
Tartary Buckwheat (Fagopyrum tataricum) Ameliorates Lipid Metabolism Disorders and Gut Microbiota Dysbiosis in High-Fat Diet-Fed Mice


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
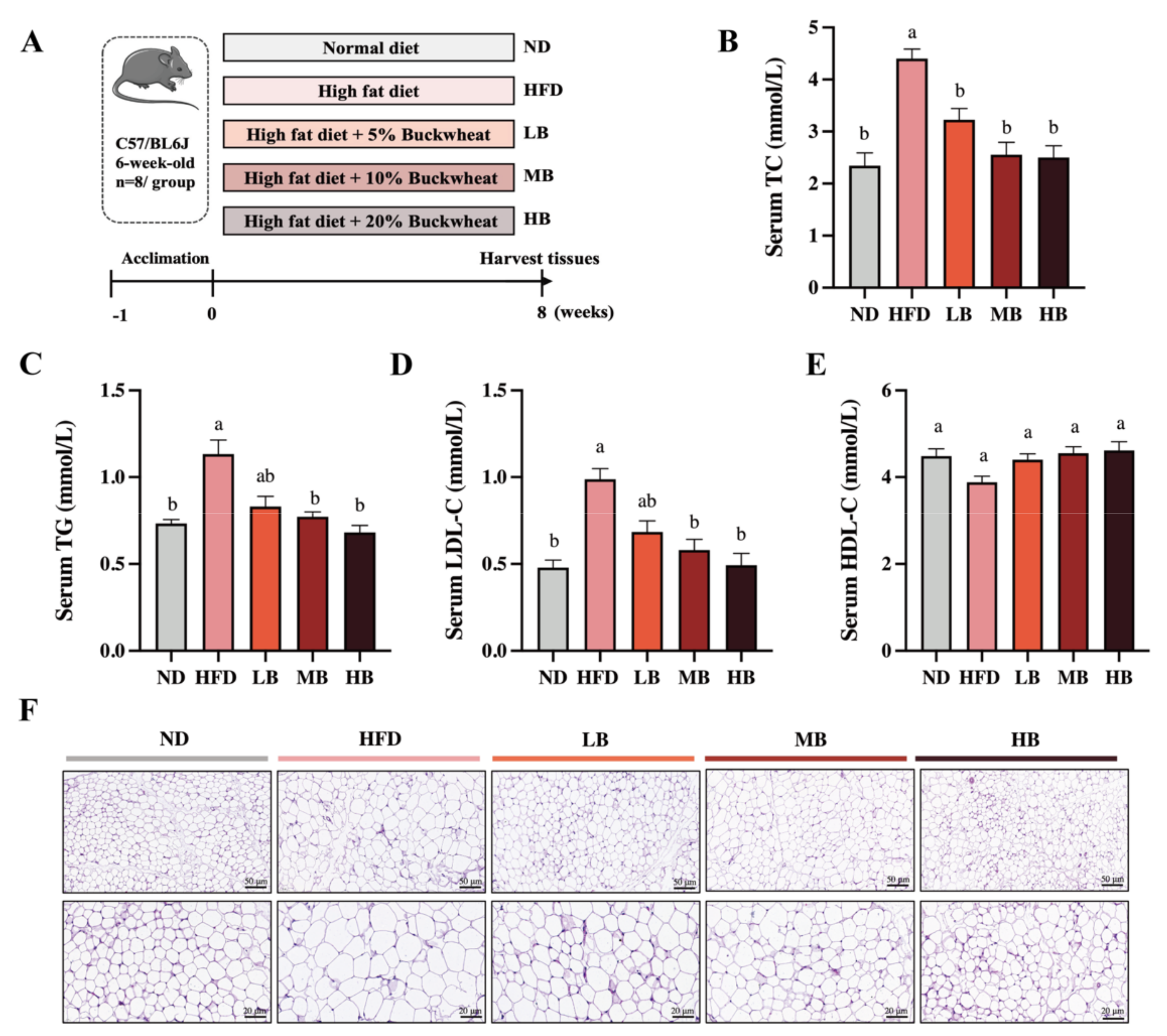
2.2. Experimental Animal Design
2.3. Biochemical Analysis
2.4. Histological Evaluation
2.5. Hormone Measurement
2.6. Gene Expression Analysis in the Liver
2.7. Western Blot
2.8. Gut Microbiota Analysis
2.9. Statistical Analysis
3. Results
3.1. Tartary Buckwheat Alleviated HFD-Induced Dyslipidemia and Lipid Accumulation
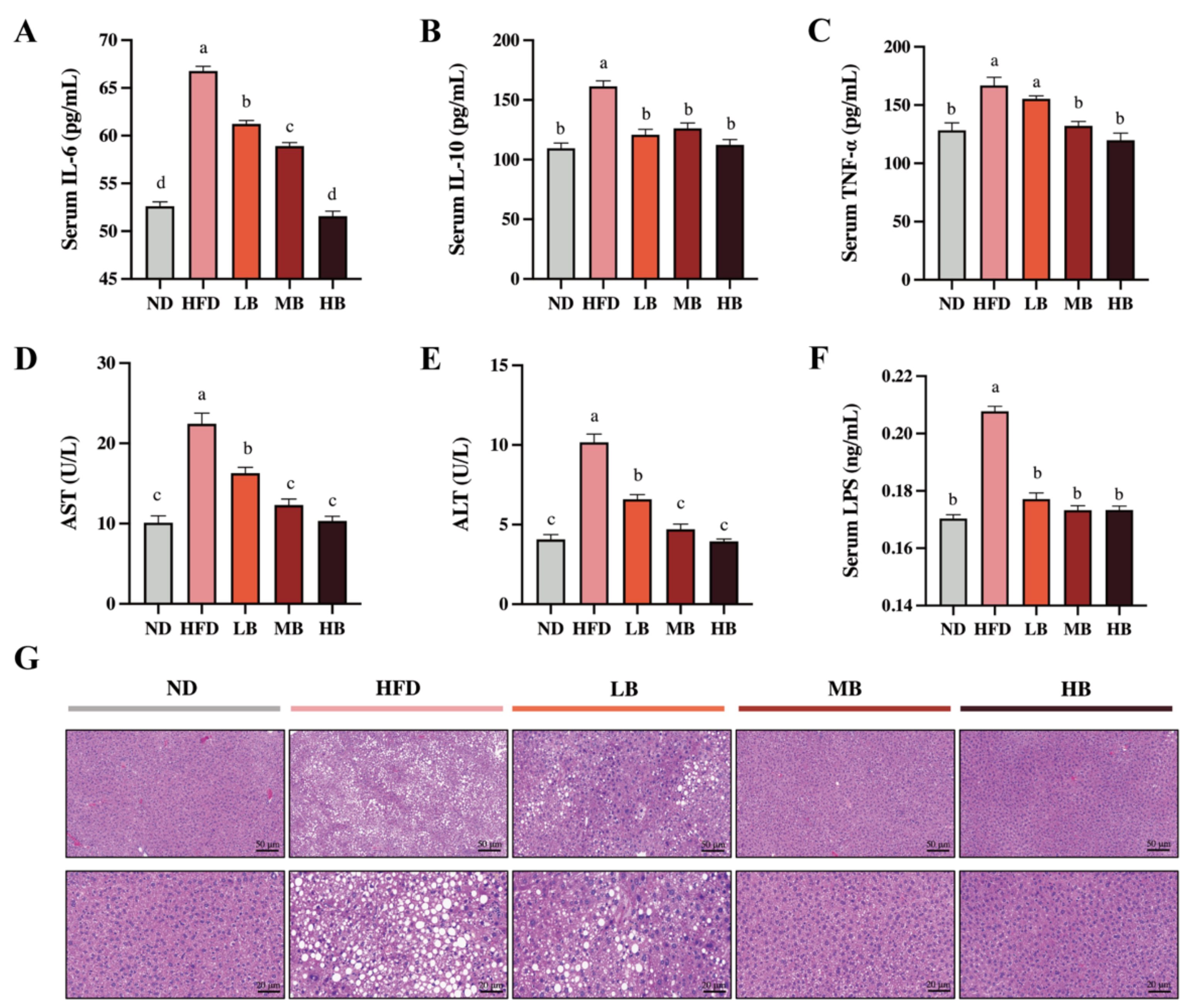
3.2. Tartary Buckwheat Attenuated HFD-Induced Low-Grade Inflammation and Hepatic Steatosis
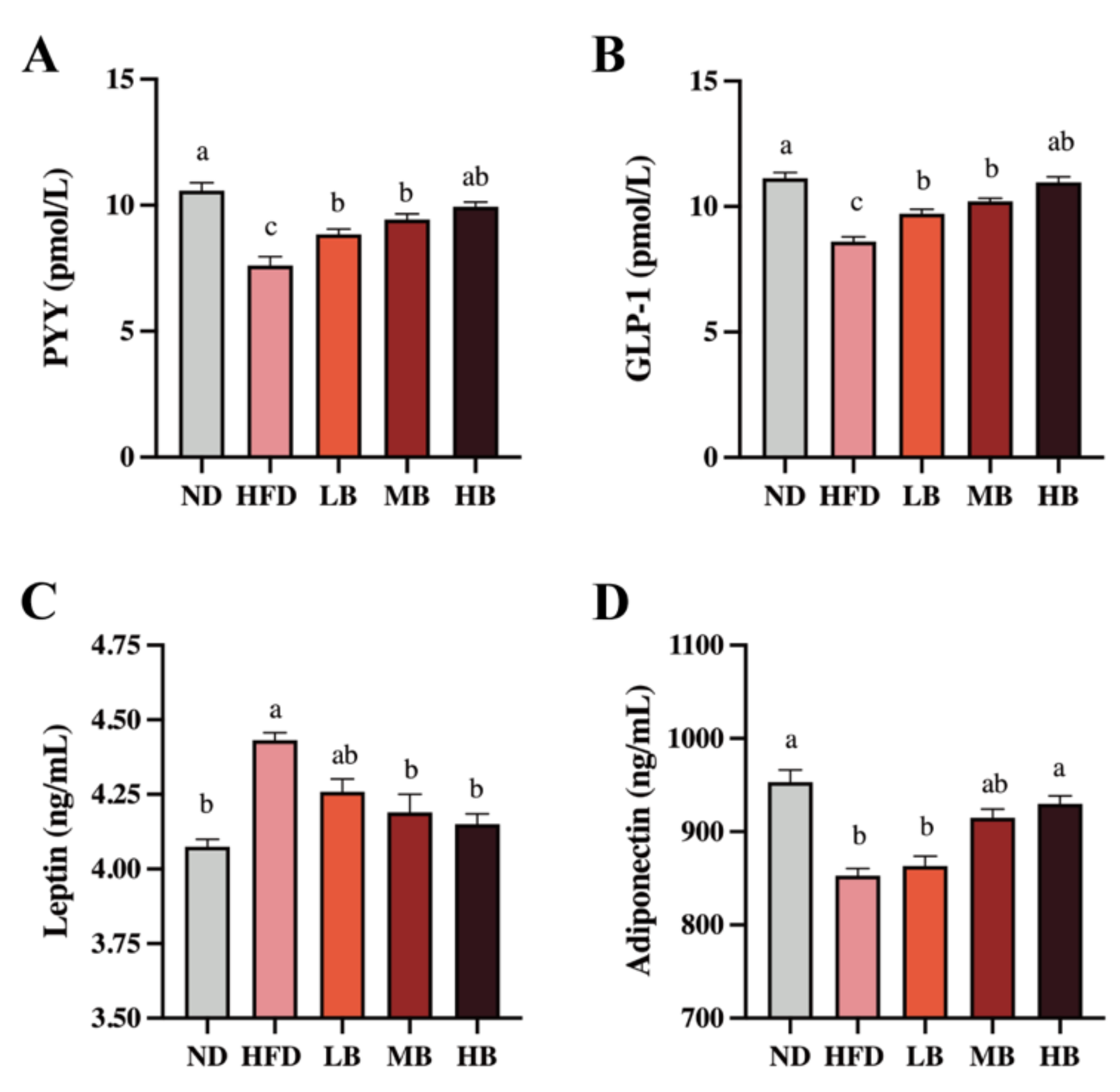
3.3. Tartary Buckwheat Modified the Circulating Gut Hormone in HFD-Fed Mice
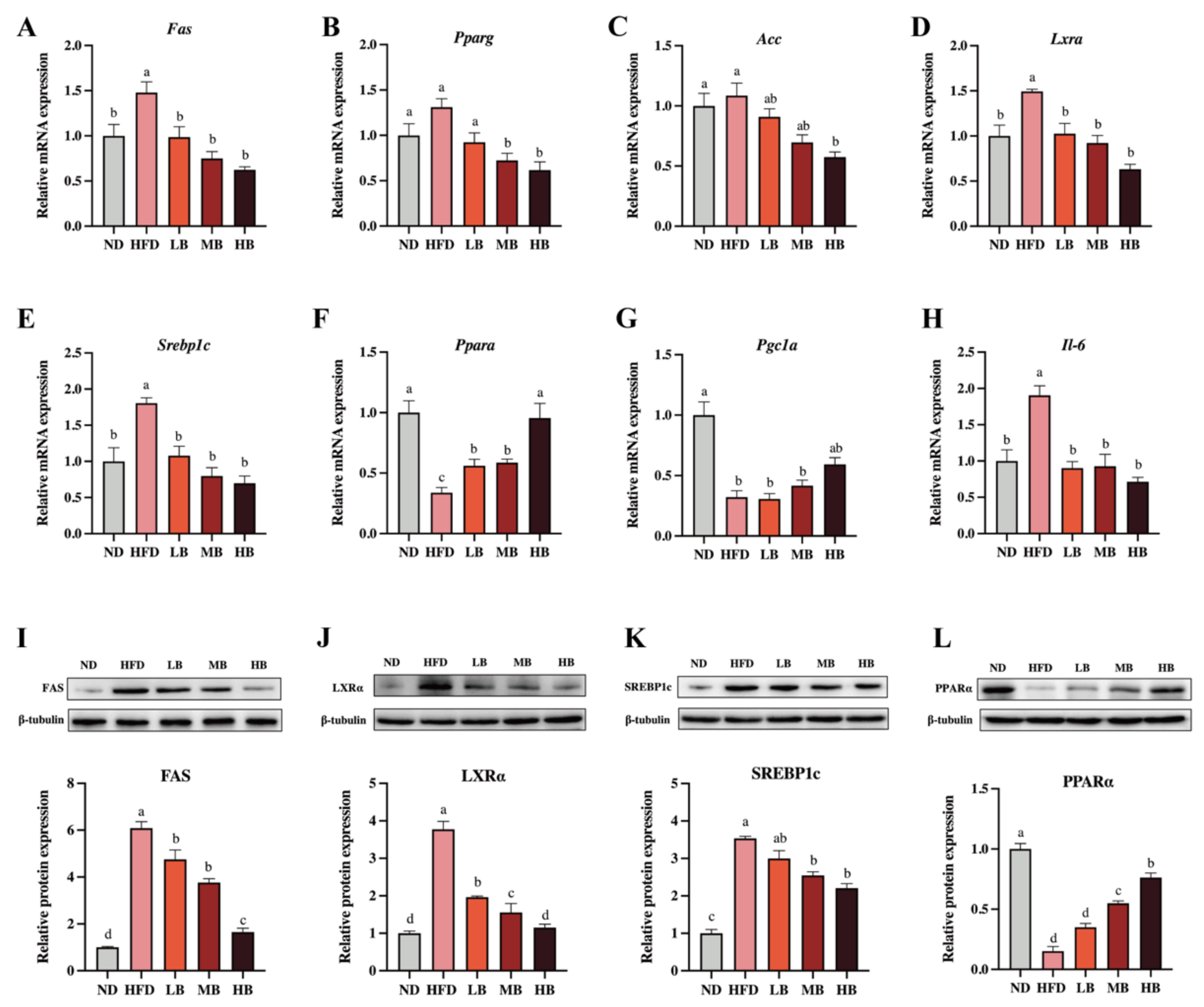
3.4. Tartary Buckwheat Regulated the Expression of Genes and Proteins Linked to Lipid Metabolism
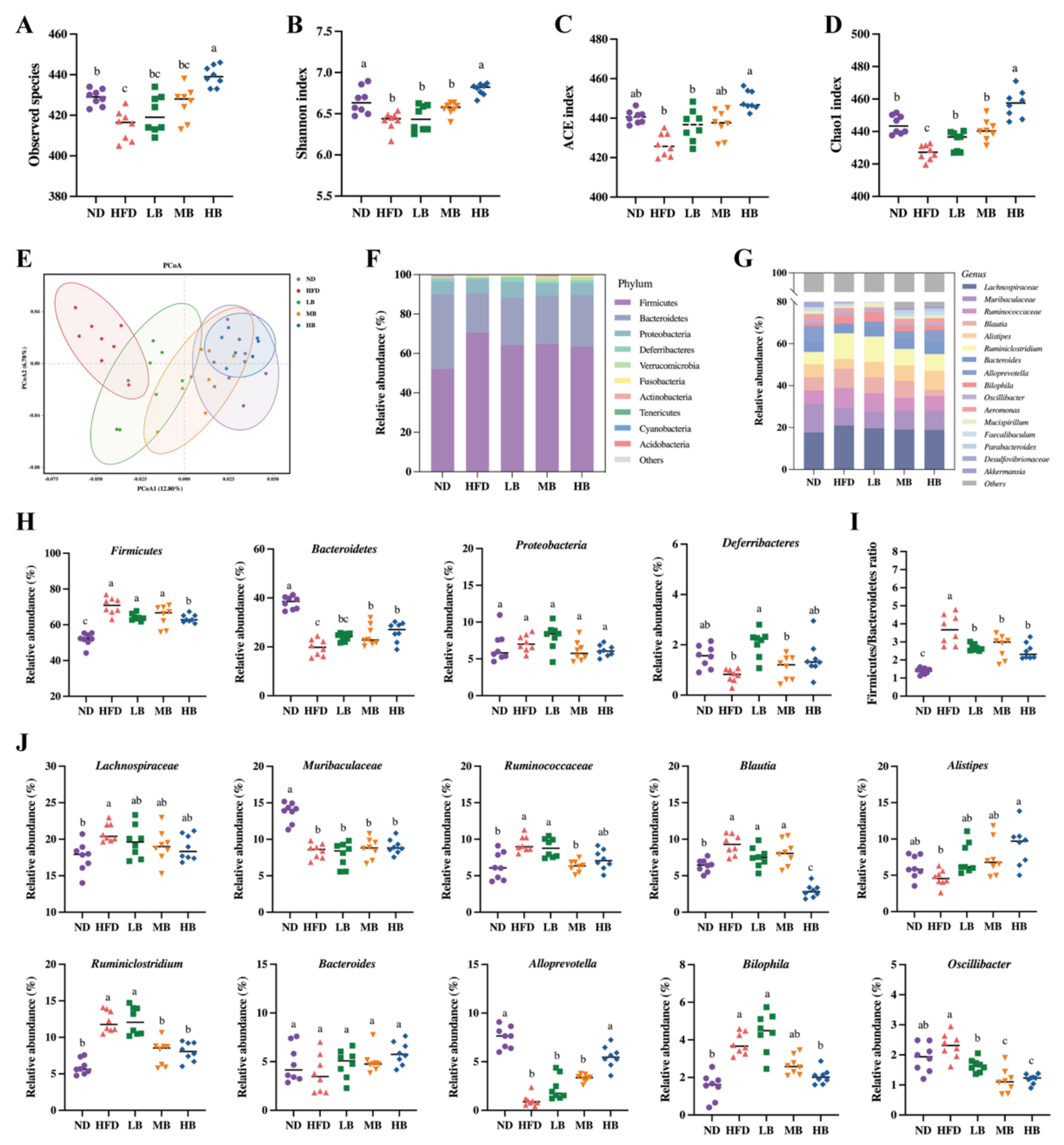
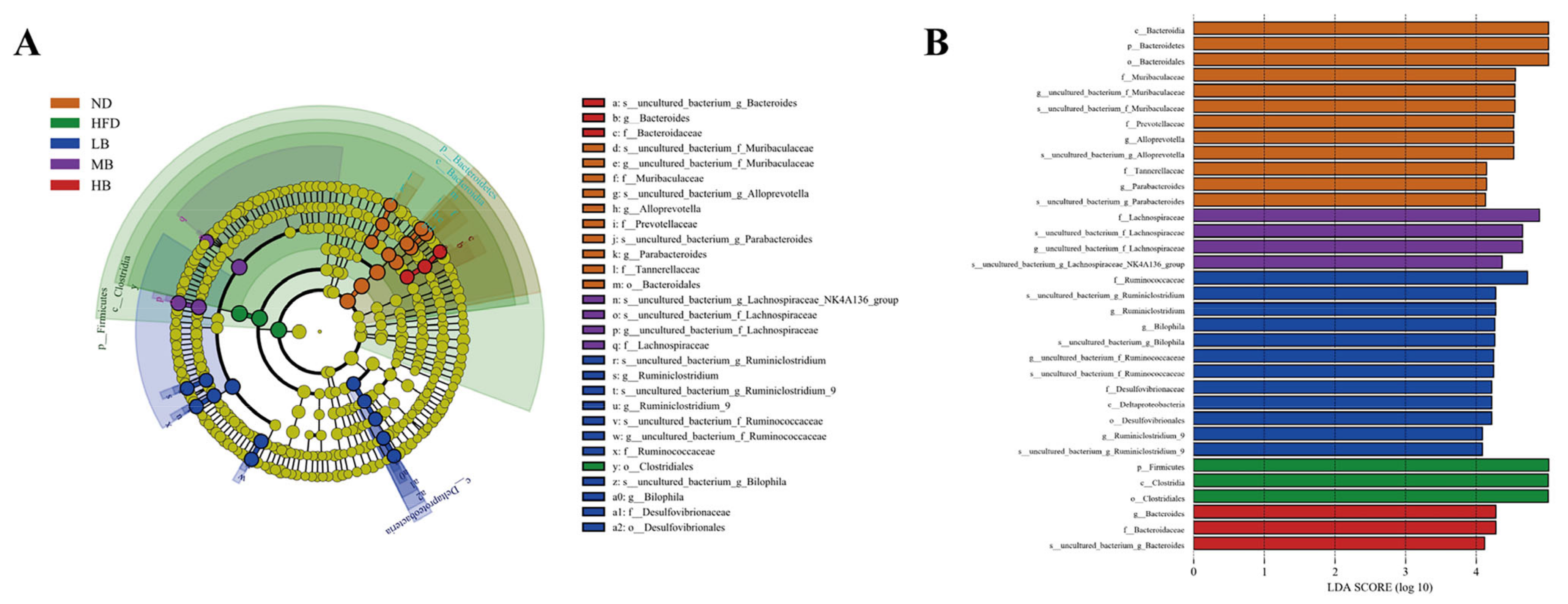
3.5. Tartary Buckwheat Modulated Gut Microbial Profile in HFD-Fed Mice
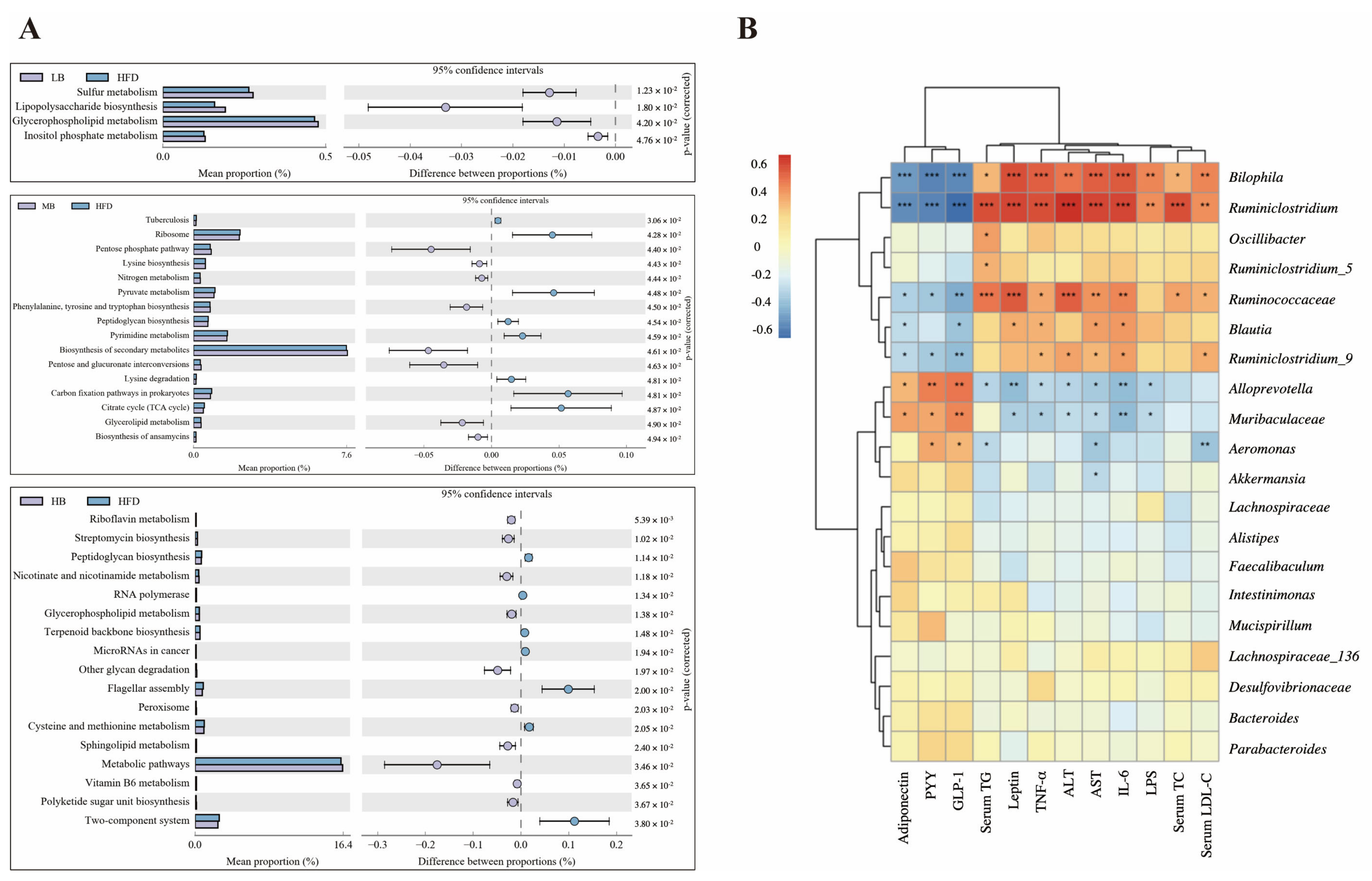
3.6. Predictive Function Profiling of the Gut Microbiome
3.7. Correlation Analysis between Gut Microbiota and Metabolic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Kishi, H.; Kobayashi, S. Add-on Therapy with Traditional Chinese Medicine: An Efficacious Approach for Lipid Metabolism Disorders. Pharmacol. Res. 2018, 134, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Montoro, J.I.; Damas-Fuentes, M.; Fernández-García, J.C.; Tinahones, F.J. Role of the Gut Microbiome in Beta Cell and Adipose Tissue Crosstalk: A Review. Front. Endocrinol. 2022, 13, 869951. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Liu, Q. Factors Affecting Gut Microbiome in Daily Diet. Front. Nutr. 2021, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lv, Q.; Liu, A.; Wang, J.; Sun, X.; Deng, J.; Chen, Q.; Wu, Q. Comparative Metabolomics Study of Tartary (Fagopyrum tataricum (L.) Gaertn) and Common (Fagopyrum esculentum Moench) Buckwheat Seeds. Food Chem. 2022, 371, 131125. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Song, Y.; Zhao, Q.; Wang, Y.; Li, C.; Zou, L.; Hu, Y. Effects of Tartary Buckwheat Protein on Gut Microbiome and Plasma Metabolite in Rats with High-Fat Diet. Foods 2021, 10, 2457. [Google Scholar] [CrossRef]
- Zhou, X.L.; Yan, B.B.; Xiao, Y.; Zhou, Y.M.; Liu, T.Y. Tartary Buckwheat Protein Prevented Dyslipidemia in High-Fat Diet-Fed Mice Associated with Gut Microbiota Changes. Food Chem. Toxicol. 2018, 119, 296–301. [Google Scholar] [CrossRef]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef]
- Bai, C.Z.; Feng, M.L.; Hao, X.L.; Zhong, Q.M.; Tong, L.G.; Wang, Z.H. Rutin, Quercetin, and Free Amino Acid Analysis in Buckwheat (Fagopyrum) Seeds from Different Locations. Genet. Mol. Res. 2015, 14, 19040–19048. [Google Scholar] [CrossRef]
- Tomotake, H.; Kayashita, J.; Kato, N. Hypolipidemic Activity of Common (Fagopyrum esculentum Moench) and Tartary (Fagopyrum tataricum Gaertn.) Buckwheat. J. Sci. Food Agric. 2015, 95, 1963–1967. [Google Scholar] [CrossRef]
- Feng, L.; Zhou, J.; Zhang, L.; Liu, P.; Zheng, P.; Gao, S.; Song, C.; Yu, Y.; Gong, Z.; Wan, X. Gut microbiota-mediated improvement of metabolic disorders by Qingzhuan tea in high fat diet-fed mice. J. Funct. Foods 2021, 78, 104366. [Google Scholar] [CrossRef]
- Cao, W.; Zhang, T.; Feng, R.; Xia, T.; Huang, H.; Liu, C.; Sun, C. Hoxa5 Alleviates Obesity-Induced Chronic Inflammation by Reducing ER Stress and Promoting M2 Macrophage Polarization in Mouse Adipose Tissue. J. Cell. Mol. Med. 2019, 23, 7029–7042. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Antony, V.; Sun, H.B.; Liang, G. Metabolism-Associated Molecular Patterns (MAMPs). Trends Endocrinol. Metab. 2020, 31, 712–724. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Zhao, F.; Cao, J.; Shen, X.; Li, C. Holothuria Leucospilota Polysaccharides Ameliorate Hyperlipidemia in High-Fat Diet-Induced Rats via Short-Chain Fatty Acids Production and Lipid Metabolism Regulation. Int. J. Mol. Sci. 2019, 20, 4738. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Chen, J.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. GOS Ameliorates Nonalcoholic Fatty Liver Disease Induced by High Fat and High Sugar Diet through Lipid Metabolism and Intestinal Microbes. Nutrients 2022, 14, 2749. [Google Scholar] [CrossRef]
- Wang, S.Y.; Cui, K.X.; Liu, J.H.; Hu, J.H.; Yan, K.; Xiao, P.; Lu, Y.Q.; Yang, X.G.; Liang, X.W. Mogroside-Rich Extract from Siraitia grosvenorii Fruits Ameliorates High-Fat Diet-Induced Obesity Associated with the Modulation of Gut Microbiota in Mice. Front. Nutr. 2022, 9, 1129. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.A.; Martin, A. Recent Insights into the Molecular Regulators and Mechanisms of Taurine to Modulate Lipid Metabolism: A Review. Crit. Rev. Food Sci. Nutr. 2022, 1–13. [Google Scholar] [CrossRef]
- Huang, Z.-R.; Chen, M.; Guo, W.-L.; Li, T.-T.; Liu, B.; Bai, W.-D.; Ai, L.-Z.; Rao, P.-F.; Ni, L.; Lv, X.-C. Monascus purpureus-fermented common buckwheat protects against dyslipidemia and non-alcoholic fatty liver disease through the regulation of liver metabolome and intestinal microbiome. Food Res. Int. 2020, 136, 109511. [Google Scholar] [CrossRef]
- Huang, Z.-R.; Deng, J.-C.; Li, Q.-Y.; Cao, Y.-J.; Lin, Y.-C.; Bai, W.-D.; Liu, B.; Rao, P.-F.; Ni, L.; Lv, X.-C. Protective Mechanism of Common Buckwheat (Fagopyrum esculentum Moench.) against Nonalcoholic Fatty Liver Disease Associated with Dyslipidemia in Mice Fed a High-Fat and High-Cholesterol Diet. J. Agric. Food Chem. 2020, 68, 6530–6543. [Google Scholar] [CrossRef]
- Feng, K.L.; Zhu, X.A.; Chen, T.; Peng, B.; Lu, M.W.; Zheng, H.; Huang, Q.R.; Ho, C.T.; Chen, Y.J.; Cao, Y. Prevention of Obesity and Hyperlipidemia by Heptamethoxyflavone in High-Fat Diet-induced Rats. J. Agric. Food Chem. 2019, 67, 2476–2489. [Google Scholar] [CrossRef]
- Peng, L.X.; Zhang, Q.; Zhang, Y.H.; Yao, Z.D.; Song, P.P.; Wei, L.J.; Zhao, G.; Yan, Z.Y. Effect of Tartary Buckwheat, Rutin, and Quercetin on Lipid Metabolism in Rats during High Dietary Fat Intake. Food Sci. Nutr. 2020, 8, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Farhadipour, M.; Depoortere, I. The Function of Gastrointestinal Hormones in Obesity—Implications for the Regulation of Energy Intake. Nutrients 2021, 13, 1839. [Google Scholar] [CrossRef] [PubMed]
- Teeuwisse, W.M.; Widya, R.L.; Paulides, M.; Lamb, H.J.; Smit, J.W.A.; de Roos, A.; van Buchem, M.A.; Pijl, H.; van der Grond, J. Short-Term Caloric Restriction Normalizes Hypothalamic Neuronal Responsiveness to Glucose Ingestion in Patients with Type 2 Diabetes. Diabetes 2012, 61, 3255–3259. [Google Scholar] [CrossRef]
- Krishnan, N.; Bonham, C.A.; Rus, I.A.; Shrestha, O.K.; Gauss, C.M.; Haque, A.; Tocilj, A.; Joshua-Tor, L.; Tonks, N.K. Harnessing Insulin- and Leptin-Induced Oxidation of PTP1B for Therapeutic Development. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 12, 585887. [Google Scholar] [CrossRef] [PubMed]
- El-Shinnawy, N.A.; Abd Elhalem, S.S.; Haggag, N.Z.; Badr, G. Ameliorative Role of Camel Whey Protein and Rosuvastatin on Induced Dyslipidemia in Mice. Food Funct. 2018, 9, 1038–1047. [Google Scholar] [CrossRef]
- Zhou, J.; Martin, R.J.; Raggio, A.M.; Shen, L.; McCutcheon, K.; Keenan, M.J. The Importance of GLP-1 and PYY in Resistant Starch’s Effect on Body Fat in Mice. Mol. Nutr. Food Res. 2015, 59, 1000–1003. [Google Scholar] [CrossRef]
- Kim, K.; Bae, G.D.; Lee, M.; Park, E.Y.; Baek, D.J.; Kim, C.Y.; Jun, H.S.; Oh, Y.S. Allomyrina Dichotoma Larva Extract Ameliorates the Hepatic Insulin Resistance of High-Fat Diet-Induced Diabetic Mice. Nutrients 2019, 11, 1522. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.C.; Xu, K.; Guo, Y.N.; Ping, L.; Gao, Y.Q.; Qiu, Y.; Ni, J.Q.; Liu, Q.F.; Wang, Z. A High-Fat Diet Reverses Metabolic Disorders and Premature Aging by Modulating Insulin and IGF1 Signaling in SIRT6 Knockout Mice. Aging Cell 2020, 19, e13104. [Google Scholar] [CrossRef] [PubMed]
- Aibara, D.; Matsusue, K.; Takiguchi, S.; Gonzalez, F.J.; Yamano, S. Fat-specific Protein 27 Is a Novel Target Gene of Liver X Receptor Alpha. Mol. Cell. Endocrinol. 2018, 474, 48–56. [Google Scholar] [CrossRef]
- Chen, G.; Wang, H.; Zhang, X.; Yang, S.T. Nutraceuticals and Functional Foods in the Management of Hyperlipidemia. Crit. Rev. Food Sci. Nutr. 2014, 54, 1180–1201. [Google Scholar] [CrossRef]
- Wang, Y.; Fei, Y.Q.; Liu, L.R.; Xiao, Y.H.; Pang, Y.L.; Kang, J.H.; Wang, Z. Polygonatum odoratum Polysaccharides Modulate Gut Microbiota and Mitigate Experimentally Induced Obesity in Rats. Int. J. Mol. Sci. 2018, 19, 3587. [Google Scholar] [CrossRef] [PubMed]
- Jun, H.J.; Joshi, Y.; Patil, Y.; Noland, R.C.; Chang, J.S. NT-PGC-1 alpha Activation Attenuates High-Fat Diet-Induced Obesity by Enhancing Brown Fat Thermogenesis and Adipose Tissue Oxidative Metabolism. Diabetes 2014, 63, 3615–3625. [Google Scholar] [CrossRef]
- Yan, J.; Xue, Q.Y.; Chen, W.Y.; Wang, K.; Peng, D.; Jiang, J.J.; Li, P.; Du, B. Probiotic-fermented Rice Buckwheat Alleviates High-fat diet-Induced Hyperlipidemia in Mice by Suppressing Lipid Accumulation and Modulating Gut Microbiota. Food Res. Int. 2022, 155, 111125. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.Y.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut Microbiome Composition Is Linked to Whole Grain-Induced Immunological Improvements. ISME J. 2013, 7, 269–280. [Google Scholar] [CrossRef]
- Cheng, R.Y.; Guo, J.W.; Pu, F.F.; Wan, C.M.; Shi, L.; Li, H.W.; Yang, Y.H.; Huang, C.Y.; Li, M.; He, F. Loading Ceftriaxone, Vancomycin, and Bifidobacteria Bifidum TMC3115 to Neonatal Mice Could Differently and Consequently Affect Intestinal Microbiota and Immunity in Adulthood. Sci. Rep. 2019, 9, 3254. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Yamada, H.; Motoyama, S.; Saburi, M.; Sugimoto, T.; Kubota, H.; Miyawaki, D.; Wakana, N.; Kami, D.; Ogata, T.; et al. Maternal High-fat Diet Exaggerates Diet -Induced Insulin Resistance in Adult Offspring by Enhancing in Flammasome Activation through Noncanonical Pathway of Caspase-11. Mol. Metab. 2020, 37, 100988. [Google Scholar] [CrossRef] [PubMed]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.L.; Rainteau, D.; Bridonneau, C.; da Costa, G.; Vlieg, J.V.; Sovran, B.; Chamignon, C.; et al. Bilophila Wadsworthia Aggravates High Fat Diet Induced Metabolic Dysfunctions in Mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Wu, M.N.; Li, P.Z.; An, Y.Y.; Ren, J.; Yan, D.; Cui, J.Z.; Li, D.; Li, M.; Wang, M.Y.; Zhong, G.S. Phloretin Ameliorates Dextran Sulfate Sodium-induced Ulcerative Colitis in Mice by Regulating the Gut Microbiota. Pharmacol. Res. 2019, 150, 104489. [Google Scholar] [CrossRef]
- Ran, B.B.; Guo, C.E.; Li, W.D.; Li, W.S.; Wang, Q.; Qian, J.X.; Li, H.L. Sea Buckthorn (Hippophae rhamnoides L.) Fermentation Liquid Protects against Alcoholic Liver Disease Linked to Regulation of Liver Metabolome and the Abundance of Gut Microbiota. J. Sci. Food Agric. 2021, 101, 2846–2854. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Galvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and Cultivation Study of Muribaculaceae Reveals Novel Species, Host Preference, and Functional Potential of This Yet Undescribed Family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, Q.; Ma, W.N.; Tian, F.; Shen, H.Y.; Zhou, M.M. A Combination of Quercetin and Resveratrol Reduces Obesity in High-Fat Diet-Fed Rats by Modulation of Gut Microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef] [PubMed]
- Pataky, Z.; Genton, L.; Spahr, L.; Lazarevic, V.; Terraz, S.; Gaia, N.; Rubbia-Brandt, L.; Golay, A.; Schrenzel, J.; Pichard, C. Impact of Hypocaloric Hyperproteic Diet on Gut Microbiota in Overweight or Obese Patients with Nonalcoholic Fatty Liver Disease: A Pilot Study. Dig. Dis. Sci. 2016, 61, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yue, S.J.; Yang, Z.R.; Feng, W.W.; Meng, X.T.; Wang, A.T.; Peng, C.; Wang, C.Y.; Yan, D. Oral Hydroxysafflor Yellow A Reduces Obesity in Mice by Modulating the Gut Microbiota and Serum Metabolism. Pharmacol. Res. 2018, 134, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut Microbial Metabolites in Obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef]
- Zhao, Q.Y.; Liu, Z.Y.; Zhu, Y.Q.; Wang, H.; Dai, Z.J.; Yang, X.H.; Ren, X.; Xue, Y.; Shen, Q. Cooked Adzuki Bean Reduces High-Fat Diet-Induced Body Weight Gain, Ameliorates Inflammation, and Modulates Intestinal Homeostasis in Mice. Front. Nutr. 2022, 9, 1205. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Group | Body | Liver | Food Intake | |||
|---|---|---|---|---|---|---|
| Final Weight (g) | Weight Gain (g) | Weight (g) | Ratio (%) | Consumption (g/d) | Energy Intake (kcal/d) | |
| ND | 23.62 ± 0.68 b 1 | 6.20 ± 1.45 b | 0.72 ± 0.02 b | 2.64 ± 0.10 b | 2.61 ± 0.16 a | 10.05 ± 0.62 b |
| HFD | 31.32 ± 1.11 a | 14.24 ± 0.84 a | 1.40 ± 0.02 a | 4.48 ± 0.07 a | 2.43 ± 0.19 a | 12.73 ± 0.98 a |
| LB | 27.02 ± 0.23 a | 10.20 ± 0.33 ab | 0.93 ± 0.04 b | 3.38 ± 0.19 b | 2.44 ± 0.06 a | 12.79 ± 0.29 a |
| MB | 24.98 ± 0.69 ab | 7.98 ± 1.39 ab | 0.86 ± 0.02 b | 3.47 ± 0.09 b | 2.38 ± 0.10 a | 12.47 ± 0.51 a |
| HB | 23.05 ± 0.49 b | 6.45 ± 1.01 b | 0.73 ± 0.03 b | 3.19 ± 0.14 b | 2.24 ± 0.16 a | 11.74 ± 0.82 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Wang, J.; Wang, Y.; Zhang, B.; Chen, Z.; Zhu, J.; Wang, X.; Wang, S. Tartary Buckwheat (Fagopyrum tataricum) Ameliorates Lipid Metabolism Disorders and Gut Microbiota Dysbiosis in High-Fat Diet-Fed Mice. Foods 2022, 11, 3028. https://doi.org/10.3390/foods11193028
Li A, Wang J, Wang Y, Zhang B, Chen Z, Zhu J, Wang X, Wang S. Tartary Buckwheat (Fagopyrum tataricum) Ameliorates Lipid Metabolism Disorders and Gut Microbiota Dysbiosis in High-Fat Diet-Fed Mice. Foods. 2022; 11(19):3028. https://doi.org/10.3390/foods11193028
Chicago/Turabian StyleLi, Ang, Jin Wang, Yuanyifei Wang, Bowei Zhang, Zhenjia Chen, Junling Zhu, Xiaowen Wang, and Shuo Wang. 2022. "Tartary Buckwheat (Fagopyrum tataricum) Ameliorates Lipid Metabolism Disorders and Gut Microbiota Dysbiosis in High-Fat Diet-Fed Mice" Foods 11, no. 19: 3028. https://doi.org/10.3390/foods11193028
APA StyleLi, A., Wang, J., Wang, Y., Zhang, B., Chen, Z., Zhu, J., Wang, X., & Wang, S. (2022). Tartary Buckwheat (Fagopyrum tataricum) Ameliorates Lipid Metabolism Disorders and Gut Microbiota Dysbiosis in High-Fat Diet-Fed Mice. Foods, 11(19), 3028. https://doi.org/10.3390/foods11193028