



Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
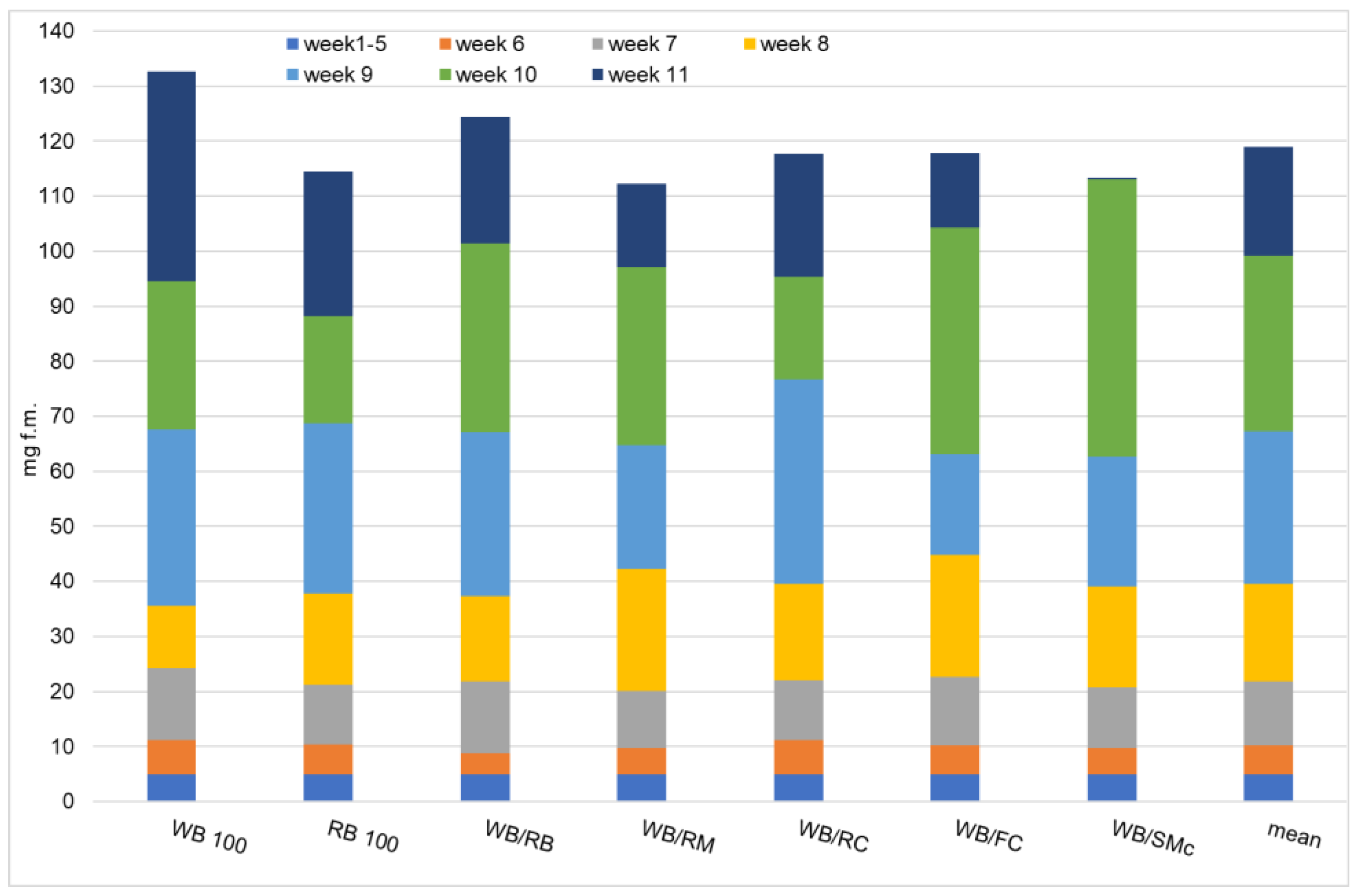
2.2. Diet Preparation, Larval Growth, and Measurements during the Experiment
2.3. Larvae Rearing Conditions
2.4. Feed Conversion Ratio
2.5. Nutritional Analysis
2.6. Statistical Analysis
3. Results and Discussion
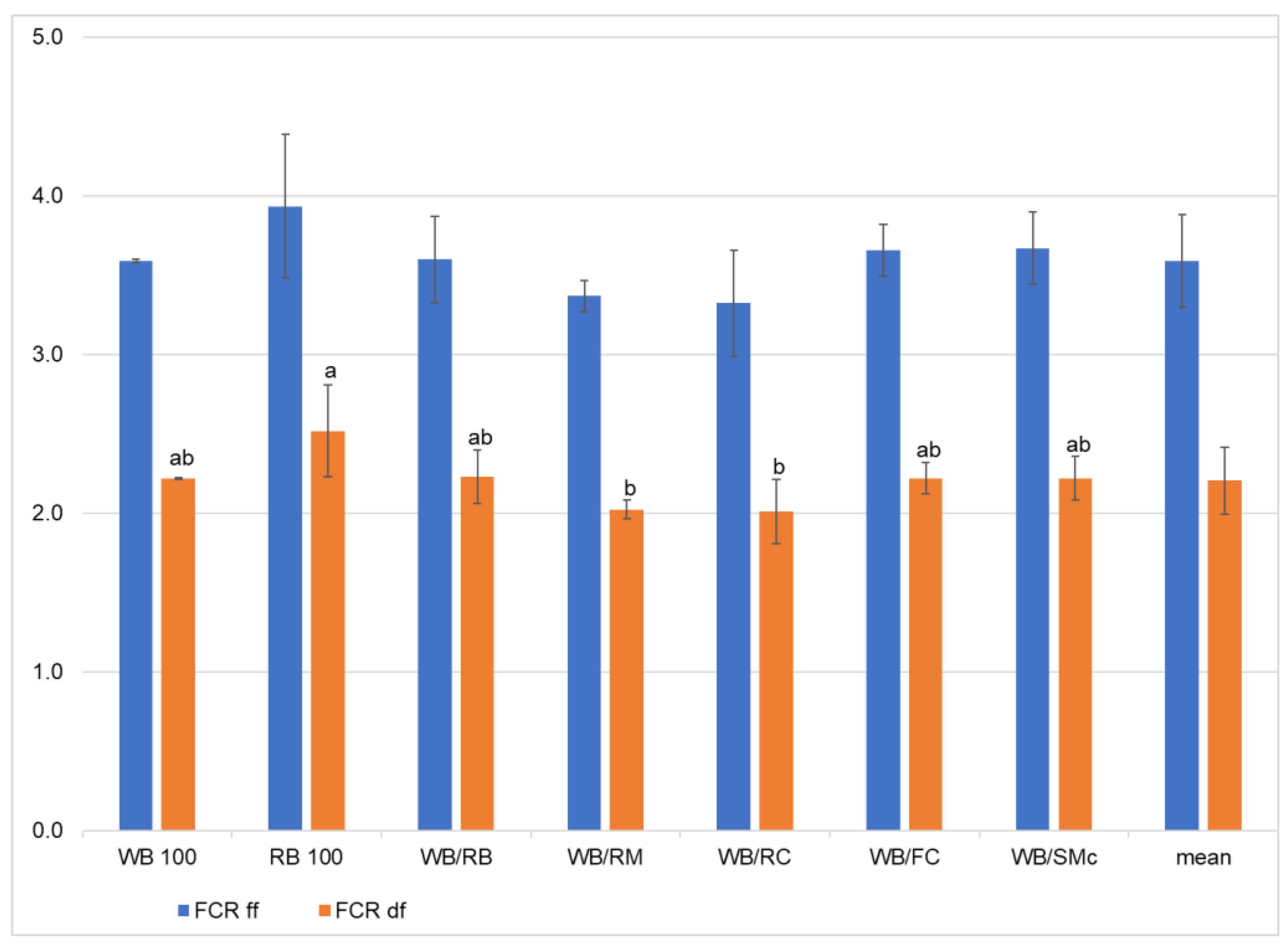
3.1. Growth Performance and Feed Conversion Ratio
3.2. Proximate Analysis
3.3. Fatty Acid Profile
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed consent statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; ESA Working Paper No. 12-03; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2012. [Google Scholar]
- FAO. Towards the Future We Want: End Hunger and Make the Transition to Sustainable Agricultural and Food Systems; FAO: Rome, Italy, 2012. [Google Scholar]
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- FAO. Global food losses and food waste—Extent, causes and prevention. In SAVE FOOD: An Initiative on Food Loss and Waste Reduction; FAO: Rome, Italy, 2011. [Google Scholar]
- Ben-Othman, S.; Jõudu, I.; Bhat, R. Bioactives from agri-food wastes: Present insights and future challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef]
- Ojha, S.; Bußler, S.; Schlüter, O.K. Food waste valorisation and circular economy concepts in insect production and processing. Waste Manag. 2020, 118, 600–609. [Google Scholar] [CrossRef]
- Murawska, D.; Daszkiewicz, T.; Sobotka, W.; Gesek, M.; Witkowska, D.; Matusevičius, P.; Bakuła, T. Partial and total replacement of soybean meal with full-fat black soldier fly (Hermetia illucens L.) larvae meal in broiler chicken diets: Impact on growth performance, carcass quality and meat quality. Animals 2021, 11, 2715. [Google Scholar] [CrossRef]
- Gałęcki, R.; Zielonka, Ł.; Zasępa, M.; Gołębiowska, J.; Bakuła, T. Potential utilization of edible insects as an alternative source of protein in animal diets in Poland. Front. Sustain. Food Syst. 2021, 5, 675796. [Google Scholar] [CrossRef]
- Chemello, G.; Renna, M.; Caimi, C.; Guerreiro, I.; Oliva-Teles, A.; Enes, P.; Biasato, I.; Schiavone, A.; Gai, F.; Gasco, L. Partially defatted Tenebrio molitor larva meal in diets for grow-out rainbow trout, Oncorhynchus mykiss (Walbaum): Effects on growth performance, diet digestibility and metabolic responses. Animals 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Volek, Z.; Adámková, A.; Zita, L.; Adámek, M.; Plachý, V.; Mlček, J.; Marounek, M. The effects of the dietary replacement of soybean meal with yellow mealworm larvae (Tenebrio molitor) on the growth, nutrient digestibility and nitrogen output of fattening rabbits. Anim. Feed Sci. Technol. 2021, 280, 115048. [Google Scholar] [CrossRef]
- Sedgh-Gooya, S.; Torki, M.; Darbemamieh, M.; Khamisabadi, H.; Abdolmohamadi, A. Growth performance and intestinal morphometric features of broiler chickens fed on dietary inclusion of yellow mealworm (Tenebrio molitor) larvae powder. Vet. Med. Sci. 2022; preprint. [Google Scholar] [CrossRef]
- Ritchie, H. How much of the world’s land would we need in order to feed the global population with the average diet of a given country? Our World in Data. October 2017, 3, 2017. Available online: https://ourworldindata.org/land-use (accessed on 5 January 2022).
- Jongema, Y. List of Edible Insects of the World, 2017. 2018. Available online: https://www.wur.nl/en/Research-Results/Chair-groups/Plant-Sciences/Laboratory-of-Entomology/Edible-insects/Worldwide-species-list.htm (accessed on 20 March 2022).
- EFSA Scientific Committee. Scientific Opinion on a risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef]
- European Commission. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 November and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, L.327, 1–22. [Google Scholar]
- European Commission. Regulation (EU) 2017/893 of 23 May 2017 amending Annexes I and IV on Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV, and XV to Commission Regulation (EU) No 142/2011 as regards the provisions on processed animal protein. Off. J. Eur. Union 2017, L. 138, 92–116. [Google Scholar]
- Federal Food Safety and Veterinary Office, Switzerland. Import of Foodstuffs and Utility Articles (Updated 2021). 2017. Available online: https://www.blv.admin.ch/blv/en/home/import-und-export/import/importe-aus-der-eu/lebensmittel-und-gebrauchsgegenstaende.html (accessed on 13 January 2022).
- EFSA Panel on Nutrition, Novel Foods and Food Allergens; Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A. Safety of frozen and dried formulations from whole yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06778. [Google Scholar]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will yellow mealworm become a source of safe proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- IPIFF (International Platform of Insects for Food and Feed). Guide on Good Hygiene Practices for European Union (EU) Producers of Insects as Food and Feed. 2019. Available online: https://ipiff.org/good-hygiene-practices/ (accessed on 5 March 2022).
- Ramos-Elorduy, J.; González, E.A.; Hernández, A.R.; Pino, J.M. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 2002, 95, 214–220. [Google Scholar] [CrossRef]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth potential of yellow mealworm reared on industrial residues. Agriculture 2020, 10, 599. [Google Scholar] [CrossRef]
- Van Broekhoven, S.; Oonincx, D.G.A.B.; Van Huis, A.; Van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P. The consumption and utilization of food by insects. Adv. Insect Physiol. 1968, 5, 229–288. [Google Scholar] [CrossRef]
- Miech, P.; Berggren, Å.; Lindberg, J.E.; Chhay, T.; Khieu, B.; Jansson, A. Growth and survival of reared Cambodian field crickets (Teleogryllus testaceus) fed weeds, agricultural and food industry by-products. J. Insects Food Feed 2016, 2, 285–292. [Google Scholar] [CrossRef]
- Boulos, S.; Tännler, A.; Nyström, L. Nitrogen-to-protein conversion factors for edible insects on the Swiss market: T. molitor, A. domesticus, and L. migratoria. Front. Nutr. 2020, 7, 89. [Google Scholar] [CrossRef]
- Zaderimowski, R.; Sosulski, F. Composition of total lipids in rapeseed. J. Am. Oil Chem. Soc. 1978, 55, 870–872. [Google Scholar] [CrossRef]
- Czaplicki, S.; Tańska, M.; Konopka, I. Sea-buckthorn oil in vegetable oils stabilisation. Ital. J. Food Sci. 2016, 28, 412–425. [Google Scholar] [CrossRef]
- Melis, R.; Braca, A.; Sanna, R.; Spada, S.; Mulas, G.; Fadda, M.L.; Sassu, M.M.; Serra, G.; Anedda, R. Metabolic response of yellow mealworm larvae to two alternative rearing substrates. Metabolomics 2019, 15, 113. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Chen, G.; Qiao, L.; Li, J.; Liu, B.; Liu, Z.; Li, M.; Liu, X. Growth performance and nutritional profile of mealworms reared on corn stover, soybean meal, and distillers’ grains. Eur. Food Res. Technol. 2019, 245, 2631–2640. [Google Scholar] [CrossRef]
- House, H.L. Effects of low levels of the nutrient content of a food and of nutrient imbalance on the feeding and the nutrition of a phytophagous larva, Celerio euphorbiae (Linnaeus) (Lepidoptera: Sphingidae). Can. Entomol. 1965, 97, 62–68. [Google Scholar] [CrossRef]
- Urrejola, S.; Nespolo, R.; Lardies, M.A. Diet-induced developmental plasticity in life histories and energy metabolism in a beetle. Rev. Chil. Hist. Nat. 2011, 84, 523–533. [Google Scholar] [CrossRef]
- Yu, X.; He, Q.; Wang, D. Dynamic analysis of major components in the different developmental stages of Tenebrio molitor. Front. Nutr. 2021, 8, 689746. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Laurent, S.; Veenenbos, M.E.; van Loon, J.J.A. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Sci. 2020, 27, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Song, Y.S.; Kim, M.W.; Moon, C.; Seo, D.J.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Park, Y.K.; Kim, S.A.; Kim, Y.W. Extraction of chitin and chitosan from larval exuvium and whole body of edible mealworm, Tenebrio molitor. Entomol. Res. 2018, 48, 227–233. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.F.; Benning, R.; Haase, H. Effect of different drying methods on nutrient quality of the yellow mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Krzyżaniak, M.; Aljewicz, M.; Bordiean, A.; Stolarski, M.J. Yellow mealworm composition after convective and freeze drying—Preliminary results. Agriculture 2022, 12, 149. [Google Scholar] [CrossRef]
- Harsányi, E.; Kovács, C.J.E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus larvae as alternative feed supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Van Peer, M.; Frooninckx, L.; Coudron, C.; Berrens, S.; Álvarez, C.; Deruytter, D.; Verheyen, G.; Van Miert, S. Valorisation potential of using organic side streams as feed for Tenebrio molitor, Acheta domesticus and Locusta migratoria. Insects 2021, 12, 796. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary fatty acids influence the growth and fatty acid composition of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef]
- Lease, H.M.; Wolf, B.O. Lipid content of terrestrial arthropods in relation to body size, phylogeny, ontogeny and sex. Physiol. Entomol. 2011, 36, 29–38. [Google Scholar] [CrossRef]
- Oonincx, D.; Dierenfeld, E.S. An investigation into the chemical composition of alternative invertebrate prey. Zoo Biol. 2012, 31, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Francardi, V.; Cito, A.; Fusi, S.; Botta, M.; Dreassi, E. Linseed to increase N-3 fatty acids in Tenebrio molitor (Coleoptera Tenebrionidae). Redia 2017, 100, 73–76. [Google Scholar] [CrossRef]
- Russo, G.L. Dietary n − 6 and n − 3 polyunsaturated fatty acids: From biochemistry to clinical implications in cardiovascular prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Sanmartin, C.; Taglieri, I.; Venturi, F.; Macaluso, M.; Zinnai, A.; Tavarini, S.; Botto, A.; Serra, A.; Conte, G.; Flamini, G. Flaxseed cake as a tool for the improvement of nutraceutical and sensorial features of sourdough bread. Foods 2020, 9, 204. [Google Scholar] [CrossRef]
- Marszałkiewicz, S.; Siger, A.; Gawrysiak-Witulska, M.; Kmiecik, D.; Rudzińska, M. The effect of drying temperature of milk thistle seeds on quality and bioactive compounds in the lipid fraction. J. Food Sci. Technol. 2020, 57, 4003–4013. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No. | Substrate | Moisture (%) | Ash (% d.m.) | Crude Protein (% d.m.) | Fiber Content (% d.m.) | Crude Fat (% d.m.) | NFE (% d.m.) |
|---|---|---|---|---|---|---|---|
| 1. | Wheat bran (WB) | 12.46 | 5.63 | 17.34 | 8.29 | 4.12 | 64.62 |
| 2. | Rye bran (RB) | 11.41 | 4.16 | 15.04 | 3.79 | 2.76 | 74.25 |
| 3. | Rapeseed meal (RM) | 11.88 | 7.33 | 35.41 | 10.57 | 1.88 | 44.81 |
| 4. | Rapeseed cake (RC) | 9.88 | 5.83 | 27.68 | 16.74 | 16.58 | 33.17 |
| 5. | Flax cake (FC) | 8.73 | 5.47 | 27.01 | 5.47 | 13.82 | 48.23 |
| 6. | Milk thistle cake (SMc) | 9.61 | 5.41 | 17.98 | 26.23 | 3.62 | 47.77 |
| No. | Diets | Moisture (%) | Ash (% d.m.) | Protein (% d.m.) | Fiber (% d.m.) | Crude Fat (% d.m.) | NFE (% d.m.) |
|---|---|---|---|---|---|---|---|
| 1 | WB 100 (control) | 12.46 | 5.63 | 17.34 | 8.29 | 4.12 | 64.62 |
| 2 | RB 100 | 11.41 | 4.16 | 15.04 | 3.79 | 2.76 | 74.25 |
| 3 | WB/RB | 12.1 | 5.2 | 16.6 | 6.9 | 3.71 | 67.51 |
| 4 | WB/RM | 12.3 | 6.1 | 22.8 | 9.0 | 3.45 | 58.68 |
| 5 | WB/RC | 11.7 | 5.7 | 20.4 | 10.8 | 7.86 | 55.18 |
| 6 | WB/FC | 11.3 | 5.6 | 20.2 | 7.4 | 7.03 | 59.70 |
| 7 | WB/SMc | 11.6 | 5.6 | 17.5 | 13.7 | 3.97 | 59.27 |
| Source of Variation | FCRdf | FCRff | ECI | Final Weight | Dry Matter | Ash | Crude Fiber | Crude Protein | Crude Fat |
|---|---|---|---|---|---|---|---|---|---|
| df | 6 | ||||||||
| F | 3.22 | 1.82 | 3.18 | 2.20 | 20.1 | 2.87 | 0.658 | 2463 | 130 |
| p | 0.03 | 0.17 | 0.035 | 0.10 | <0.001 | 0.05 | 0.68 | <0.001 | <0.001 |
| Feed | Dry Matter (%) | Ash (%) | Fiber Content (%) | Crude Protein (%) | Crude Fat (%) |
|---|---|---|---|---|---|
| WB 100 | 31.7 ± 0.31 bc | 3.80 ± 0.08 | 6.85 ± 0.32 | 47.9 ± 0.10 f | 26.1 ± 0.82 c |
| RB 100 | 33.3 ± 0.15 a | 3.64 ± 0.05 | 7.12 ± 0.63 | 43.6 ± 0.02 g | 30.0 ± 0.47 a |
| WB/RB | 31.6 ± 0.15 bc | 4.00 ± 0.09 | 7.12 ± 0.42 | 49.3 ± 0.02 e | 27.5 ± 0.69 bc |
| WB/RM | 32.1 ± 0.25 b | 3.85 ± 0.06 | 7.06 ± 0.34 | 52.5 ± 0.06 b | 22.3 ± 0.09 e |
| WB/RC | 32.3 ± 0.77 ab | 3.81 ± 0.07 | 6.79 ± 0.59 | 50.9 ± 0.19 d | 28.6 ± 0.52 b |
| WB/FC | 30.6 ± 0.30 c | 5.19 ± 1.18 | 7.31 ± 0.31 | 53.4 ± 0.11 a | 24.4 ± 0.40 d |
| WB/SMc | 30.3 ± 0.46 d | 4.83 ± 1.11 | 7.29 ± 0.25 | 52.0 ± 0.19 c | 24.5 ± 0.23 d |
| Common Name | WB 100 * | RB 100 * | WB/RB | WB/RM * | WB/RC * | WB/ FC * | WB/SMc | |
|---|---|---|---|---|---|---|---|---|
| Lauric acid | C12:0 | 0.22 | 0.26 | 0.23 | 0.19 | 0.14 | 0.14 | 0.18 |
| Tridecanoic acid | C13:0 | 0.05 | 0.04 | 0.05 | 0.05 | 0.04 | 0.04 | 0.05 |
| Myristic acid | C14:0 | 2.55 | 3.29 | 2.67 | 2.19 | 1.71 | 1.91 | 2.27 |
| Myristoleic acid | C14:1 | 0.12 | 0.15 | 0.13 | 0.12 | 0.13 | 0.07 | 0.10 |
| C14:2 | 0.12 | 0.13 | 0.13 | 0.14 | 0.10 | 0.08 | 0.11 | |
| Pentadecanoic acid | C15:0 | 0.15 | 0.11 | 0.14 | 0.21 | 0.13 | 0.13 | 0.16 |
| Pentadecanoic acid | C15:1 | 0.07 | 0.06 | 0.06 | 0.14 | 0.08 | 0.08 | 0.08 |
| Palmitic acid | C16:0 | 17.33 | 17.81 | 17.25 | 16.26 | 12.80 | 14.25 | 15.44 |
| Palmitoleic acid | C16:1 n (7 + 9) | 2.12 | 1.91 | 2.42 | 2.27 | 2.08 | 1.30 | 1.33 |
| Hexadecadienoic acid | C16:2 n-6,9 | 0.48 | 0.27 | 0.40 | 0.55 | 0.50 | 0.39 | 0.51 |
| Margaroleic acid | C17:1 | 0.14 | 0.13 | 0.13 | 0.74 | n.d. | 0.52 | 0.63 |
| Stearic acid | C18:0 | 2.43 | 2.33 | 2.32 | 2.50 | 2.21 | 0.24 | 3.17 |
| Oleic | C18:1 n-9 | 36.53 | 46.74 | 39.58 | 39.32 | 46.34 | 32.97 | 33.46 |
| Vaccenic | C18:1 | 0.23 | 0.38 | 0.29 | 0.34 | 0.31 | 0.54 | 0.26 |
| Linoleic acid | C18:2 n-6 | 35.55 | 22.79 | 31.66 | 31.73 | 29.75 | 29.12 | 38.98 |
| α-linolenic acid | C18:3 | 1.73 | 1.11 | 2.33 | 1.86 | 2.60 | 17.00 | 1.50 |
| Arachidic acid | C20:0 | 0.08 | 0.14 | 0.11 | 0.07 | 0.08 | n.d. | 0.16 |
| Gondoic acid | C20:1 n-9 | 0.09 | n.d. | 0.10 | 0.11 | 0.16 | 0.09 | 0.12 |
| Eicosadienoic acid | C20:2 n-6 | 0.06 | n.d. | n.d. | n.d. | n.d. | 0.04 | 0.05 |
| SFA (% TFA) | 22.79 | 23.96 | 22.77 | 21.45 | 17.11 | 16.71 | 21.42 | |
| MUFA (% TFA) | 39.29 | 49.35 | 42.71 | 43.03 | 49.08 | 35.56 | 35.97 | |
| PUFA (% TFA) | 37.93 | 24.30 | 34.52 | 34.27 | 32.95 | 46.63 | 41.15 | |
| UFA (% TFA) | 77.21 | 73.65 | 77.23 | 77.30 | 82.03 | 82.19 | 77.12 | |
| SFA/UFA | 0.30 | 0.33 | 0.29 | 0.28 | 0.21 | 0.20 | 0.28 | |
| PUFA/SFA | 1.66 | 1.01 | 1.52 | 1.60 | 1.93 | 2.79 | 1.92 | |
| ω6/ω3 (n6/n3) | 20.55 | 20.62 | 13.56 | 17.11 | 11.44 | 1.71 | 25.99 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bordiean, A.; Krzyżaniak, M.; Aljewicz, M.; Stolarski, M.J. Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods 2022, 11, 3075. https://doi.org/10.3390/foods11193075
Bordiean A, Krzyżaniak M, Aljewicz M, Stolarski MJ. Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods. 2022; 11(19):3075. https://doi.org/10.3390/foods11193075
Chicago/Turabian StyleBordiean, Anna, Michał Krzyżaniak, Marek Aljewicz, and Mariusz Jerzy Stolarski. 2022. "Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm" Foods 11, no. 19: 3075. https://doi.org/10.3390/foods11193075
APA StyleBordiean, A., Krzyżaniak, M., Aljewicz, M., & Stolarski, M. J. (2022). Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods, 11(19), 3075. https://doi.org/10.3390/foods11193075