
Novel Candidate Microorganisms for Fermentation Technology: From Potential Benefits to Safety Issues

 ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Certain Novel Microorganisms for Fermentation
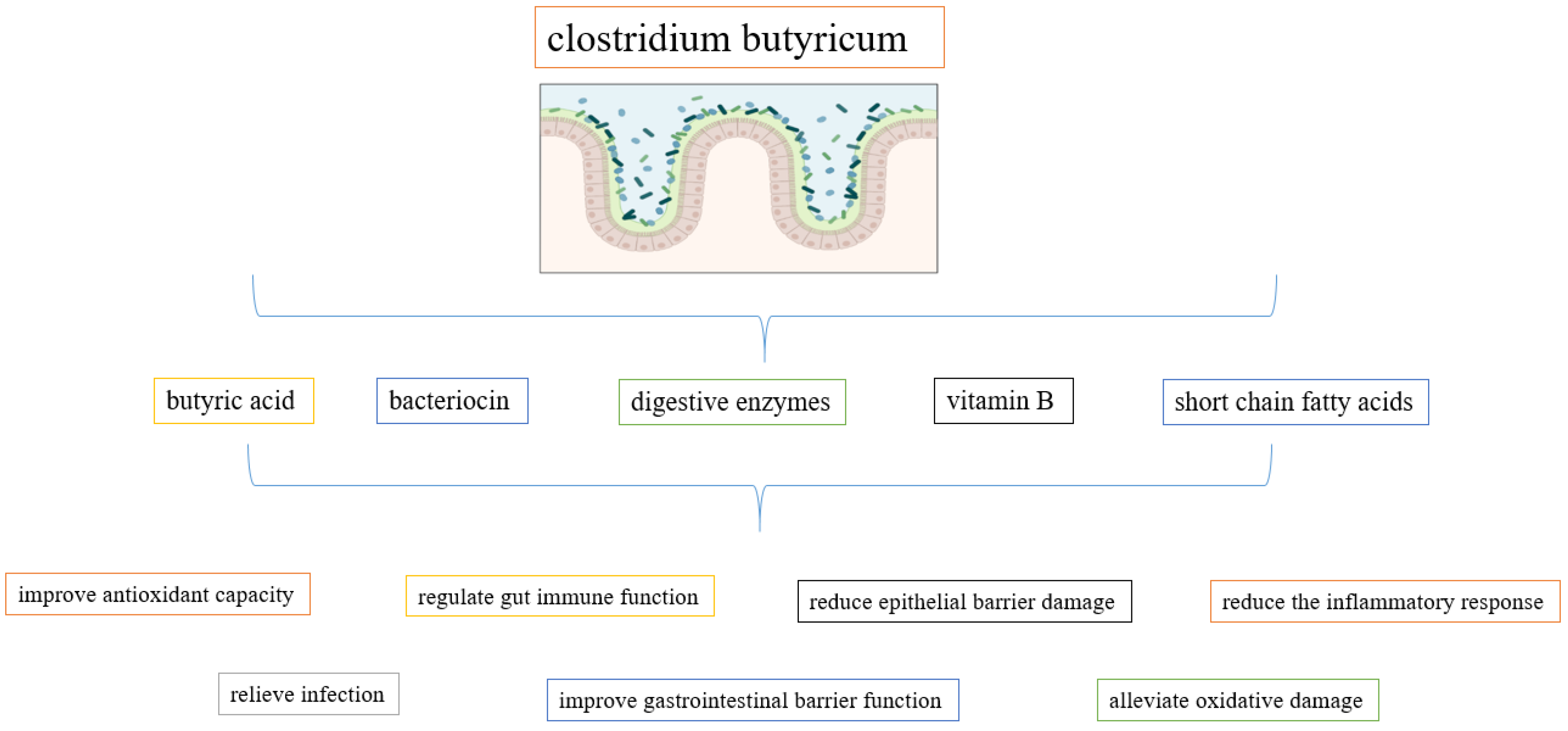
2.1. Clostridium butyricum
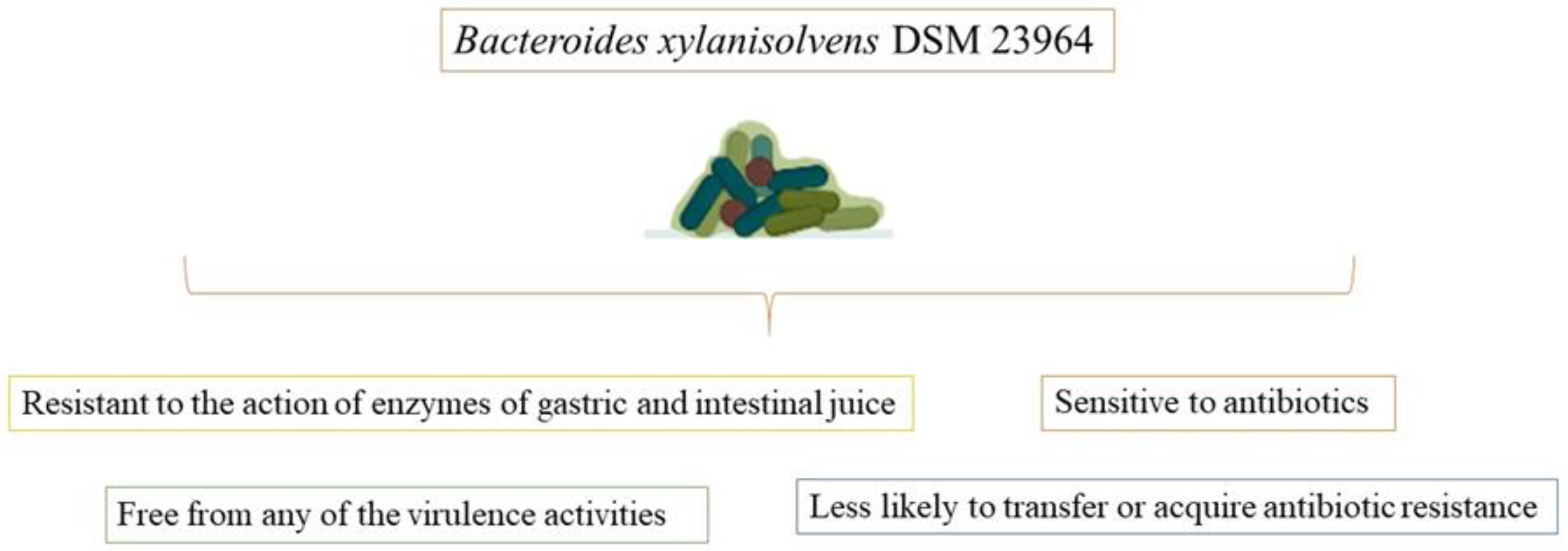
2.2. Bacteroides xylanisolvens
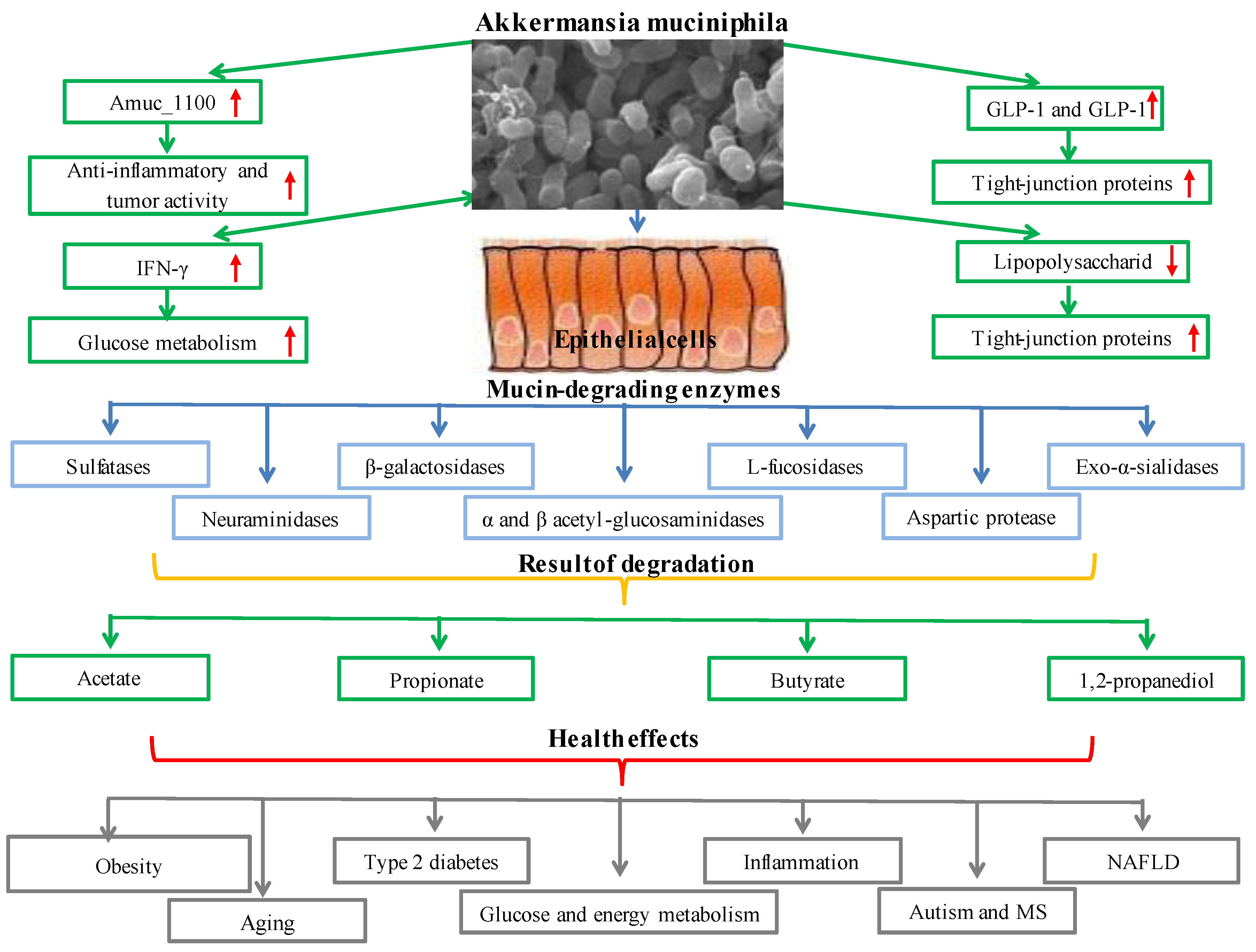
2.3. Akkermansia municiphila
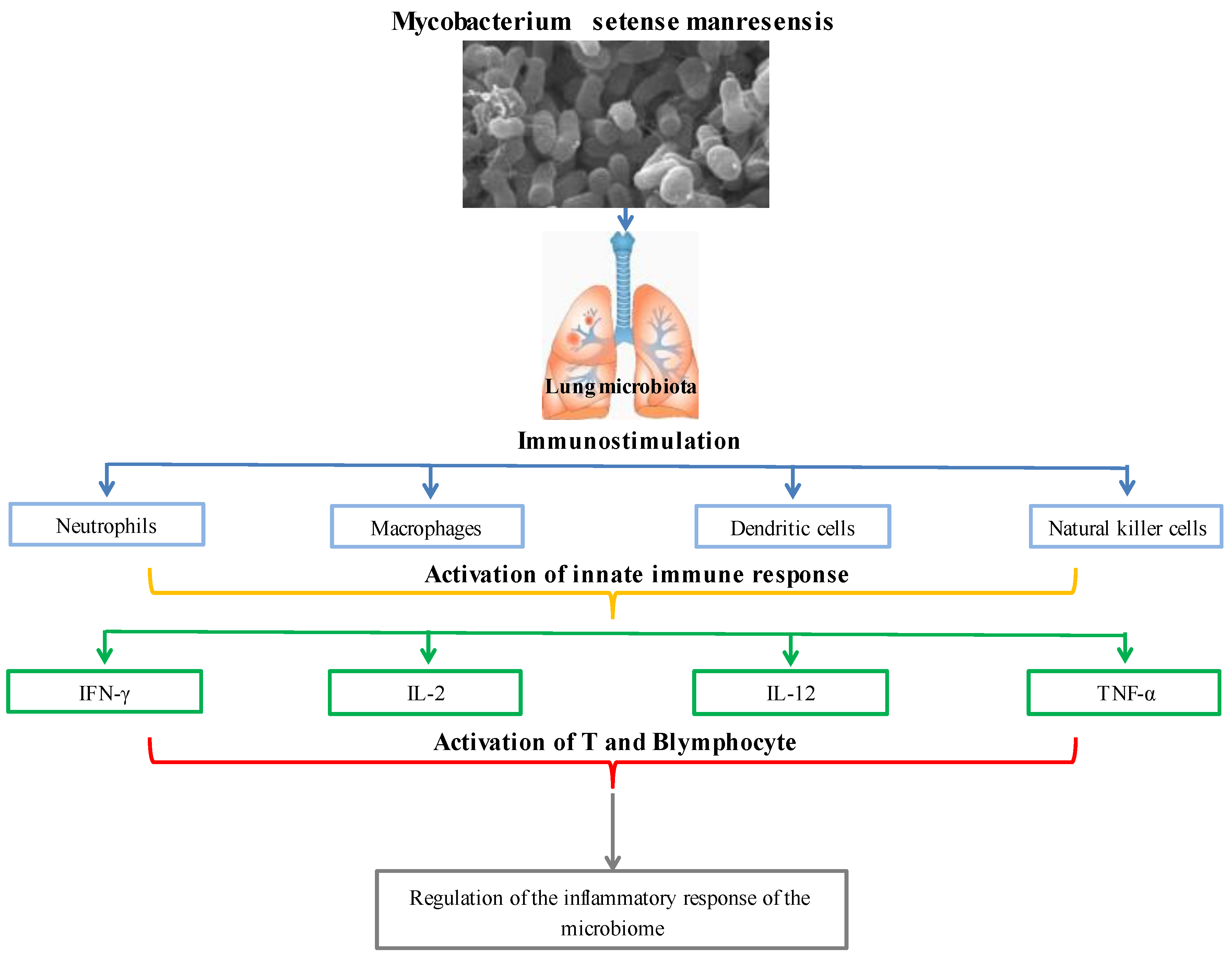
2.4. Mycobacterium setense manresensis
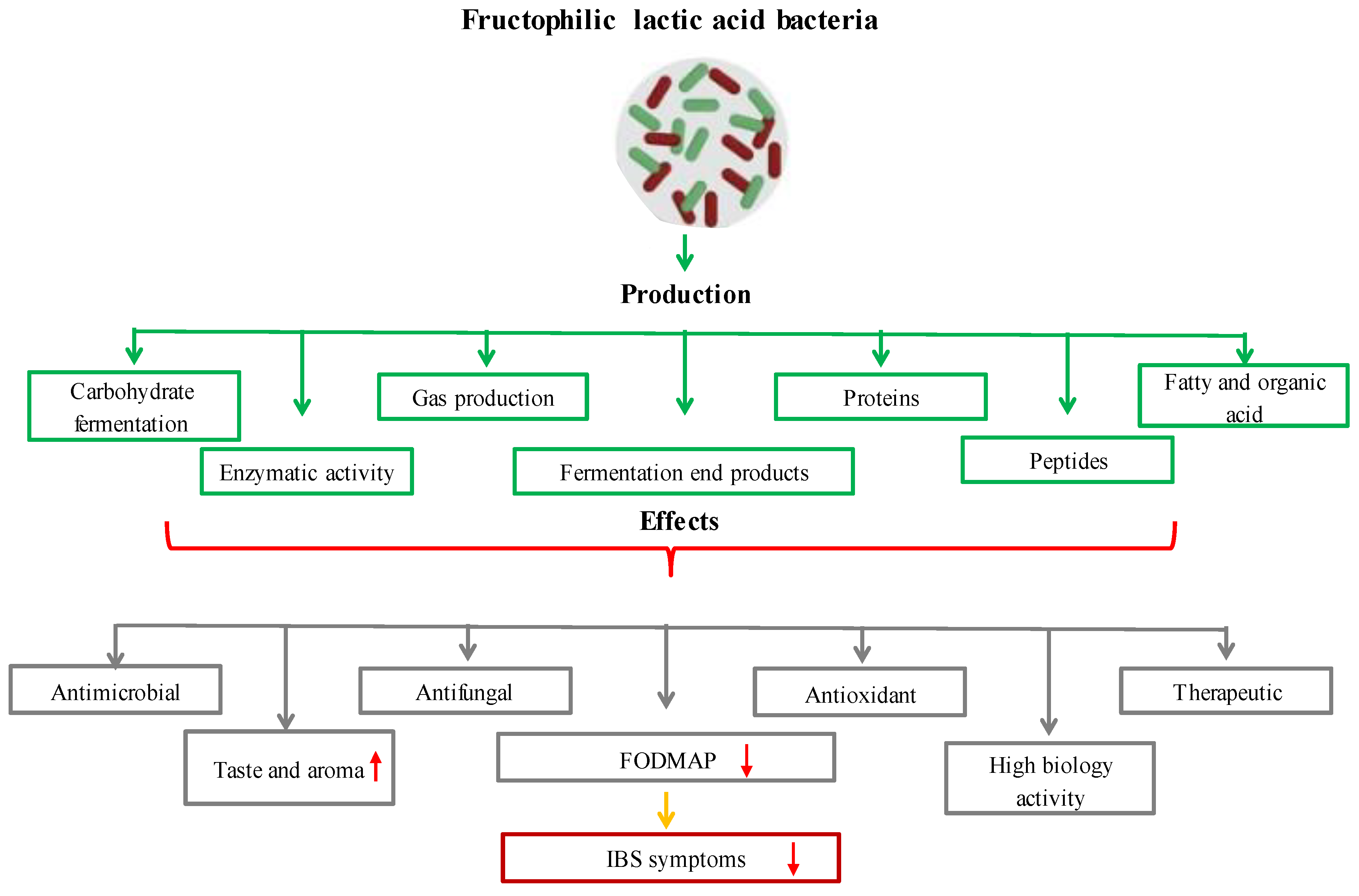
2.5. Novel Lactic Acid Bacteria (Fructophilic Lactic Acid Bacteria (FLAB))
2.6. Some Other Novel Microorganisms
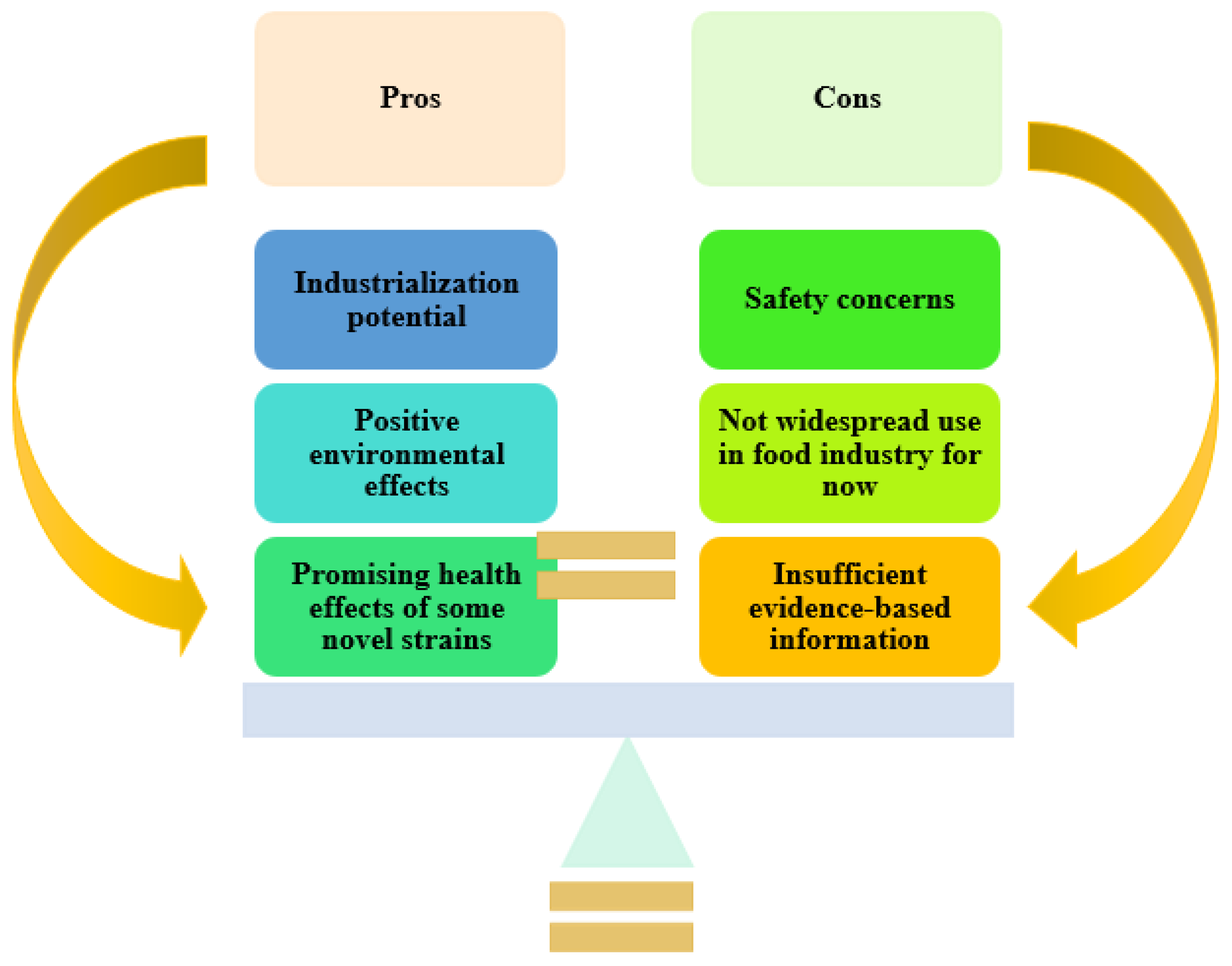
2.7. The Safety Issues of Novel Microorganisms
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gänzle, M. The periodic table of fermented foods: Limitations and opportunities. Appl. Microbiol. Biotechnol. 2022, 106, 2815–2826. [Google Scholar] [CrossRef] [PubMed]
- Hayden, B.; Canuel, N.; Shanse, J. What Was Brewing in the Natufian? An Archaeological Assessment of Brewing Technology in the Epipaleolithic. J. Archaeol. Method Theory 2013, 20, 102–150. [Google Scholar] [CrossRef]
- Arranz-Otaegui, A.; Gonzalez Carretero, L.; Ramsey, M.N.; Fuller, D.Q.; Richter, T. Archaeobotanical evidence reveals the origins of bread 14,400 years ago in northeastern Jordan. Proc. Natl. Acad. Sci. USA 2018, 115, 7925–7930. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.T.; Lu, P.; Parrella, J.A.; Leggette, H.R. Consumer Acceptance toward Functional Foods: A Scoping Review. Int. J. Environ. Res. Public Health 2022, 19, 1217. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- Vilela, A.; Bacelar, E.; Pinto, T.; Anjos, R.; Correia, E.; Gonçalves, B.; Cosme, F. Beverage and Food Fragrance Biotechnology, Novel Applications, Sensory and Sensor Techniques: An Overview. Foods 2020, 8, 643. [Google Scholar] [CrossRef]
- The International Scientific Association for Probiotics and Prebiotics (ISAPP). Fermented Foods. Available online: https://isappscience.org/for-scientists/resources/fermented-foods/#:~:text=Photo%20by%20https%3A%2F%2Fdeankalyan,enzymatic%20conversions%20of%20food%20components%E2%80%9D (accessed on 28 July 2022).
- Harnentis, H.; Marlida, Y.; Nur, Y.S.; Wizna, W.; Santi, M.A.; Septiani, N.; Adzitey, F.; Huda, N. Novel probiotic lactic acid bacteria isolated from indigenous fermented foods from West Sumatera, Indonesia. Vet. World 2020, 13, 1922–1927. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.-H.; Jung, S.-J.; Chae, S.-W. Functional Properties of Microorganisms in Fermented Foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health Benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. BioMed Res. Int. 2018, 2018, 9361614. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Gómez-Sala, B.; O’Connor, E.M.; Kenny, J.G.; Cotter, P.D. Global Regulatory Frameworks for Fermented Foods: A Review. Front. Nutr. 2022, 9, 902642. [Google Scholar] [CrossRef]
- EUR-Lex. European Parliament and Council Regulation (EC)N178/2002 of 28 January 2002 laying down the general principles and requirements of food law, establishing the European Food Safety Authority and laying down procedures in matters of food safety. Off. J. Eur. Commun. L 2015, 31, 1–24. [Google Scholar]
- Lavefve, L.; Marasini, D.; Carbonero, F. Microbial Ecology of Fermented Vegetables and Non-Alcoholic Drinks and Current Knowledge on Their Impact on Human Health. Adv. Food Nutr. Res. 2019, 87, 147–185. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, K. Health benefits of fermented foods. Nat. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Castellone, V.; Bancalari, E.; Rubert, J.; Gatti, M.; Neviani, E.; Bottari, B. Eating Fermented: Health Benefits of LAB-Fermented Foods. Foods 2021, 10, 2639. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Ray, R.; Joshi, V. Fermented foods: Past, present and future. In Microorganisms and Fermentation of Traditional Foods; Ray, R.C., Montet, D., Eds.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Khorshidian, N.; Yousefi, M.; Mortazavian, A.M. Chapter Three—Fermented milk: The most popular probiotic food carrier. In Advances in Food and Nutrition Research; da Cruz, A.G., Prudencio, E.S., Esmerino, E.A., da Silva, M.C., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 94, pp. 91–114. [Google Scholar]
- Kabak, B.; Dobson, A.D. An introduction to the traditional fermented foods and beverages of Turkey. Crit. Rev. Food Sci. Nutr. 2011, 51, 248–260. [Google Scholar] [CrossRef]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented foods, the gut and mental health: A mechanistic overview with implications for depression and anxiety. Nutr. Neurosci. 2020, 23, 659–671. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V.; Luziatelli, F.; Ficca, A.G.; Ruzzi, M. Health-Promoting Components in Fermented Foods: An Up-to-Date Systematic Review. Nutrients 2019, 11, 1189. [Google Scholar] [CrossRef]
- Grujović, M.Ž.; Mladenović, K.G.; Semedo-Lemsaddek, T.; Laranjo, M.; Stefanović, O.D.; Kocić-Tanackov, S.D. Advantages and disadvantages of non-starter lactic acid bacteria from traditional fermented foods: Potential use as starters or probiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1537–1567. [Google Scholar] [CrossRef] [PubMed]
- Faintuch, J.; Faintuch, J.J. Chapter 46—Precision medicine: The microbiome and metabolome. In Microbiome and Metabolome in Diagnosis, Therapy, and other Strategic Applications; Faintuch, J., Faintuch, S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 435–449. [Google Scholar]
- Cai, G.; Jin, B.; Monis, P.; Saint, C. A genetic and metabolic approach to redirection of biochemical pathways of Clostridium butyricum for enhancing hydrogen production. Biotechnol. Bioeng. 2013, 110, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Jin, B.; Saint, C.; Monis, P. Genetic manipulation of butyrate formation pathways in Clostridium butyricum. J. Biotechnol. 2011, 155, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Chen, T. Engineered Akkermansia muciniphila: A promising agent against diseases (Review). Exp. Ther. Med. 2020, 20, 285. [Google Scholar] [CrossRef] [PubMed]
- Brodmann, T.; Endo, A.; Gueimonde, M.; Vinderola, G.; Kneifel, W.; de Vos, W.M.; Salminen, S.; Gómez-Gallego, C. Safety of Novel Microbes for Human Consumption: Practical Examples of Assessment in the European Union. Front. Microbiol. 2017, 8, 1725. [Google Scholar] [CrossRef]
- Ariyoshi, T.; Hagihara, M.; Takahashi, M.; Mikamo, H. Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines 2022, 10, 483. [Google Scholar] [CrossRef]
- Ariyoshi, T.; Hagihara, M.; Tomono, S.; Eguchi, S.; Minemura, A.; Miura, D.; Oka, K.; Takahashi, M.; Yamagishi, Y.; Mikamo, H. Clostridium butyricum MIYAIRI 588 Modifies Bacterial Composition under Antibiotic-Induced Dysbiosis for the Activation of Interactions via Lipid Metabolism between the Gut Microbiome and the Host. Biomedicines 2021, 9, 1065. [Google Scholar] [CrossRef]
- Kim, D.-H.; Yoon, J.-J.; Kim, S.-H.; Park, J.-H. Acceleration of lactate-utilizing pathway for enhancing biohydrogen production by magnetite supplementation in Clostridium butyricum. Bioresour. Technol. 2022, 359, 127448. [Google Scholar] [CrossRef]
- Martins, F.F.; Liberato, V.d.S.; Ribeiro, C.M.S.; Coelho, M.A.Z.; Ferreira, T.F. Low-cost medium for 1, 3-propanediol production from crude glycerol by Clostridium butyricum. Biofuels Bioprod. Biorefining 2020, 14, 1125–1134. [Google Scholar] [CrossRef]
- Szymanowska-Powałowska, D.; Orczyk, D.; Leja, K. Biotechnological potential of Clostridium butyricum bacteria. Braz. J. Microbiol. 2014, 45, 892–901. [Google Scholar] [CrossRef]
- Ortigueira, J.; Martins, L.; Pacheco, M.; Silva, C.; Moura, P. Improving the non-sterile food waste bioconversion to hydrogen by microwave pretreatment and bioaugmentation with Clostridium butyricum. Waste Manag. 2019, 88, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Fang, B.; Wang, S. Effects of culture conditions on the kinetic behavior of 1, 3-propanediol fermentation by Clostridium butyricum with a kinetic model. Bioresour. Technol. 2016, 212, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Liberato, V.S.; Martins, F.F.; Ribeiro, C.M.S.; Coelho, M.A.Z.; Ferreira, T.F. Two-waste culture medium to produce 1, 3-propanediol through a wild Clostridium butyricum strain. Fuel 2022, 322, 124202. [Google Scholar] [CrossRef]
- Arisht, S.N.; Roslan, R.; Gie, G.A.; Mahmod, S.S.; Sajab, M.S.; Lay, C.-H.; Wu, S.-Y.; Ding, G.-T.; Jamali, N.S.; Jahim, J.M. Effect of nano zero-valent iron (nZVI) on biohydrogen production in anaerobic fermentation of oil palm frond juice using Clostridium butyricum JKT37. Biomass Bioenergy 2021, 154, 106270. [Google Scholar] [CrossRef]
- Masset, J.; Hiligsmann, S.; Hamilton, C.; Beckers, L.; Franck, F.; Thonart, P. Effect of pH on glucose and starch fermentation in batch and sequenced-batch mode with a recently isolated strain of hydrogen-producing Clostridium butyricum CWBI1009. Int. J. Hydrog. Energy 2010, 35, 3371–3378. [Google Scholar] [CrossRef]
- Gupta, P.; Kumar, M.; Gupta, R.P.; Puri, S.K.; Ramakumar, S. Fermentative reforming of crude glycerol to 1, 3-propanediol using Clostridium butyricum strain L4. Chemosphere 2022, 292, 133426. [Google Scholar] [CrossRef]
- Sun, Y.-Q.; Shen, J.-T.; Yan, L.; Zhou, J.-J.; Jiang, L.-L.; Chen, Y.; Yuan, J.-L.; Feng, E.; Xiu, Z.-L. Advances in bioconversion of glycerol to 1, 3-propanediol: Prospects and challenges. Process Biochem. 2018, 71, 134–146. [Google Scholar] [CrossRef]
- Kumar, V.; Park, S. Potential and limitations of Klebsiella pneumoniae as a microbial cell factory utilizing glycerol as the carbon source. Biotechnol. Adv. 2018, 36, 150–167. [Google Scholar] [CrossRef]
- Zhou, S.; Lama, S.; Sankaranarayanan, M.; Park, S. Metabolic engineering of Pseudomonas denitrificans for the 1, 3-propanediol production from glycerol. Bioresour. Technol. 2019, 292, 121933. [Google Scholar] [CrossRef]
- Liberato, V.; Benevenuti, C.; Coelho, F.; Botelho, A.; Amaral, P.; Pereira Jr, N.; Ferreira, T. Clostridium sp. as bio-catalyst for fuels and chemicals production in a biorefinery context. Catalysts 2019, 9, 962. [Google Scholar] [CrossRef]
- Dolejš, I.; Líšková, M.; Krasňan, V.; Markošová, K.; Rosenberg, M.; Lorenzini, F.; Marr, A.C.; Rebroš, M. Production of 1, 3-propanediol from pure and crude glycerol using immobilized Clostridium butyricum. Catalysts 2019, 9, 317. [Google Scholar] [CrossRef]
- Tiang, M.F.; Hanipa, M.A.F.; Abdul, P.M.; Jahim, J.M.; Mahmod, S.S.; Takriff, M.S.; Lay, C.-H.; Reungsang, A.; Wu, S.-Y. Recent advanced biotechnological strategies to enhance photo-fermentative biohydrogen production by purple non-sulphur bacteria: An overview. Int. J. Hydrog. Energy 2020, 45, 13211–13230. [Google Scholar] [CrossRef]
- Hiligsmann, S.; Masset, J.; Hamilton, C.; Beckers, L.; Thonart, P. Comparative study of biological hydrogen production by pure strains and consortia of facultative and strict anaerobic bacteria. Bioresour. Technol. 2011, 102, 3810–3818. [Google Scholar] [CrossRef] [PubMed]
- Hiligsmann, S.; Beckers, L.; Masset, J.; Hamilton, C.; Thonart, P. Improvement of fermentative biohydrogen production by Clostridium butyricum CWBI1009 in sequenced-batch, horizontal fixed bed and biodisc-like anaerobic reactors with biomass retention. Int. J. Hydrog. Energy 2014, 39, 6899–6911. [Google Scholar] [CrossRef]
- Hamilton, C.; Calusinska, M.; Baptiste, S.; Masset, J.; Beckers, L.; Thonart, P.; Hiligsmann, S. Effect of the nitrogen source on the hydrogen production metabolism and hydrogenases of Clostridium butyricum CWBI1009. Int. J. Hydrog. Energy 2018, 43, 5451–5462. [Google Scholar] [CrossRef]
- Hagihara, M.; Kuroki, Y.; Ariyoshi, T.; Higashi, S.; Fukuda, K.; Yamashita, R.; Matsumoto, A.; Mori, T.; Mimura, K.; Yamaguchi, N. Clostridium butyricum modulates the microbiome to protect intestinal barrier function in mice with antibiotic-induced dysbiosis. iScience 2020, 23, 100772. [Google Scholar] [CrossRef]
- Yan, H.; Ajuwon, K.M. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PLoS ONE 2017, 12, e0179586. [Google Scholar] [CrossRef]
- Gao, Q.; Wu, T.; Wang, J.; Zhuang, Q. Inhibition of bacterial adhesion to HT-29 cells by lipoteichoic acid extracted from Clostridium butyricum. Afr. J. Biotechnol. 2011, 10, 7633–7639. [Google Scholar]
- Kong, Q.; He, G.-Q.; Jia, J.-L.; Zhu, Q.-L.; Ruan, H. Oral administration of Clostridium butyricum for modulating gastrointestinal microflora in mice. Curr. Microbiol. 2011, 62, 512–517. [Google Scholar] [CrossRef]
- Liu, J.; Fu, Y.; Zhang, H.; Wang, J.; Zhu, J.; Wang, Y.; Guo, Y.; Wang, G.; Xu, T.; Chu, M. The hepatoprotective effect of the probiotic Clostridium butyricum against carbon tetrachloride-induced acute liver damage in mice. Food Funct. 2017, 8, 4042–4052. [Google Scholar] [CrossRef]
- Chassard, C.; Delmas, E.; Lawson, P.A.; Bernalier-Donadille, A. Bacteroides xylanisolvens sp. nov., a xylan-degrading bacterium isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Narushima, S.; Itoh, K.; Takamine, F.; Uchida, K. Absence of cecal secondary bile acids in gnotobiotic mice associated with two human intestinal bacteria with the ability to dehydroxylate bile acids in vitro. Microbiol. Immunol. 1999, 43, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van de Wiele, T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Kasper, D.L. Novel Tools for Modulating Immune Responses in the Host—Polysaccharides from the Capsule of Commensal Bacteria. Adv. Immunol. 2010, 106, 61–91. [Google Scholar] [PubMed]
- Ulsemer, P.; Toutounian, K.; Schmidt, J.; Karsten, U.; Goletz, S. Preliminary safety evaluation of a new Bacteroides xylanisolvens isolate. Appl. Environ. Microbiol. 2012, 78, 528–535. [Google Scholar] [CrossRef]
- Ulsemer, P.; Toutounian, K.; Schmidt, J.; Leuschner, J.; Karsten, U.; Goletz, S. Safety assessment of the commensal strain Bacteroides xylanisolvens DSM 23964. Regul. Toxicol. Pharmacol. 2012, 62, 336–346. [Google Scholar] [CrossRef]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies. Scientific Opinion on the safety of ‘heat-treated milk products fermented with Bacteroides xylanisolvens DSM 23964′as a novel food. EFSA J. 2015, 13, 3956. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Xing, J.; Li, X.; Sun, Y.; Zhao, J.; Miao, S.; Xiong, Q.; Zhang, Y.; Zhang, G. Comparative genomic and functional analysis of Akkermansia muciniphila and closely related species. Genes Genom. 2019, 41, 1253–1264. [Google Scholar] [CrossRef]
- Derrien, M.; Collado, M.C.; Ben-Amor, K.; Salminen, S.; de Vos, W.M. The Mucin degrader Akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl. Environ. Microbiol. 2008, 74, 1646–1648. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Sunder, S.; Verma, S.R. Disease-associated dysbiosis and potential therapeutic role of Akkermansia muciniphila, a mucus degrading bacteria of gut microbiome. Folia Microbiol. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kosciow, K.; Deppenmeier, U. Characterization of three novel β-galactosidases from Akkermansia muciniphila involved in mucin degradation. Int. J. Biol. Macromol. 2020, 149, 331–340. [Google Scholar] [CrossRef]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H. Mucolytic bacteria with increased prevalence in IBD mucosa augmentin vitroutilization of mucin by other bacteria. Off. J. Am. Coll. Gastroenterol. ACG 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- Kostopoulos, I.; Elzinga, J.; Ottman, N.; Klievink, J.T.; Blijenberg, B.; Aalvink, S.; Boeren, S.; Mank, M.; Knol, J.; de Vos, W.M. Akkermansia muciniphila uses human milk oligosaccharides to thrive in the early life conditions in vitro. Sci. Rep. 2020, 10, 14330. [Google Scholar] [CrossRef]
- Meng, X.; Wang, W.; Lan, T.; Yang, W.; Yu, D.; Fang, X.; Wu, H. A purified aspartic protease from Akkermansia muciniphila plays an important role in degrading Muc2. Int. J. Mol. Sci. 2019, 21, 72. [Google Scholar] [CrossRef]
- Zhao, Q.; Yu, J.; Hao, Y.; Zhou, H.; Hu, Y.; Zhang, C.; Zheng, H.; Wang, X.; Zeng, F.; Hu, J. Akkermansia muciniphila plays critical roles in host health. Crit. Rev. Microbiol. 2022, 1–19. [Google Scholar] [CrossRef]
- Ottman, N.; Geerlings, S.Y.; Aalvink, S.; de Vos, W.M.; Belzer, C. Action and function of Akkermansia muciniphila in microbiome ecology, health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 637–642. [Google Scholar] [CrossRef]
- Ekici, L.; Polat, H. Akkermansia muciniphila: Obezite ve Diyabetten Korunmada Yeni Bir Alternatif Olabilir mi? Avrupa Bilim Ve Teknol. Derg. 2019, 16, 533–543. [Google Scholar] [CrossRef]
- Zhai, Q.; Feng, S.; Arjan, N.; Chen, W. A next generation probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, W.; Wang, R.; Cheng, R.; Tang, Z.; Zhang, M. The outer membrane protein Amuc_1100 of Akkermansia muciniphila promotes intestinal 5-HT biosynthesis and extracellular availability through TLR2 signalling. Food Funct. 2021, 12, 3597–3610. [Google Scholar] [CrossRef]
- Mou, L.; Peng, X.; Chen, Y.; Xiao, Q.; Liao, H.; Liu, M.; Guo, L.; Liu, Y.; Zhang, X.; Deng, D. Crystal structure of monomeric Amuc_1100 from Akkermansia muciniphila. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2020, 76, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Fam, B.C.; Cempako, G.R.; Steinberg, G.R.; Walder, K.; Kay, T.W.; Proietto, J.; Andrikopoulos, S. Deficiency in interferon-γ results in reduced body weight and better glucose tolerance in mice. Endocrinology 2011, 152, 3690–3699. [Google Scholar] [CrossRef] [PubMed]
- Greer, R.L.; Dong, X.; Moraes, A.C.F.; Zielke, R.A.; Fernandes, G.R.; Peremyslova, E.; Vasquez-Perez, S.; Schoenborn, A.A.; Gomes, E.P.; Pereira, A.C. Akkermansia muciniphila mediates negative effects of IFNγ on glucose metabolism. Nat. Commun. 2016, 7, 13329. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, N.; Tan, H.-Y.; Li, S.; Zhang, C.; Feng, Y. Function of Akkermansia muciniphila in obesity: Interactions with lipid metabolism, immune response and gut systems. Front. Microbiol. 2020, 11, 219. [Google Scholar] [CrossRef]
- Abuqwider, J.N.; Mauriello, G.; Altamimi, M. Akkermansia muciniphila, a new generation of beneficial microbiota in modulating obesity: A systematic review. Microorganisms 2021, 9, 1098. [Google Scholar] [CrossRef]
- Zhang, J.; Ni, Y.; Qian, L.; Fang, Q.; Zheng, T.; Zhang, M.; Gao, Q.; Zhang, Y.; Ni, J.; Hou, X. Decreased abundance of Akkermansia muciniphila leads to the impairment of insulin secretion and glucose homeostasis in lean type 2 diabetes. Adv. Sci. 2021, 8, 2100536. [Google Scholar] [CrossRef]
- Cani, P.D.; Geurts, L.; Matamoros, S.; Plovier, H.; Duparc, T. Glucose metabolism: Focus on gut microbiota, the endocannabinoid system and beyond. Diabetes Metab. 2014, 40, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Wu, F.; Wang, X.; Feng, Y.; Wang, Y. MDG, an Ophiopogon japonicus polysaccharide, inhibits non-alcoholic fatty liver disease by regulating the abundance of Akkermansia muciniphila. Int. J. Biol. Macromol. 2022, 196, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, S.; Abbasi, A.; Somi, M.H.; Moaddab, S.Y.; Nikniaz, L.; Kafil, H.S.; Ebrahimzadeh Leylabadlo, H. Akkermansia muciniphila: From its critical role in human health to strategies for promoting its abundance in human gut microbiome. Crit. Rev. Food Sci. Nutr. 2022, 3, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Lee, J.-C.; Lee, H.-Y.; Kim, M.-S.; Whon, T.W.; Lee, M.-S.; Bae, J.-W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Cho, C.H.; Yun, M.S.; Jang, S.J.; You, H.J.; Kim, J.-h.; Han, D.; Cha, K.H.; Moon, S.H.; Lee, K. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat. Microbiol. 2021, 6, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Cassilly, C.D.; Liu, X.; Park, S.-M.; Tusi, B.K.; Chen, X.; Kwon, J.; Filipčík, P.; Bolze, A.S.; Liu, Z. Akkermansia muciniphila phospholipid induces homeostatic immune responses. Nature 2022, 608, 168–173. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kang, S.W.; Lee, J.H.; Park, S.-H.; Lee, J.-S. The evolution and competitive strategies of Akkermansia muciniphila in gut. Gut Microbes 2022, 14, 2025017. [Google Scholar] [CrossRef]
- Kumar, R.; Kane, H.; Wang, Q.; Hibberd, A.; Jensen, H.M.; Kim, H.-S.; Bak, S.Y.; Auzanneau, I.; Bry, S.; Christensen, N. Identification and Characterization of a Novel Species of Genus Akkermansia with Metabolic Health Effects in a Diet-Induced Obesity Mouse Model. Cells 2022, 11, 2084. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of heat-killed Mycobacterium setense manresensis as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. Eur. Food Saf. Auth. 2019, 17, e05824. [Google Scholar] [CrossRef]
- Johansen, M.D.; Kremer, L. CFTR depletion confers hypersusceptibility to Mycobacterium fortuitum in a zebrafish model. Front. Cell. Infect. Microbiol. 2020, 10, 357. [Google Scholar] [CrossRef]
- Zulu, M.; Monde, N.; Nkhoma, P.; Malama, S.; Munyeme, M. Nontuberculous mycobacteria in humans, animals, and water in Zambia: A systematic review. Front. Trop. Dis. 2021, 2, 9. [Google Scholar] [CrossRef]
- Pereira, A.C.; Ramos, B.; Reis, A.C.; Cunha, M.V. Non-tuberculous mycobacteria: Molecular and physiological bases of virulence and adaptation to ecological niches. Microorganisms 2020, 8, 1380. [Google Scholar] [CrossRef]
- Thornton, C.S.; Mellett, M.; Jarand, J.; Barss, L.; Field, S.K.; Fisher, D.A. The respiratory microbiome and nontuberculous mycobacteria: An emerging concern in human health. Eur. Respir. Rev. 2021, 30, 200299. [Google Scholar] [CrossRef]
- Lee, G.J.; Lee, H.-M.; Kim, T.S.; Kim, J.K.; Sohn, K.M.; Jo, E.-K. Mycobacterium fortuitum induces A20 expression that impairs macrophage inflammatory responses. Pathog. Dis. 2016, 74, ftw015. [Google Scholar] [CrossRef] [PubMed]
- Lamy, B.; Marchandin, H.; Hamitouche, K.; Laurent, F. Mycobacterium setense sp. nov., a Mycobacterium fortuitum-group organism isolated from a patient with soft tissue infection and osteitis. Int. J. Syst. Evol. Microbiol. 2008, 58, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Keikha, M. Case report of isolation of Mycobacterium setense from a hospital water supply. Environ. Dis. 2018, 3, 52. [Google Scholar] [CrossRef]
- Rech, G.; Vilaplana, C.; Velasco, J.; Pluvinet, R.; Santín, S.; Prat, C.; Julián, E.; Alcaide, F.; Comas, I.; Sumoy, L. Draft genome sequences of Mycobacterium setense type strain DSM-45070 and the nonpathogenic strain manresensis, isolated from the Bank of the Cardener River in Manresa, Catalonia, Spain. Genome Announc. 2015, 3, e01485-14. [Google Scholar] [CrossRef] [PubMed]
- Comberiati, P.; Di Cicco, M.; Paravati, F.; Pelosi, U.; Di Gangi, A.; Arasi, S.; Barni, S.; Caimmi, D.; Mastrorilli, C.; Licari, A. The role of gut and lung microbiota in susceptibility to tuberculosis. Int. J. Environ. Res. Public Health 2021, 18, 12220. [Google Scholar] [CrossRef]
- Koizumi, S.-i.; Wakita, D.; Sato, T.; Mitamura, R.; Izumo, T.; Shibata, H.; Kiso, Y.; Chamoto, K.; Togashi, Y.; Kitamura, H. Essential role of Toll-like receptors for dendritic cell and NK1. 1+ cell-dependent activation of type 1 immunity by Lactobacillus pentosus strain S-PT84. Immunol. Lett. 2008, 120, 14–19. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Cardona, P.; Marzo-Escartin, E.; Tapia, G.; Díaz, J.; García, V.; Varela, I.; Vilaplana, C.; Cardona, P.-J. Oral administration of heat-killed Mycobacterium manresensis delays progression toward active tuberculosis in C3HeB/FeJ mice. Front. Microbiol. 2016, 6, 1482. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Amaro, C.; Cardona, P.; Gassó, D.; Arias, L.; Velarde, R.; Tvarijonativiciute, A.; Serrano, E.; Cardona, P.-J. Protective Effect of Intestinal Helminthiasis Against Tuberculosis Progression Is Abrogated by Intermittent Food Deprivation. Front. Immunol. 2021, 12, 1233. [Google Scholar] [CrossRef] [PubMed]
- Tukvadze, N.; Cardona, P.; Vashakidze, S.; Shubladze, N.; Avaliani, Z.; Vilaplana, C.; Cardona, P.-J. Development of the food supplement Nyaditum resae as a new tool to reduce the risk of tuberculosis development. Int. J. Mycobacteriol. 2016, 5, S101–S102. [Google Scholar] [CrossRef]
- Cardona, P.-J. The progress of therapeutic vaccination with regard to tuberculosis. Front. Microbiol. 2016, 7, 1536. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Futagawa-Endo, Y.; Dicks, L.M. Isolation and characterization of fructophilic lactic acid bacteria from fructose-rich niches. Syst. Appl. Microbiol. 2009, 32, 593–600. [Google Scholar] [CrossRef]
- Endo, A. Fructophilic lactic acid bacteria inhabit fructose-rich niches in nature. Microb. Ecol. Health Dis. 2012, 23, 18563. [Google Scholar] [CrossRef]
- Muslimah, R.; Mahatmanto, T.; Kusnadi, J.; Murdiyatmo, U. General methods to isolate, characterize, select, and identify fructophilic lactic acid bacteria from fructose-rich environments—A mini-review. In Proceedings of the 2021 International Conference on Green Agro-industry and Bioeconomy, Malang, Indonesia, 6–7 July 2021; IOP Conference Series: Earth and Environmental Science. p. 012079. [Google Scholar]
- Pachla, A.; Wicha, M.; Ptaszyńska, A.A.; Borsuk, G.; Trokenheim, Ł.Ł.; Małek, W. The molecular and phenotypic characterization of fructophilic lactic acid bacteria isolated from the guts of Apis mellifera L. derived from a Polish apiary. J. Appl. Genet. 2018, 59, 503–514. [Google Scholar] [CrossRef]
- Asenjo, F.; Olmos, A.; Henríquez-Piskulich, P.; Polanco, V.; Aldea, P.; Ugalde, J.A.; Trombert, A.N. Genome sequencing and analysis of the first complete genome of Lactobacillus kunkeei strain MP2, an Apis mellifera gut isolate. PeerJ 2016, 4, e1950. [Google Scholar] [CrossRef]
- Butler, É.; Oien, R.F.; Lindholm, C.; Olofsson, T.C.; Nilson, B.; Vásquez, A. A pilot study investigating lactic acid bacterial symbionts from the honeybee in inhibiting human chronic wound pathogens. Int. Wound J. 2016, 13, 729–737. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Butler, È.; Markowicz, P.; Lindholm, C.; Larsson, L.; Vásquez, A. Lactic acid bacterial symbionts in honeybees–an unknown key to honey’s antimicrobial and therapeutic activities. Int. Wound J. 2016, 13, 668–679. [Google Scholar] [CrossRef]
- Dicks, L.; Endo, A. Are fructophilic lactic acid bacteria (FLAB) beneficial to humans? Benef. Microbes 2022, 13, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Ulusoy, E.; Kolayli, S. Phenolic composition and antioxidant properties of Anzer bee pollen. J. Food Biochem. 2014, 38, 73–82. [Google Scholar] [CrossRef]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc. Natl. Acad. Sci. USA 2013, 110, 8842–8846. [Google Scholar] [CrossRef] [PubMed]
- De Las Rivas, B.; Rodríguez, H.C.; Curiel, J.A.; Landete, J.M.A.; Munoz, R. Molecular screening of wine lactic acid bacteria degrading hydroxycinnamic acids. J. Agric. Food Chem. 2009, 57, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; de Paulis, T.; May, J.M. Antioxidant effects of dihydrocaffeic acid in human EA. hy926 endothelial cells. J. Nutr. Biochem. 2004, 15, 722–729. [Google Scholar] [CrossRef]
- Endo, A.; Tanizawa, Y.; Tanaka, N.; Maeno, S.; Kumar, H.; Shiwa, Y.; Okada, S.; Yoshikawa, H.; Dicks, L.; Nakagawa, J. Comparative genomics of Fructobacillus spp. and Leuconostoc spp. reveals niche-specific evolution of Fructobacillus spp. BMC Genom. 2015, 16, 1117. [Google Scholar] [CrossRef]
- Endo, A.; Okada, S. Reclassification of the genus Leuconostoc and proposals of Fructobacillus fructosus gen. nov., comb. nov., Fructobacillus durionis comb. nov., Fructobacillus ficulneus comb. nov. and Fructobacillus pseudoficulneus comb. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 2195–2205. [Google Scholar] [CrossRef]
- Gustaw, K.; Michalak, M.; Polak-Borecka, M.; Wasko, A. Fruktofilne bakterie kwasu mlekowego (FLAB)–Nowa grupa heterofermentatywnych mikroorganizmów ze środowiska roślinnego. Postępy Mikrobiol. 2017, 56, 56–66. [Google Scholar]
- Bautista-Gallego, J.; Medina, E.; Sánchez, B.; Benítez-Cabello, A.; Arroyo-López, F.N. Role of lactic acid bacteria in fermented vegetables. Grasas Aceites 2020, 71, e358. [Google Scholar] [CrossRef]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Rani, R.P. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef]
- Endo, A.; Maeno, S.; Tanizawa, Y.; Kneifel, W.; Arita, M.; Dicks, L.; Salminen, S. Fructophilic lactic acid bacteria, a unique group of fructose-fermenting microbes. Appl. Environ. Microbiol. 2018, 84, e01290-18. [Google Scholar] [CrossRef] [PubMed]
- Ispirli, H.; Dertli, E. Detection of fructophilic lactic acid bacteria (FLAB) in bee bread and bee pollen samples and determination of their functional roles. J. Food Process. Preserv. 2021, 45, e15414. [Google Scholar] [CrossRef]
- Neveling, D.P.; Endo, A.; Dicks, L.M. Fructophilic Lactobacillus kunkeei and Lactobacillus brevis isolated from fresh flowers, bees and bee-hives. Curr. Microbiol. 2012, 65, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Ganassi, S.; Albanese, G.; Letizia, F.; Testa, B.; Tedino, C.; Petrarca, S.; Mutinelli, F.; Mazzeo, A.; De Cristofaro, A. Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases. Vet. Sci. 2022, 9, 236. [Google Scholar] [CrossRef]
- Pachla, A.; Ptaszyńska, A.A.; Wıcha, M.; Oleńska, E.; Małek, W. Fascinating fructophilic lactic acid bacteria associated with various fructose-rich niches. Ann. Univ. Mariae Curie-Sklodowska Sect. C Biol. 2019, 72, 41–50. [Google Scholar] [CrossRef]
- Acín Albiac, M.; Di Cagno, R.; Filannino, P.; Cantatore, V.; Gobbetti, M. How fructophilic lactic acid bacteria may reduce the FODMAPs content in wheat-derived baked goods: A proof of concept. Microb. Cell Factories 2020, 19, 182. [Google Scholar] [CrossRef]
- Mtshali, P.S.; Divol, B.; Du Toit, M. Identification and characterization of Lactobacillus florum strains isolated from South African grape and wine samples. Int. J. Food Microbiol. 2012, 153, 106–113. [Google Scholar] [CrossRef]
- Endo, A.; Futagawa-Endo, Y.; Sakamoto, M.; Kitahara, M.; Dicks, L.M. Lactobacillus florum sp. nov., a fructophilic species isolated from flowers. Int. J. Syst. Evol. Microbiol. 2010, 60, 2478–2482. [Google Scholar] [CrossRef]
- Chuah, L.-O.; Shamila-Syuhada, A.K.; Liong, M.T.; Rosma, A.; Thong, K.L.; Rusul, G. Physio-chemical, microbiological properties of tempoyak and molecular characterisation of lactic acid bacteria isolated from tempoyak. Food Microbiol. 2016, 58, 95–104. [Google Scholar] [CrossRef]
- Ouattara, H.D.; Ouattara, H.G.; Droux, M.; Reverchon, S.; Nasser, W.; Niamke, S.L. Lactic acid bacteria involved in cocoa beans fermentation from Ivory Coast: Species diversity and citrate lyase production. Int. J. Food Microbiol. 2017, 256, 11–19. [Google Scholar] [CrossRef]
- Ruiz Rodríguez, L.G.; Aller, K.; Bru, E.; De Vuyst, L.; Hébert, E.M.; Mozzi, F. Enhanced mannitol biosynthesis by the fruit origin strain Fructobacillus tropaeoli CRL 2034. Appl. Microbiol. Biotechnol. 2017, 101, 6165–6177. [Google Scholar] [CrossRef] [PubMed]
- Viesser, J.A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Vandenberghe, L.P.d.S.; Azevedo, V.; Brenig, B.; Rogez, H.; Góes-Neto, A.; Soccol, C.R. Exploring the contribution of fructophilic lactic acid bacteria to cocoa beans fermentation: Isolation, selection and evaluation. Food Res. Int. 2020, 136, 109478. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Bangar, S.P.; Echegaray, N.; Suri, S.; Tomasevic, I.; Manuel Lorenzo, J.; Melekoglu, E.; Rocha, J.M.; Ozogul, F. The Impacts of Lactiplantibacillus plantarum on the Functional Properties of Fermented Foods: A Review of Current Knowledge. Microorganisms 2022, 10, 826. [Google Scholar] [CrossRef]
- Asama, T.; Arima, T.H.; Gomi, T.; Keishi, T.; Tani, H.; Kimura, Y.; Tatefuji, T.; Hashimoto, K. Lactobacillus kunkeei YB 38 from honeybee products enhances IgA production in healthy adults. J. Appl. Microbiol. 2015, 119, 818–826. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Tlais, A.Z.A.; Cantatore, V.; Gobbetti, M. Fructose-rich niches traced the evolution of lactic acid bacteria toward fructophilic species. Crit. Rev. Microbiol. 2019, 45, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Masood, M.I.; Qadir, M.I.; Shirazi, J.H.; Khan, I.U. Beneficial effects of lactic acid bacteria on human beings. Crit. Rev. Microbiol. 2011, 37, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Papalexandratou, Z.; Vrancken, G.; De Bruyne, K.; Vandamme, P.; De Vuyst, L. Spontaneous organic cocoa bean box fermentations in Brazil are characterized by a restricted species diversity of lactic acid bacteria and acetic acid bacteria. Food Microbiol. 2011, 28, 1326–1338. [Google Scholar] [CrossRef]
- Lefeber, T.; Gobert, W.; Vrancken, G.; Camu, N.; De Vuyst, L. Dynamics and species diversity of communities of lactic acid bacteria and acetic acid bacteria during spontaneous cocoa bean fermentation in vessels. Food Microbiol. 2011, 28, 457–464. [Google Scholar] [CrossRef]
- Vergalito, F.; Testa, B.; Cozzolino, A.; Letizia, F.; Succi, M.; Lombardi, S.J.; Tremonte, P.; Pannella, G.; Di Marco, R.; Sorrentino, E. Potential application of Apilactobacillus kunkeei for human use: Evaluation of probiotic and functional properties. Foods 2020, 9, 1535. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Kubow, S.; Sadiq, F.A. Isolation and in-vitro probiotic characterization of fructophilic lactic acid bacteria from Chinese fruits and flowers. Lwt 2019, 104, 70–75. [Google Scholar] [CrossRef]
- Takatani, N.; Endo, A. Viable fructophilic lactic acid bacteria present in honeybee-based food products. FEMS Microbiol. Lett. 2021, 368, fnab150. [Google Scholar] [CrossRef] [PubMed]
- Gallego, C.G.; Salminen, S. Novel probiotics and prebiotics: How can they help in human gut microbiota dysbiosis? Appl. Food Biotechnol. 2016, 3, 72–81. [Google Scholar]
- Slyepchenko, A.; Carvalho, A.F.; Cha, D.S.; Kasper, S.; McIntyre, R.S. Gut emotions-mechanisms of action of probiotics as novel therapeutic targets for depression and anxiety disorders. CNS Neurol. Disord. Drug Targets 2014, 13, 1770–1786. [Google Scholar] [CrossRef] [PubMed]
- Zommiti, M.; Feuilloley, M.G.; Connil, N. Update of probiotics in human world: A nonstop source of benefactions till the end of time. Microorganisms 2020, 8, 1907. [Google Scholar] [CrossRef]
- Sabbatini, S.; Monari, C.; Ballet, N.; Mosci, P.; Decherf, A.C.; Pélerin, F.; Perito, S.; Scarpelli, P.; Vecchiarelli, A. Saccharomyces cerevisiae–based probiotic as novel anti-microbial agent for therapy of bacterial vaginosis. Virulence 2018, 9, 954–966. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, G.; Xu, D.; Fan, L.; Wu, X.; Ni, X.; Zhao, S.; Zheng, P.; Sun, J.; Ma, Y. A Novel Corynebacterium glutamicum l-Glutamate Exporter. Appl. Environ. Microbiol. 2018, 84, e02691-17. [Google Scholar] [CrossRef]
- Wahyono, A.; Lee, S.-B.; Kang, W.-W.; Park, H.-D. Improving bread quality using co-cultures of Saccharomyces cerevisiae, Torulaspora delbrueckii JK08, and Pichia anomala JK04. Ital. J. Food Sci. 2016, 28, 298. [Google Scholar]
- Vinderola, G.; Ouwehand, A.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Mazhar, S.; Yasmeen, R.; Chaudhry, A.; Summia, K.; Hussain, I.; Amjad, S.; Ali, E. Role of Microorganisms in Modern Food Industry. Int. J. Food Sci. Technol. 2022, 4, 65–77. [Google Scholar]
- Macori, G.; Cotter, P.D. Novel insights into the microbiology of fermented dairy foods. Curr. Opin. Biotechnol. 2018, 49, 172–178. [Google Scholar] [CrossRef]
- Nemati, M.; Hamidi, A.; Dizaj, S.M.; Javaherzadeh, V.; Lotfipour, F. An overview on novel microbial determination methods in pharmaceutical and food quality control. Adv. Pharm. Bull. 2016, 6, 301. [Google Scholar] [CrossRef]
- Druart, C.; Plovier, H.; Van Hul, M.; Brient, A.; Phipps, K.R.; de Vos, W.M.; Cani, P.D. Toxicological safety evaluation of pasteurized Akkermansia muciniphila. J. Appl. Toxicol. 2021, 41, 276–290. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Fernández-Navarro, T.; Arboleya, S.; de Los Reyes-Gavilán, C.; Salazar, N.; Gueimonde, M. Fermented dairy foods: Impact on intestinal microbiota and health-linked biomarkers. Front. Microbiol. 2019, 10, 1046. [Google Scholar] [CrossRef]
- Li, Z.-T.; Zhu, L.; Hu, G.-A.; Sun, Z.-L.; Zhan, X.-B.; Gao, M.-J. Akkermansia muciniphila fermentation culture based on a novel bionic large intestine dynamic digestion model. Food Biosci. 2021, 43, 101260. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens; Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of pasteurised Akkermansia muciniphila as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06780. [Google Scholar] [PubMed]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens; Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of heat-killed Mycobacterium setense manresensis as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, e05824. [Google Scholar]
- Asama, T.; Kimura, Y.; Kono, T.; Tatefuji, T.; Hashimoto, K.; Benno, Y. Effects of heat-killed Lactobacillus kunkeei YB38 on human intestinal environment and bowel movement: A pilot study. Benef. Microbes 2016, 7, 337–344. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ağagündüz, D.; Yılmaz, B.; Koçak, T.; Altıntaş Başar, H.B.; Rocha, J.M.; Özoğul, F. Novel Candidate Microorganisms for Fermentation Technology: From Potential Benefits to Safety Issues. Foods 2022, 11, 3074. https://doi.org/10.3390/foods11193074
Ağagündüz D, Yılmaz B, Koçak T, Altıntaş Başar HB, Rocha JM, Özoğul F. Novel Candidate Microorganisms for Fermentation Technology: From Potential Benefits to Safety Issues. Foods. 2022; 11(19):3074. https://doi.org/10.3390/foods11193074
Chicago/Turabian StyleAğagündüz, Duygu, Birsen Yılmaz, Tevfik Koçak, Hilal Betül Altıntaş Başar, João Miguel Rocha, and Fatih Özoğul. 2022. "Novel Candidate Microorganisms for Fermentation Technology: From Potential Benefits to Safety Issues" Foods 11, no. 19: 3074. https://doi.org/10.3390/foods11193074
APA StyleAğagündüz, D., Yılmaz, B., Koçak, T., Altıntaş Başar, H. B., Rocha, J. M., & Özoğul, F. (2022). Novel Candidate Microorganisms for Fermentation Technology: From Potential Benefits to Safety Issues. Foods, 11(19), 3074. https://doi.org/10.3390/foods11193074