
Optimization of Fermentation Conditions of Artemisia capillaris for Enhanced Acetylcholinesterase and Butyrylcholinesterase


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preliminary Experiments for Fermentation Conditions
2.3. Box–Behnken Experiment Design
2.4. Fermentation
2.5. Acetylcholinesterase (AChE) and Butyrylcholinesterase (BuChE) Inhibitory Activity
Ssample: Slope of the sample
Sblank: Slope of the control
2.6. Antioxidant Activity
2.6.1. DPPH Radical Scavenging Activity
Acontrol: Absorbance of the control
Asample: Absorbance of the sample
2.6.2. Reducing Power
2.6.3. Ferric Reducing Antioxidant Power (FRAP)
2.7. Total Polyphenol Contents
2.8. Statistical Analysis
3. Results and Discussion
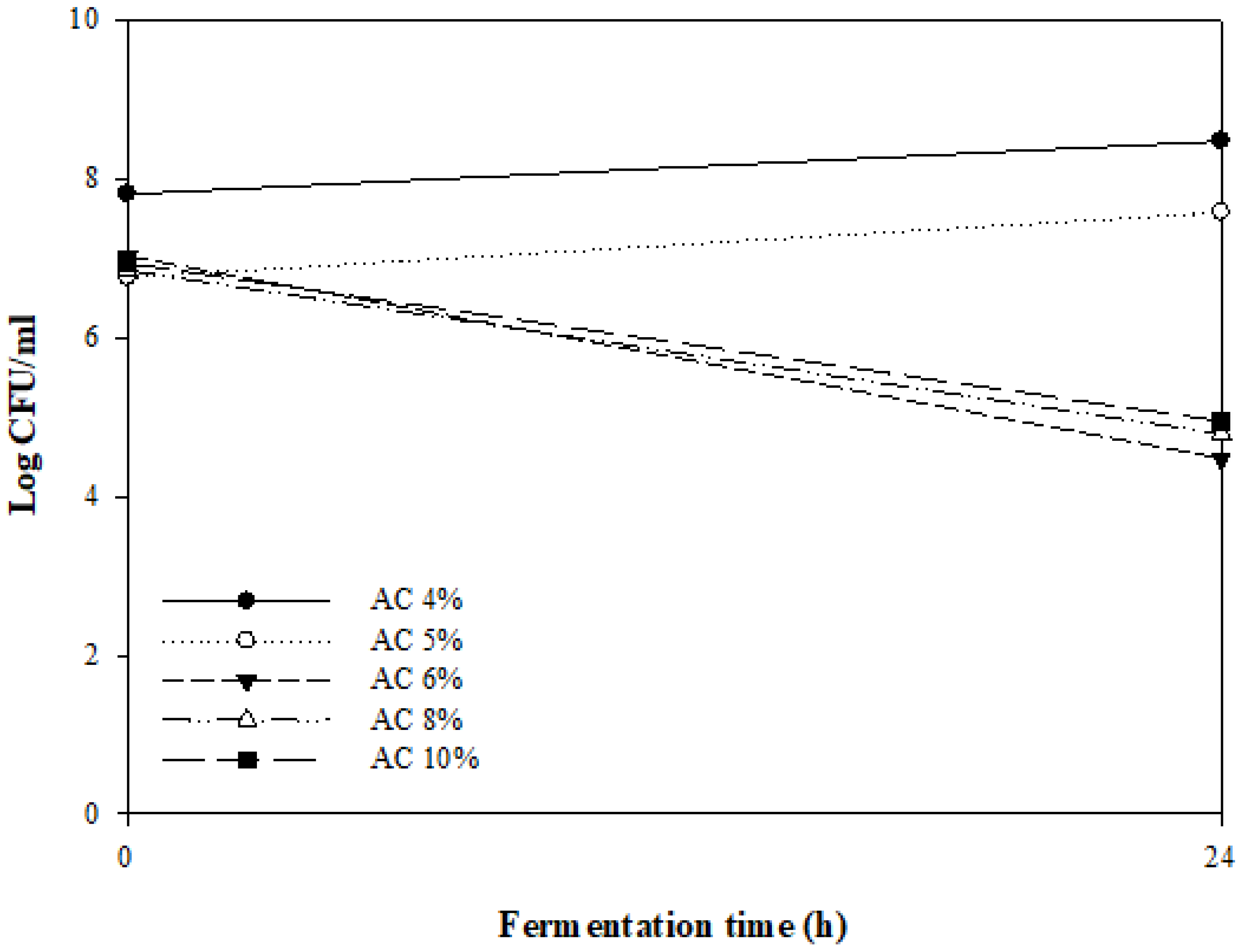
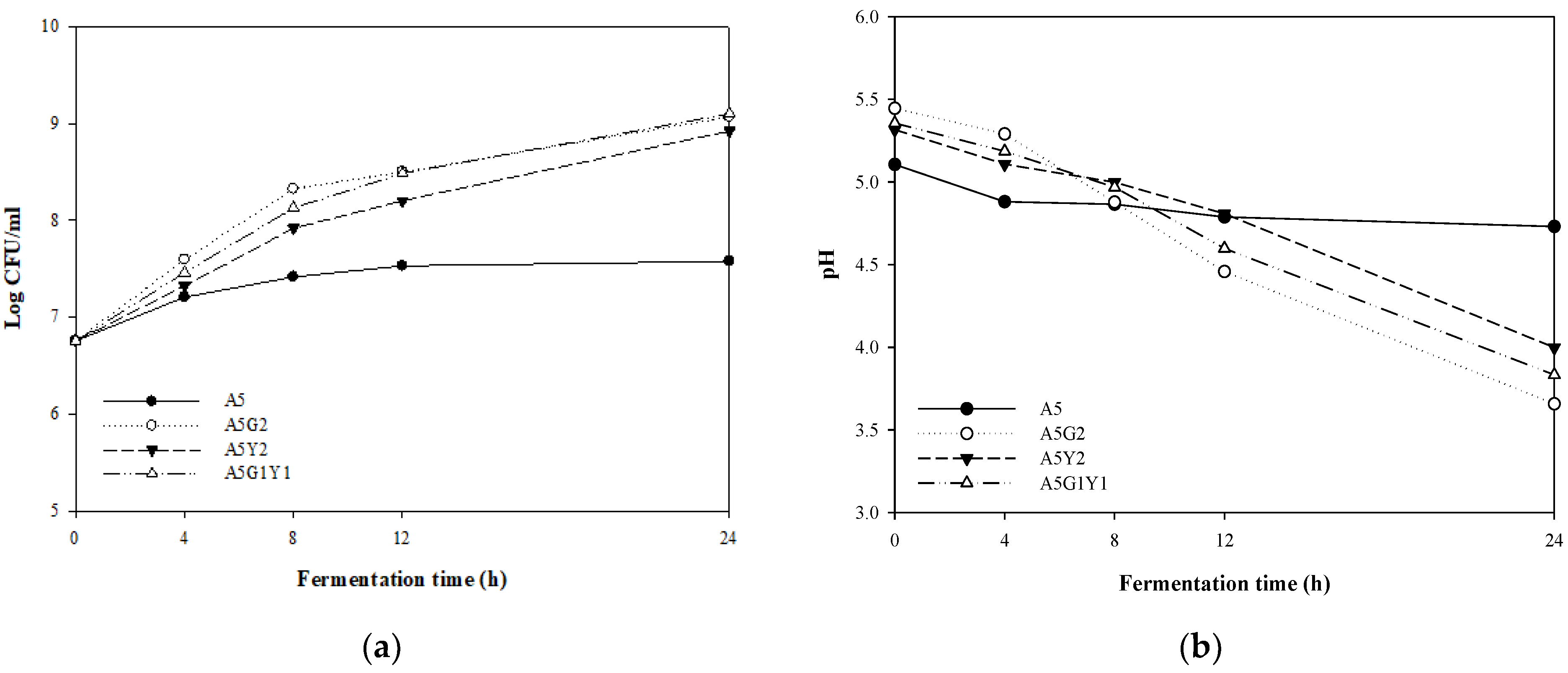
3.1. Preliminary Experiments for Fermentation Conditions
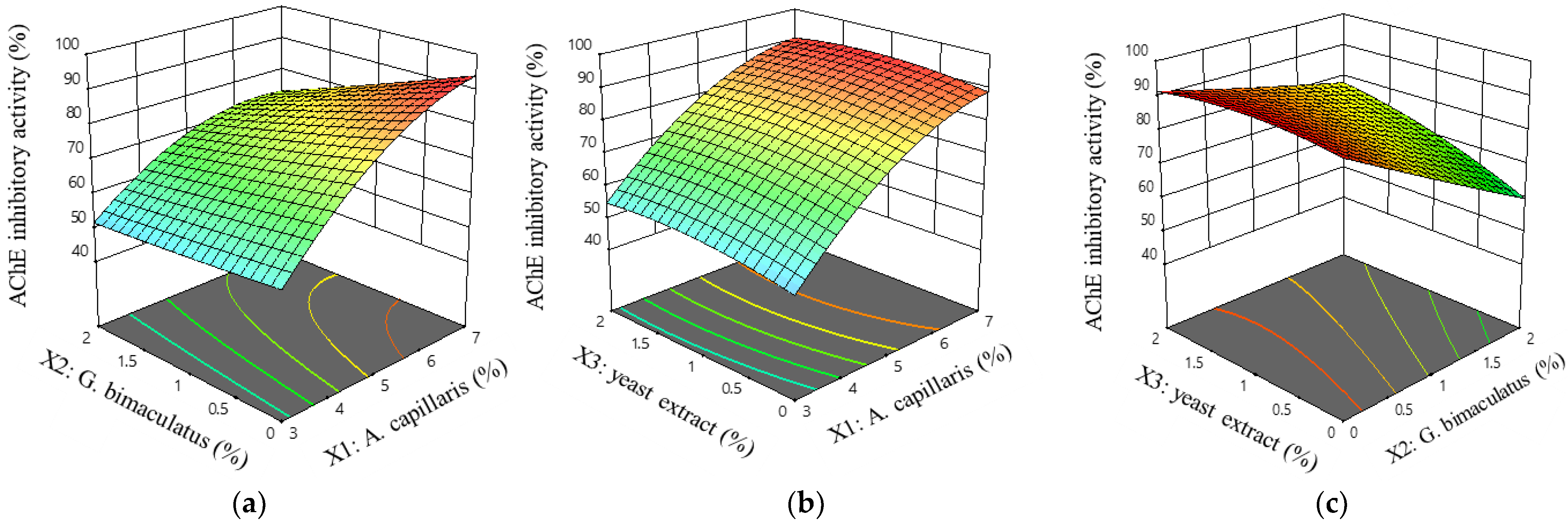
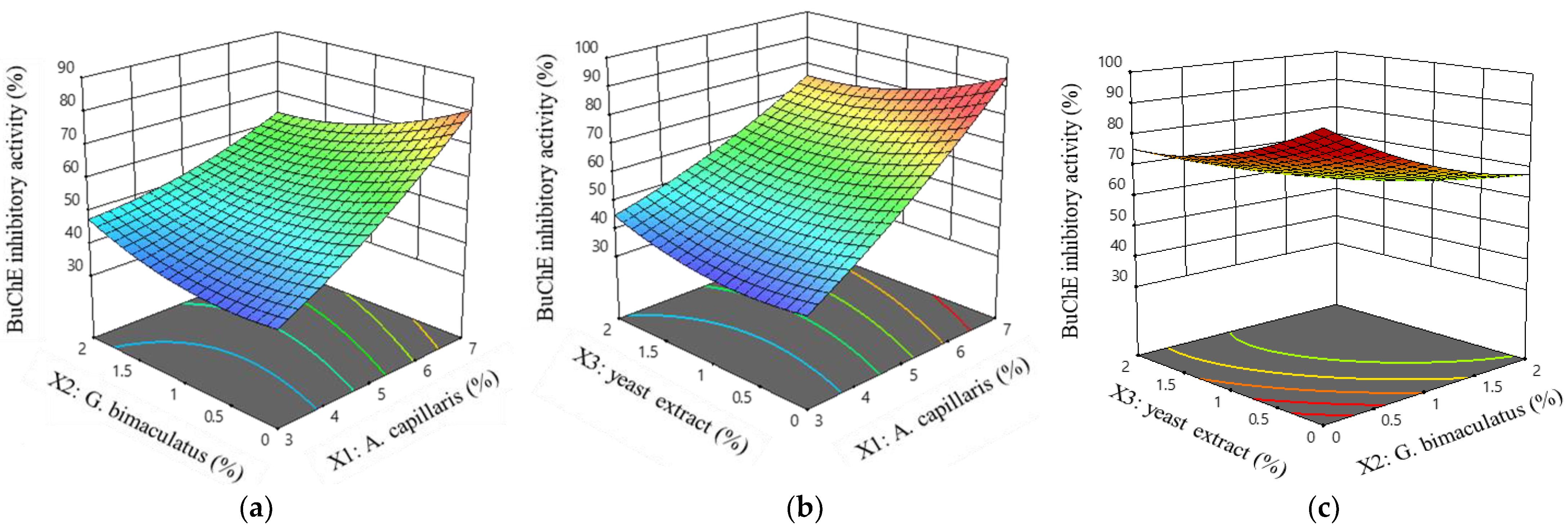
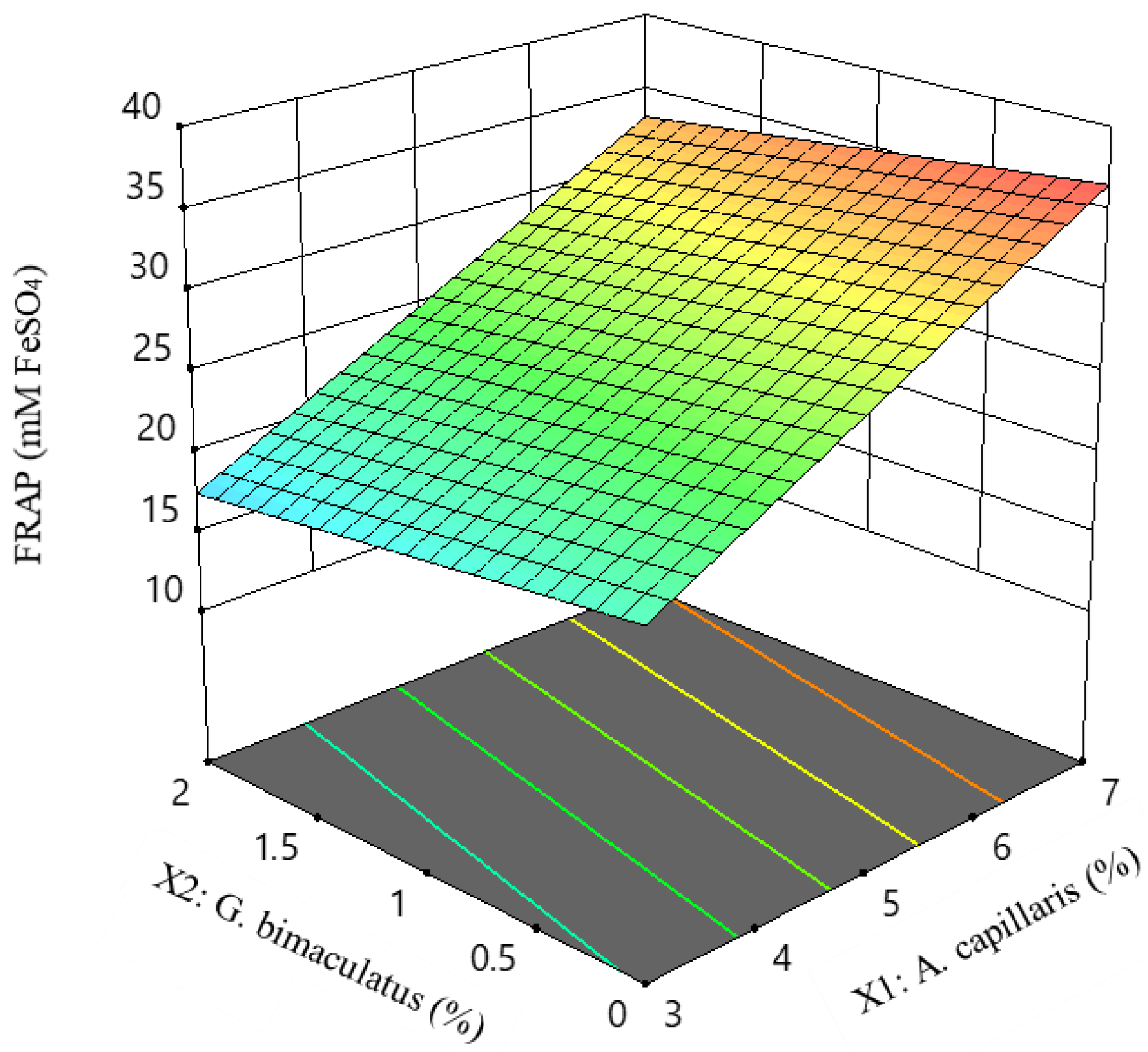
3.2. Experiment Design for the Optimal Condition of A. capillaris Fermentation
3.2.1. Microbiological and Physiological Analysis of Fermented A. capillaris
3.2.2. Suitability of the Regression Model
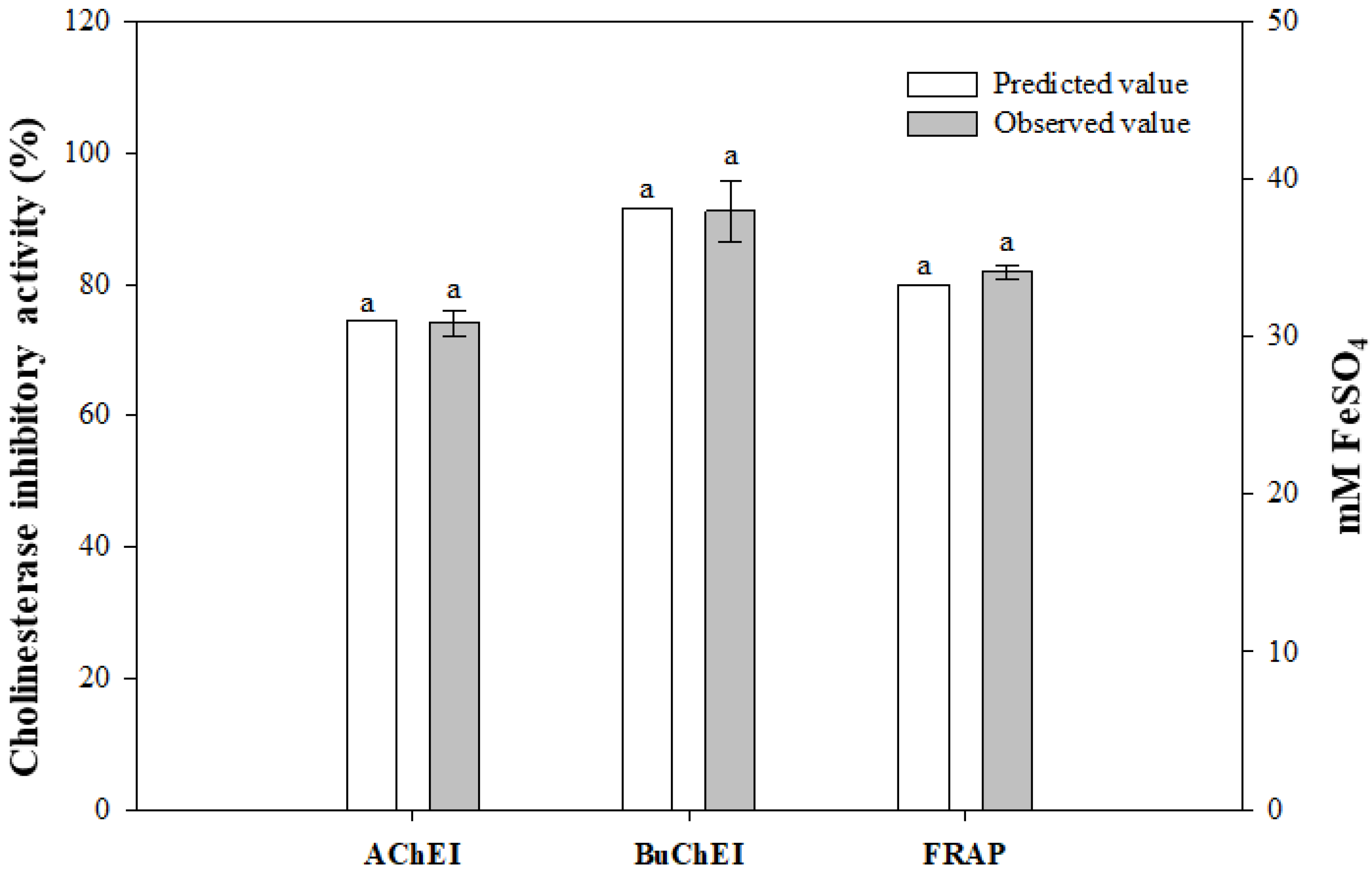
3.2.3. Model Verification
3.3. Optimization of A. capillaris Fermented Product
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2016 Neurology Collaborators. Global, regional, and national burden of neurological disorders, 1996-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef]
- Santana-Galvez, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. A practical guide for designing effective nutraceutical combinations in the form of foods, beverages, and dietary supplements against chronic degenerative diseases. Trends Food Sci. Technol. 2019, 88, 179–193. [Google Scholar] [CrossRef]
- Bartus, R.T.; Dean, R.L.; Beer, B.; Lippa, A.S. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Tõugu, V. Acetylcholinesterase: Mechanism of catalysis and inhibition. Curr. Med. Chem. 2001, 1, 155–170. [Google Scholar] [CrossRef]
- Masson, P.; Bec, N.; Froment, M.T.; Nachon, F.; Balny, C.; Lockridge, O.; Schopfer, L.M. Rate-determining step of butyrylcholinesterase-catalyzed hydrolysis of benzoylcholine and benzoylthiocholine. Eur. J. Biochem 2004, 271, 1980–1990. [Google Scholar] [CrossRef]
- McGleenon, B.M.; Dynan, K.B.; Passmore, A.P. Acetylcholinesterase inhibitors in Alzheimer’s disease. Br. J. Clin. Pharmacol. 1999, 48, 471–480. [Google Scholar] [CrossRef]
- Mukhergee, P.K.; Kumar, V.K.; Mal, M.N.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2020, 190, 108352. [Google Scholar] [CrossRef]
- Choi, B.B.; Lee, H.J.; Bang, S.K. Studies on the amino acid, sugar analysis and antioxidative effect of extracts from Artemisia sp. Korean J. Food Nutr. 2004, 17, 86–91. [Google Scholar]
- Park, J.C.; Yu, Y.B.; Lee, J.H.; Kim, N.J. Studies on the chemical components and biological activities of edible plants in Korea. Korean J. Soc. Food Nutr. 1994, 23, 116–119. [Google Scholar]
- Kim, K.C.; Kim, J.S. Effect of varying ethanol concentrations on the extraction properties and physiological activity of Artemisia annua L. Korean J. Food Sci. Technol. 2020, 52, 130–137. [Google Scholar]
- Guo, S.; Ma, J.; Xing, Y.; Xu, Y.; Jin, X.; Yan, S.; Shi, B. Artemisia annua L. aqueous extract as an alternative to antibiotics improving growth performance and antioxidant function in broilers. Ital. J. Anim. Sci. 2020, 19, 399–409. [Google Scholar] [CrossRef]
- Protti, M.; Mandriolo, R.; Mandrone, M.; Cappadone, C.; Farruggia, G.; Chiocchio, I.; Malucelli, E.; Poli, F.; Mercolini, L. Analysis of Artemisia annua extracts and related products by high performance liquid chromatography-tandem mass spectrometry coupled to sample treatment miniaturization. J. Pharm. Biomed. Anal. 2019, 174, 81–88. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, K.L. Anti-oxidative and anti-inflammatory effects of Artemisia capillaris extract. Korean J. Aesthet. Cosmetol. 2015, 13, 805–812. [Google Scholar]
- Shoaib, M.; Shah, I.; Ali, N.; Shah, S.W.A. In vitro acetylcholinesterase and butyrylcholinesterase inhibitory potentials of essential oil of Artemisia macrocephala. Bangladesh J. Pharmaacol. 2015, 10, 87–91. [Google Scholar] [CrossRef]
- Choi, J.S.; Song, B.M.; Park, H.J. Identification of the component with anti-acetylcholinesterase activity from the essential oil of Artemisia iwayomogi. J. Plant Res. 2017, 30, 17–21. [Google Scholar]
- Saarela, M.; Mogensen, G.; Fonden, R.; Matto, J.; Sandholm, T.M. Probiotic bacteria: Safety, functional and technological properties. J. Biotechnol. 2000, 84, 197–215. [Google Scholar] [CrossRef]
- Mayer, E.A.; Knight, R.; Mazmanian, S.K.; Cryan, J.F.; Tillisch, K. Gut microbes and the brain: Paradigm shift in neuroscience. J. Neurosci. 2014, 34, 15490–15496. [Google Scholar] [CrossRef]
- Lee, H.Y.; Lee, H.S. The roles of dietary polyphenols in brain neuromodulation. J. Life Sci. 2018, 28, 1386–1395. [Google Scholar]
- Choi, J.H.; Jin, Y.H.; Kim, J.H.; Hong, Y.G. Does the gut microbiota regulate a cognitive function? J. Life Sci. 2019, 29, 747–753. [Google Scholar]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health benefits of lactic acid bacteria (LAB) fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Zhao, M.; Tang, J.; Deng, L.; Feng, F. Probiotics-fermented blueberry juices as potential antidiabetic product: Antioxidant, antimicrobial and antidiabetic potentials. J. Sci. Food Agric. 2021, 101, 4420–4427. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yoon, J.; Seong, H.; Jeong, Y. Novel Leuconostoc Mesenteroides MKJW and Its Uses Thereof. KR Patent 102021001464, 2 February 2021. [Google Scholar]
- Sharififar, F.; Moshafi, M.H.; Shafazand, E.; Koohpayeh, A. Acetylcholinesterase inhibitory, antioxidant and cytotoxic activity of three dietary medicinal plants. Food Chem. 2012, 130, 20–23. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, S.K.; Park, S.I.; Kim, I.Y.; Jeong, Y.H.; Yu, S.Y.; Shin, S.C.; Kim, M.S. Antioxidant activity of Korea traditional soy sauce fermented in Korean earthenware, onggi, from different regions. J. Korean Soc. Food Sci. Nutr. 2015, 44, 847–853. [Google Scholar] [CrossRef]
- Lin, M.Y.; Yen, C.L. Antioxidative ability of lactic acid bacteria. J. Agric. Food Chem. 1999, 47, 1460–1466. [Google Scholar] [CrossRef]
- Cermeno, M.; FitzGerald, R.J.; O’Brien, N.M. In vitro antioxidant, and immunomodulatory activity of transglutaminase-treated sodium caseinate hydrolysates. Int. Dairy J. 2016, 63, 107–114. [Google Scholar] [CrossRef]
- Anesini, C.; Ferraro, G.E.; Filip, R. Total polyphenol content and antioxidant capacity of commercially available tea (Camellia sinensis) in Argentina. J. Agric. Food Chem. 2008, 56, 9225–9229. [Google Scholar] [CrossRef]
- Isshiki, K.; Nishinomiya, T.; Nozaka, N.; Tokuoka, K. Growth inhibition of microorganisms by plant extracts. Nippon Shokuhin Kogyo Gakkaishi 1993, 40, 525–527. [Google Scholar] [CrossRef]
- Sagdic, O.; Karahan, A.G.; Ozcan, M.; Ozkan, G. Note: Effect of some spice extracts on bacterial inhibition. Food Sci. Technol. Int. 2003, 9, 353–356. [Google Scholar] [CrossRef]
- Naili, M.B.; Alghazeer, R.O.; Saleh, N.A.; Al-Najjar, A.Y. Evaluation of antibacterial and antioxidant activities of Artemisia campestris (Astraceae) and ZiZiphus lotus (Rhamnacea). Arab. J. Chem. 2010, 3, 79–84. [Google Scholar] [CrossRef]
- Mannan, H.A.; Ahmed, I.; Hussain, I.; Jamil, M.; Miza, B. Antibacterial activity and brine shrimp toxicity of Artemisia dubia extract. Pak. J. Bot. 2012, 44, 1487–1490. [Google Scholar]
- Ghlissi, Z.; Sayari, N.; Kallel, R.; Bougatef, A.; Sahnoun, Z. Antioxidant, antibacterial, anti-inflammatory, and wound healing effects of Artemisia campestris aqueous extract in rat. Biomed. Pharmacother. 2016, 84, 115–122. [Google Scholar] [CrossRef]
- Kyoung, S.S.; Hyung, J.J.; Kyeong, W.Y. Antimicrobial activity and chemical components of two plants. Artemisia capillaris and Artemisia iwayomogi, used as Korean herbal Injin. J. Ecol. Field Biol. 2010, 33, 141–147. [Google Scholar]
- Park, I.B.; Park, J.W.; Lee, Y.J.; Shin, G.W.; Kim, H.S.; Jo, Y.C. Quality characteristic of glasswort (Salicornia herbacea L.) fermented by Bacillus subtilis. Korean J. Soc. Food Sci. Nutr. 2009, 38, 902–908. [Google Scholar] [CrossRef]
- Yang, S.J.; Kim, Y.C.; Hong, J.H. Cultural characteristics and biological activities of cereals fermented by Bacillus subtilis CBD2. J. Chitin Chitosan 2014, 19, 277–284. [Google Scholar]
- Rheem, S.; Rheem, I.; Oh, S. Response surface methodology using a fullest balanced model: A re-analysis of a dataset in the Korean journal for food science of animal resources. Food Sci. Anim. Resour. 2017, 37, 139–146. [Google Scholar] [CrossRef]
- Lee, A.R.; Niu, K.M.; Kang, S.K.; Han, S.G.; Lee, J.B.; Kim, S.K. Antioxidant, and antibacterial activities of Lactobacillus-fermented Artemisia annua L. as a potential fish feed additive. J. Life Sci. 2017, 27, 652–660. [Google Scholar]
- Lee, D.S.; Lee, I.H. Development of Monacolin K-enriched Ganghwayakssuk (Artemisia princeps Pamp.) by fermentation with Monascus pilosus. J. Microbiol. Biotechnol. 2012, 22, 975–980. [Google Scholar] [CrossRef]
- Mekinic, I.G.; Burcul, F.; Blazevic, I.B.; Skroza, D.; Kerum, D.; Katalinic, V. Antioxidative/Acetylcholinesterase inhibitory activity of some Asteraceae Plants. Nat. Prod. Commun. 2013, 8, 471–474. [Google Scholar] [CrossRef]
- Mesquita, R.S.; Kyrylchuk, A.; Oliveira, R.C.; Sa’, I.S.C.; Camargo, G.C.B.; Pontes, G.S.; Silva, F.M.A.; Nunomura, R.C.S.; Grafov, A. Alkaloids of Abuta panurensis Eichler: In silico and in vitro study of acetylcholinesterase inhibition, cytotoxic and immunomodulatory activities. PLoS ONE 2020, 15, e0239364. [Google Scholar]
- Christen, Y. Oxidative stress and Alzheimer disease. Am. J. Clin. Nutr. 2000, 71, 621S–629S. [Google Scholar] [CrossRef]
- Papandreou, M.A.; Dimakopoulou, A.; Linardaki, Z.I.; Cordopatis, P.; Zacas, D.K.; Margarity, M.; Lamari, F.N. Effect of a polyphenol-rich wild blueberry extract on cognitive performance of mice, brain antioxidant markers and acetylcholinesterase activity. Behav. Brain Res. 2009, 198, 352–358. [Google Scholar] [CrossRef]
- Kim, Y.M.; Jeong, H.J.; Chung, H.S.; Seong, J.H.; Kim, H.S.; Kim, D.S.; Lee, Y.G. Anti-oxidative activity of the extracts from Houttuynia cordata Thunb. Fermented by lactic acid bacteria. J. Life Sci. 2016, 26, 468–474. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Levels | ||
|---|---|---|---|
| Low | Mid | High | |
| X1: A. capillaris (%) | 3 | 5 | 7 |
| X2: Gryllus bimaculatus (%) | 0 | 1 | 2 |
| X3: Yeast extract (%) | 0 | 1 | 2 |
| Run | A. capillaris (%, X1) | G. bimaculatus (%, X2) | Yeast Extract (%, X3) | Log CFU/mL 1 | pH | AChE Inhibitory Activity (%, Y1) | BuChE Inhibitory Activity (%, Y2) | FRAP (mM FeSO4, Y3) | Reducing Power (%, Y4) | DPPH 2 (%, Y5) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 12 h | 0 h | 12 h | |||||||||
| 1 | 5 | 1 | 1 | 6.85 | 8.34 | 5.36 | 4.84 | 73.8 | 50.0 | 24.7 | 34.1 | 43.1 |
| 2 | 5 | 1 | 1 | 6.85 | 8.41 | 5.36 | 4.86 | 78.0 | 52.6 | 21.5 | 31.5 | 42.2 |
| 3 | 7 | 2 | 1 | 6.85 | 8.28 | 5.28 | 5.08 | 69.9 | 64.4 | 28.5 | 39.3 | 53.3 |
| 4 | 5 | 2 | 0 | 6.85 | 8.48 | 5.45 | 4.76 | 56.2 | 50.6 | 20.6 | 24.3 | 30.9 |
| 5 | 5 | 1 | 1 | 6.85 | 8.39 | 5.36 | 4.79 | 74.3 | 48.3 | 24.6 | 31.4 | 39.7 |
| 6 | 5 | 1 | 1 | 6.85 | 8.43 | 5.36 | 4.73 | 70.9 | 48.9 | 23.5 | 31.5 | 39.4 |
| 7 | 3 | 1 | 0 | 6.85 | 8.68 | 5.15 | 4.15 | 40.9 | 36.4 | 12.6 | 13.8 | 18.5 |
| 8 | 7 | 0 | 1 | 6.85 | 7.57 | 5.19 | 5.02 | 91.9 | 82.4 | 33.2 | 42.2 | 62.8 |
| 9 | 5 | 0 | 2 | 6.85 | 7.97 | 5.32 | 5.03 | 79.9 | 60.2 | 30.7 | 44.6 | 60.5 |
| 10 | 7 | 1 | 0 | 6.85 | 7.73 | 5.24 | 5.12 | 75.9 | 80.5 | 29.4 | 35.1 | 52.4 |
| 11 | 3 | 0 | 1 | 6.85 | 8.51 | 5.11 | 4.36 | 56.9 | 36.8 | 17.5 | 24.6 | 27.6 |
| 12 | 5 | 2 | 2 | 6.85 | 8.61 | 5.37 | 4.77 | 72.7 | 60.6 | 27.1 | 40.8 | 45.9 |
| 13 | 3 | 2 | 1 | 6.85 | 8.99 | 5.44 | 4.22 | 51.3 | 44.9 | 16.8 | 22.8 | 23.4 |
| 14 | 7 | 1 | 2 | 6.85 | 8.06 | 5.34 | 5.08 | 87.6 | 67.0 | 36.5 | 51.9 | 66.7 |
| 15 | 5 | 0 | 0 | 6.85 | 7.53 | 5.11 | 4.99 | 81.1 | 64.8 | 25.0 | 30.0 | 43.7 |
| 16 | 5 | 1 | 1 | 6.85 | 8.38 | 5.36 | 4.81 | 76.0 | 50.7 | 23.4 | 30.7 | 39.2 |
| 17 | 3 | 1 | 2 | 6.85 | 8.90 | 5.41 | 4.36 | 55.2 | 47.2 | 18.0 | 24.5 | 26.6 |
| AChE Inhibitory Activity (Y1) | BuChE Inhibitory Activity (Y2) | FRAP (Y3) | ||||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| Model 1 | 32.83 | <0.0001 | 99.08 | <0.0001 | 77.12 | <0.0001 |
| X1 | 186.78 | <0.0001 | 674.66 | <0.0001 | 192.66 | <0.0001 |
| X2 | 45.47 | 0.0003 | 22.77 | 0.0020 | 8.80 | 0.0109 |
| X3 | 21.76 | 0.0023 | 0.2956 | 0.6036 | 29.90 | 0.0001 |
| X1X2 | 6.86 | 0.0344 | 55.24 | 0.0001 | ||
| X1X3 | 0.1725 | 0.6904 | 47.88 | 0.0002 | ||
| X2X3 | 7.99 | 0.0255 | 17.28 | 0.0043 | ||
| X12 | 23.14 | 0.0019 | 11.29 | 0.0121 | ||
| X22 | 0.0242 | 0.8807 | 23.52 | 0.0019 | ||
| X32 | 2.40 | 0.1654 | 31.46 | 0.0008 | ||
| Lack of fit | 1.94 | 0.2650 | 1.21 | 0.4129 | 1.77 | 0.3053 |
| Run | AChE Inhibitory Activity (%) | BuChE Inhibitory Activity (%) | FRAP (mM FeSO4) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Observed Y | Predicted Y | Residuals | Observed Y | Predicted Y | Residuals | Observed Y | Predicted Y | Residuals | |
| 1 | 73.80 | 74.60 | –0.80 | 50.00 | 50.10 | –0.10 | 24.70 | 24.33 | 0.37 |
| 2 | 78.00 | 74.60 | 3.40 | 52.60 | 50.10 | 2.50 | 21.50 | 24.33 | –2.83 |
| 3 | 69.90 | 71.06 | –1.16 | 64.40 | 63.76 | 0.64 | 28.50 | 30.49 | –1.99 |
| 4 | 56.20 | 55.43 | 0.78 | 50.60 | 52.10 | –1.50 | 20.60 | 19.57 | 1.03 |
| 5 | 74.30 | 74.60 | –0.30 | 48.30 | 50.10 | –1.80 | 24.60 | 24.33 | 0.27 |
| 6 | 70.90 | 74.60 | –3.70 | 48.90 | 50.10 | –1.20 | 23.50 | 24.33 | –0.83 |
| 7 | 40.90 | 43.96 | –3.06 | 36.40 | 35.24 | 1.16 | 12.60 | 13.40 | –0.80 |
| 8 | 91.90 | 94.19 | –2.29 | 82.40 | 82.74 | –0.34 | 33.20 | 33.84 | –0.64 |
| 9 | 79.90 | 80.68 | –0.77 | 60.20 | 58.70 | 1.50 | 30.70 | 29.09 | 1.61 |
| 10 | 75.90 | 75.51 | 0.39 | 80.50 | 79.64 | 0.86 | 29.40 | 29.08 | 0.32 |
| 11 | 56.90 | 55.74 | 1.16 | 36.80 | 37.44 | –0.64 | 17.50 | 18.17 | –0.67 |
| 12 | 72.70 | 74.60 | –1.90 | 60.60 | 60.08 | 0.52 | 27.10 | 25.74 | 1.36 |
| 13 | 51.30 | 49.01 | 2.29 | 44.90 | 44.56 | 0.34 | 16.80 | 14.82 | 1.98 |
| 14 | 87.60 | 84.54 | 3.06 | 67.00 | 68.16 | –1.16 | 36.50 | 35.25 | 1.25 |
| 15 | 81.10 | 79.20 | 1.90 | 64.80 | 65.33 | –0.53 | 25.00 | 22.92 | 2.08 |
| 16 | 76.00 | 74.60 | 1.40 | 50.70 | 50.10 | 0.60 | 23.40 | 24.33 | –0.93 |
| 17 | 55.20 | 55.59 | –0.39 | 47.20 | 48.06 | –0.86 | 18.00 | 19.58 | –1.58 |
| DPPH Radical Scavenging Activity (%) | Reducing Power (Abs at 700 nm) | FRAP (mM FeSO4) | TPC (mg GAE/mL) | |
|---|---|---|---|---|
| Optimal conditions 1 | 86.5 ± 0.89 | 2.14 ± 0.02 | 34.1 ± 0.43 | 1.52 ± 0.61 |
| Ascorbic acid 2 | 97.3 ± 0.65 | 2.68 ± 0.15 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Yoon, J.; Kim, M. Optimization of Fermentation Conditions of Artemisia capillaris for Enhanced Acetylcholinesterase and Butyrylcholinesterase. Foods 2022, 11, 2268. https://doi.org/10.3390/foods11152268
Choi J, Yoon J, Kim M. Optimization of Fermentation Conditions of Artemisia capillaris for Enhanced Acetylcholinesterase and Butyrylcholinesterase. Foods. 2022; 11(15):2268. https://doi.org/10.3390/foods11152268
Chicago/Turabian StyleChoi, Jina, Jiwon Yoon, and Misook Kim. 2022. "Optimization of Fermentation Conditions of Artemisia capillaris for Enhanced Acetylcholinesterase and Butyrylcholinesterase" Foods 11, no. 15: 2268. https://doi.org/10.3390/foods11152268
APA StyleChoi, J., Yoon, J., & Kim, M. (2022). Optimization of Fermentation Conditions of Artemisia capillaris for Enhanced Acetylcholinesterase and Butyrylcholinesterase. Foods, 11(15), 2268. https://doi.org/10.3390/foods11152268