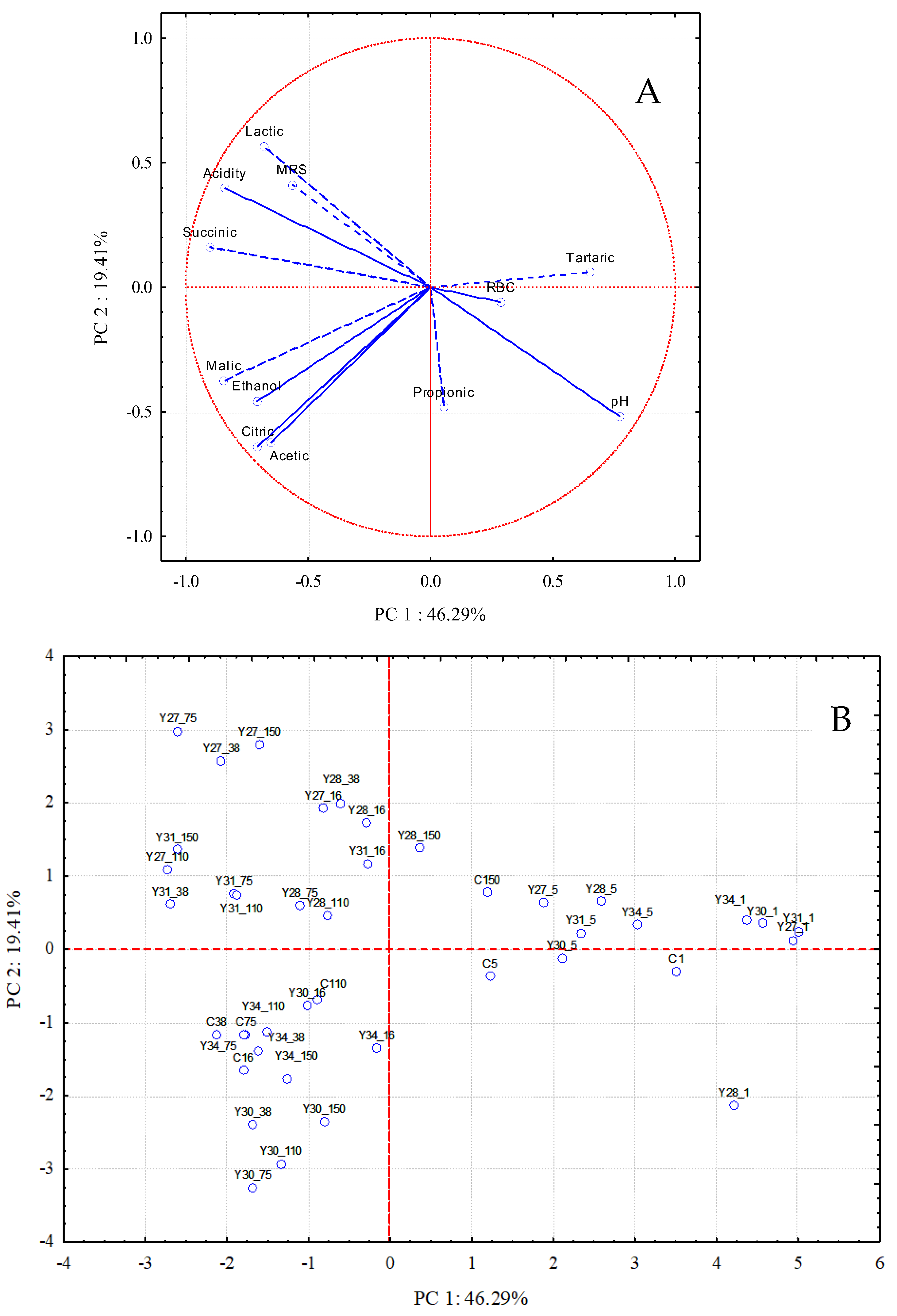
3.4. Exploratory Data Analysis of the Fermentation Profiles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Heperkan, D.; Dazkir, G.S.; Kansu, D.Z.; Güler, F.K. Influence of temperature on citrinin accumulation by Penicillium citrinum and Penicillium verrucosum in black table olives. Toxin Rev. 2009, 28, 180–186. [Google Scholar] [CrossRef]
- Conte, P.; Fadda, C.; Del Caro, A.; Urgeghe, P.P.; Piga, A. Table olives: An overview on effects of processing on nutritional and sensory quality. Foods 2020, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table olives more than a fermented food. Foods 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Ciafardini, G.; Zullo, B.A. Use of selected yeast starter cultures in industrial-scale processing of brined Taggiasca black table olives. Food Microbiol. 2019, 84, 130250. [Google Scholar] [CrossRef] [PubMed]
- Chytiri, A.; Tasioula-Margari, M.; Bleve, G.; Kontogianni, V.G.; Kallimanis, A.; Kontominas, M.G. Effect of different inoculation strategies of selected yeast and LAB cultures on Conservolea and Kalamàta table olives considering phenol content, texture, and sensory attributes. J. Sci. Food Agric. 2020, 100, 926–935. [Google Scholar] [CrossRef]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.J.N. Table olive fermentation using starter cultures with multifunctional potential microorganisms. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef]
- Anagnostopoulos, A.D.; Tsaltas, D. Current status, recent advances, and main challenges on table olive fermentation: The present meets the future. Front. Microbiol. 2022, 12, 797295. [Google Scholar] [CrossRef]
- Tufariello, M.; Durante, M.; Ramires, F.A.; Grieco, F.; Tommasi, L.; Perbellini, E.; Falco, V.; Tasioula-Margari, M.; Logrieco, A.F.; Mita, G.; et al. New process for production of fermented black table olives using selected autochthonous microbial resources. Front. Microbiol. 2015, 6, 1007–1022. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil1, V.; Arroyo-López, F.N.; Roldán-Reyes, J.C.; Torres-Gallardo, R.; Bautista-Gallego1, J.; García-García, P.; Garrido-Fernández, A. Assessing the challenges in the application of potential probiotic lactic acid bacteria in the large-scale fermentation of Spanish-style table olives. Front. Microbiol. 2017, 8, 915. [Google Scholar] [CrossRef]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 1–6. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Evaluation of a single and combined inoculation of a Lactobacillus pentosus starter for processing cv. Arbequina natural green olives. Food Microbiol. 2010, 27, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, C.L.; Rajendram, R.; Caggia, C. Lactic acid bacteria in table olive fermentations. In Olives and Olive Oil in Health and Disease Prevention, 2nd ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press: London, UK, 2010; pp. 369–376. [Google Scholar]
- Panagou, E.Z.; Schillinger, U.; Franz, C.M.A.P.; Nychas, G.J.E. Microbiological and biochemical profile of cv. Conservolea naturally black olives during controlled fermentation with selected strains of lactic acid bacteria. Food Microbiol. 2008, 25, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Barba, J.L.; Brenes-Balbuena, M.; Jiménez-Díaz, R.; García-García, P.; Garrido Fernández, A. Inhibition of Lactobacillus plantarum by polyphenols extracted from two different kinds of olive brine. J. Appl. Bacteriol. 1993, 74, 15–19. [Google Scholar] [CrossRef]
- Bonatsou, S.; Paramithiotis, S.; Panagou, E.Z. Evolution of yeast consortia during the fermentation of Kalamata natural black olives upon two initial acidification treatments. Front. Microbiol. 2017, 8, 2673. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; Garcia-Garcia, P.; Querol, A.; Garrido-Fernández, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 3, 161. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L. Use of selected autochthonous lactic acid bacteria for Spanish style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Querol, A.; Bautista-Gallego, J.; Garrido-Fernández, A. Role of yeasts in table olive production. Int. J. Food Microbiol. 2008, 128, 189–196. [Google Scholar] [CrossRef]
- D’Antuono, I.; Kontogianni, V.G.; Kotsiou, K.; Linsalata, V.; Logrieco, A.F.; Tasioula-Margari, M.; Cardinali, A. Polyphenolic characterization of olive mill wastewaters, coming from Italian and Greek olive cultivars, after membrane technology. Food Res. Int. 2014, 65, 301–310. [Google Scholar] [CrossRef]
- Porru, C.; Rodríguez-Gómez, F.; Benítez-Cabello, A.; Jiménez-Díaz, R.; Zara, G.; Budroni, M.; Mannazzu, I.; Arroyo-López, F.N. Genotyping, identification and multifunctional features of yeasts associated to Bosana naturally black table olive fermentations. Food Microbiol. 2018, 69, 33–42. [Google Scholar] [CrossRef]
- Bleve, G.; Tufariello, M.; Ramires, F.A.; Mita, G.; Tasioula-Margari, M.; Logrieco, A.F. Physicochemical process of Corservolea and Kalamata table olives and development of a protocol for the pre-selection of fermentation starters. Food Microbiol. 2015, 46, 368–382. [Google Scholar] [CrossRef]
- Bonatsou, S.; Benítez, A.; Rodríguez-Gómez, F.; Panagou, E.Z.; Arroyo-López, F.N. Selection of yeasts with multifunctional features for application as starters in natural black table olive processing. Food Microbiol. 2015, 46, 66–73. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Campanella, D.; Cosmai, L.; Summo, C.; Rizzello, C.G.; Caponio, F. Microbiota and metabolome of un-started and started Greek-type fermentation of Bella di Cerignola table olives. Food Microbiol. 2015, 52, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Anglana, C.; Crupi, P.; Virtuosi, I.; Fiume, P.; Di Terlizzi, B.; Moselhy, N.; Attay, H.; Pati, S.; Logrieco, A.F.; et al. Efficacy of yeast starters to drive and improve Picual, Manzanilla and Kalamata table olive fermentation. J. Sci. Food Agric. 2019, 99, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Selection of yeasts as starter cultures for table olives: A step-by-step procedure. Front. Microbiol. 2012, 3, 194. [Google Scholar] [CrossRef]
- Silva, T.; Reto, M.; Sol, M.; Peito, A.; Peres, C.M.; Peres, C.; Xavier Malcata, F. Characterization of yeasts from Portuguese brined olives, with a focus on their potentially probiotic behavior. LWT Food Sci. Technol. 2011, 44, 1349–1354. [Google Scholar] [CrossRef]
- Benítez-Cabello, A.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garrido-Fernández, A.; Jiménez-Díaz, R.; Arroyo-López, F.N. Evaluation and identification of poly-microbial biofilms on natural green Gordal table olives. Anton. Leeuw. 2015, 108, 597–610. [Google Scholar] [CrossRef]
- Pereira, L.E.; Ramalhosa, E.; Borges, A.; Pereira, J.A.; Baptista, P. Yeast dynamics during the natural fermentation process of table olives (Negrinha de Freixo cv.). Food Microbiol. 2015, 46, 582–586. [Google Scholar] [CrossRef]
- Alves, M.; Gonçalves, T.; Quintas, T. Microbial quality and yeast population dynamics in cracked green table olives’ fermentations. Food Cont. 2012, 23, 363–368. [Google Scholar] [CrossRef]
- Botta, C.; Cocolin, L. Microbial dynamics and biodiversity in table olive fermentation: Culture-dependent and -independent approaches. Front. Microbiol. 2012, 3, 245. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Rodríguez-Gómez, F.; Barrio, E.; Querol, A.; Garrido-Fernández, A.; Arroyo-López, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microbiol. 2011, 147, 89–96. [Google Scholar] [CrossRef]
- Nisiotou, A.A.; Chorianopoulos, N.; Nychas, G.J.E.; Panagou, E.Z. Yeast heterogeneity during spontaneous fermentation of black Conservolea olives in different brine solutions. J. Appl. Microbiol. 2010, 108, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Reguant, C.; Esteve-Zarzoso, B.; Bordons, A.; Rozès, N. Microbial population dynamics during the processing of Aberquina table olives. Food Res. Int. 2008, 41, 738–744. [Google Scholar] [CrossRef]
- Coton, E.; Coton, M.; Levert, D.; Casaregola, S.; Sohier, D. Yeast ecology in Frech cider and black olive natural fermentations. Int. J. Food Microbiol. 2006, 108, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gómez, F.; Arroyo-López, F.N.; López-López, A.; Bautista-Gallego, J.; Garrido-Fernández, A. Lipolytic activity of the yeast species associated with the fermentation/storage phase of ripe olive processing. Food Microbiol. 2010, 27, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Martín, A.; Córdoba, M.G.; Benito, M.J.; Aranda, E.; Pérez-Nevado, F. Determination of killer activity in yeasts isolated from the elaboration of seasoned green table olives. Int. J. Food Microbiol. 2008, 121, 178–188. [Google Scholar] [CrossRef]
- Psani, M.; Kotzekidou, P. Technological characteristics of yeast strains and their potential as starter adjuncts in Greek-style black olive fermentation. World J. Microbiol. Biotechnol. 2006, 22, 1329–1336. [Google Scholar] [CrossRef]
- Hernández, A.; Martín, A.; Aranda, E.; Pérez-Nevado, F.; Córdoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–351. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Bautista-Gallego, J.; Garrido-Fernández, A.; Arroyo-López, F.N. Multivariate analysis to discriminate yeast strains with technological applications in table olive processing. World J. Microbiol. Biotechnol. 2012, 28, 1761–1770. [Google Scholar] [CrossRef]
- Blana, V.A.; Grounta, A.; Tassou, C.C.; Nychas, G.J.E.; Panagou, E.Z. Inoculated fermentation of green olives with potential probiotic Lactobacillus pentosus and Lactobacillus plantarum starter cultures isolated from industrially fermented olives. Food Microbiol. 2014, 38, 208–218. [Google Scholar] [CrossRef]
- Harrigan, W.F. Laboratory Methods in Food Microbiology, 3rd ed.; Academic Press: London, UK, 1998. [Google Scholar]
- Garrido Fernández, A.; Fernández Díez, M.J.; Adams, M.R. Table Olives. Production and Processing; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, 251–257. [Google Scholar] [CrossRef]
- Abbas, C. Production of antioxidants, aromas, colours, flavours and vitamins. In Yeasts in Food and Beverages; Querol, A., Fleet, G., Eds.; Springer: New York, NY, USA, 2006; pp. 285–334. [Google Scholar]
- Pistarino, E.; Aliakbarian, B.; Casazza, A.A.; Paini, M.; Cosulich, M.E. Combined effect of starter culture and temperature on phenolic compounds during fermentation of Taggiasca black olives. Food Chem. 2013, 138, 2043–2049. [Google Scholar] [CrossRef] [PubMed]
- Grounta, A.; Doulgeraki, A.I.; Nychas, G.J.E.; Panagou, E.Z. Biofilm formation on Conservolea natural black olives during single and combined inoculation with a functional Lactobacillus pentosus culture. Food Microbiol. 2016, 56, 35–44. [Google Scholar] [CrossRef] [PubMed]
- International Olive Council (IOC). Quality Management Guide for the Table Olive Industry. T.OT/Doc. No. 14, Madrid, Spain. 2005. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2019/11/T.OT-Doc.14-2005-Eng.pdf (accessed on 1 September 2022).
- International Olive Council (IOC). Trade Standard Applying to Table Olives. DOI/OT/NC no. 1, Madrid, Spain. 2004. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2019/11/COI-OT-NC1-2004-Eng.pdf (accessed on 1 September 2022).
- Bobillo, M.; Marshall, V.M. Effect of salt and culture aeration on lactate and acetate production by Lactobacillus plantarum. Food Microbiol. 1991, 8, 153–160. [Google Scholar] [CrossRef]
- Bleve, G.; Tufariello, M.; Durante, M.; Perbellini, E.; Ramires, F.A.; Grieco, F. Physico-chemical and microbiological characterization of spontaneous fermentation of Cellina di Nardò and Leccino table olives. Front. Microbiol. 2014, 5, 570. [Google Scholar] [CrossRef] [PubMed]
- Laëtitia, G.; Pascal, D.; Yann, D. The citrate metabolism in homo and heterofermentative LAB: A selective means of becoming dominant over other microorganisms in complex ecosystems. Food Nutr. Sci. 2014, 5, 953–969. [Google Scholar] [CrossRef]
- Fleming, H.P.; Etchells, J.L.; Bell, T.A. Vapor analysis of fermented Spanish-type green olives by gas chromatography. J. Food Sci. 1969, 34, 419–422. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starter | Fermentation Time (Days) | Survival Rate (%) |
|---|---|---|
| C. boidinii Y27 | 0 | 100 |
| 75 | 45 | |
| 150 brine | 50 | |
| 150 olives | 45 | |
| C. boidinii Y28 | 0 | 100 |
| 75 | 10 | |
| 150 brine | 25 | |
| 150 olives | 0 | |
| C. boidinii Y30 | 0 | 85 |
| 75 | 30 | |
| 150 brine | 25 | |
| 150 olives | 10 | |
| C. boidinii Y31 | 0 | 90 |
| 75 | 15 | |
| 150 brine | 0 | |
| 150 olives | 10 | |
| S. cerevisiae Y34 | 0 | 100 |
| 75 | 5 | |
| 150 brine | 5 | |
| 150 olives | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonatsou, S.; Panagou, E.Z. Fermentation of cv. Kalamata Natural Black Olives with Potential Multifunctional Yeast Starters. Foods 2022, 11, 3106. https://doi.org/10.3390/foods11193106
Bonatsou S, Panagou EZ. Fermentation of cv. Kalamata Natural Black Olives with Potential Multifunctional Yeast Starters. Foods. 2022; 11(19):3106. https://doi.org/10.3390/foods11193106
Chicago/Turabian StyleBonatsou, Stamatoula, and Efstathios Z. Panagou. 2022. "Fermentation of cv. Kalamata Natural Black Olives with Potential Multifunctional Yeast Starters" Foods 11, no. 19: 3106. https://doi.org/10.3390/foods11193106
APA StyleBonatsou, S., & Panagou, E. Z. (2022). Fermentation of cv. Kalamata Natural Black Olives with Potential Multifunctional Yeast Starters. Foods, 11(19), 3106. https://doi.org/10.3390/foods11193106