
Effects of a Mixed Limosilactobacillus fermentum Formulation with Claimed Probiotic Properties on Cardiometabolic Variables, Biomarkers of Inflammation and Oxidative Stress in Male Rats Fed a High-Fat Diet

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Animals and Ethical Aspects
2.2. Probiotic Strains and Preparation of Probiotic Suspension
2.3. Experimental Design
2.4. Quantification of Organic Acids in Colonic Contents
2.5. Biochemical Analysis and Atherogenic Indices
2.6. Measurement of Oxidative Stress in Colon and Heart
2.7. Statistical Analysis
3. Results
3.1. Body Weight, Biochemical Parameters and Cytokine Serum Levels
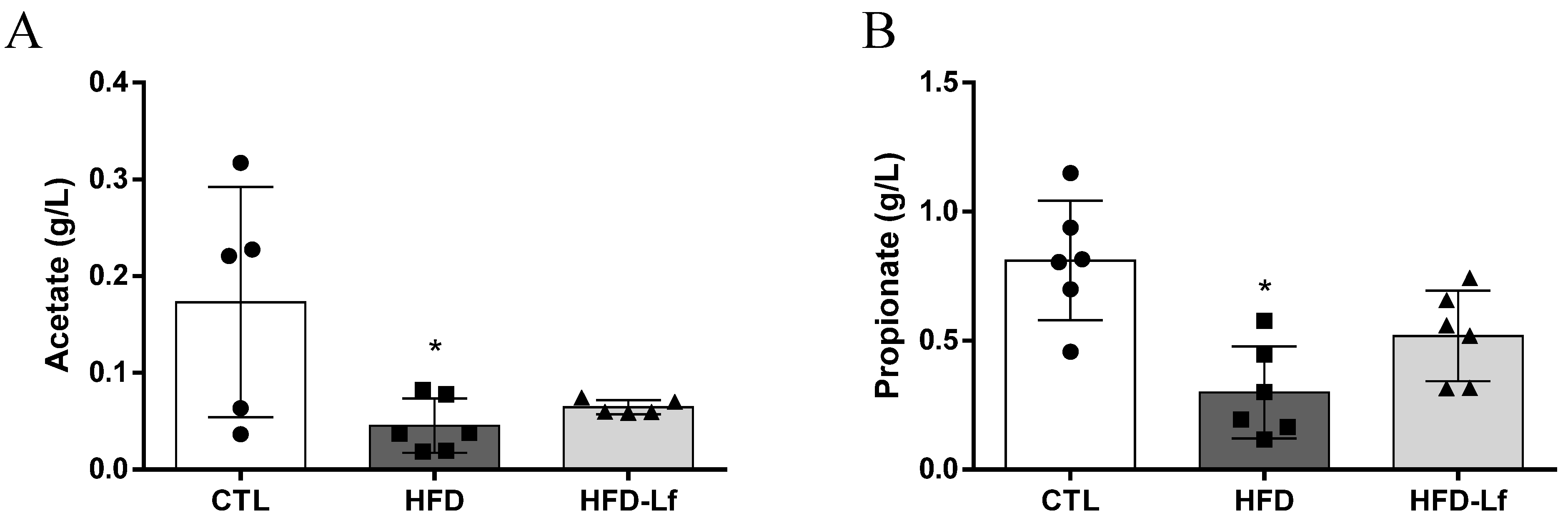
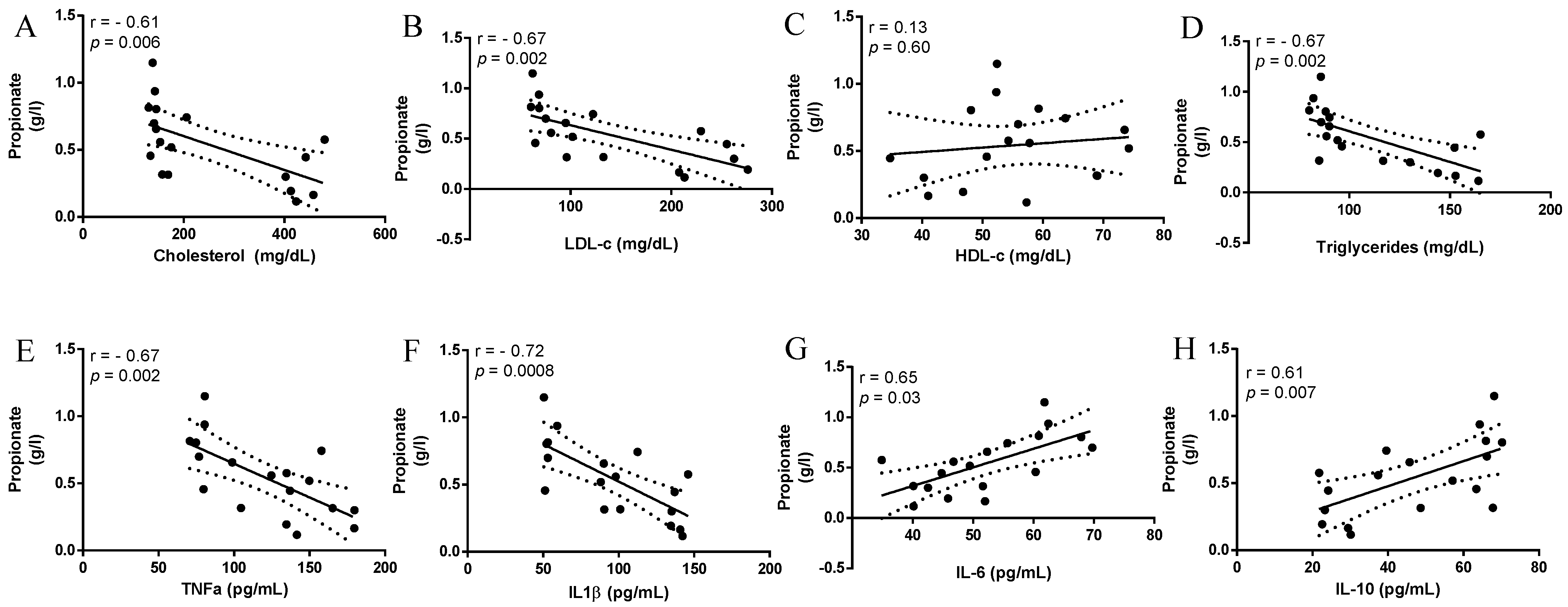
3.2. Short-Chain Fatty Acids, Fructose and Raffinose in Feces
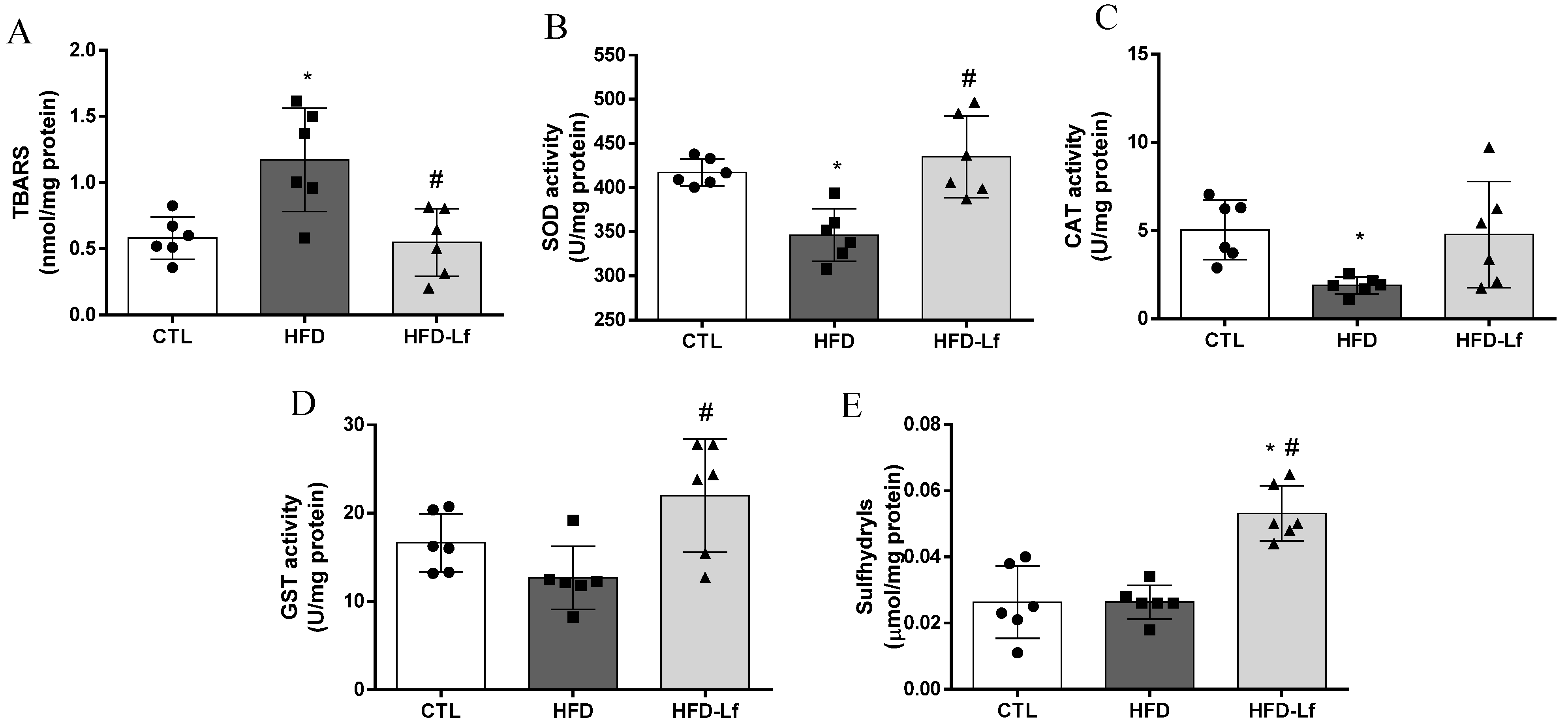
3.3. Oxidative Stress Biomarkers in Colon Tissues
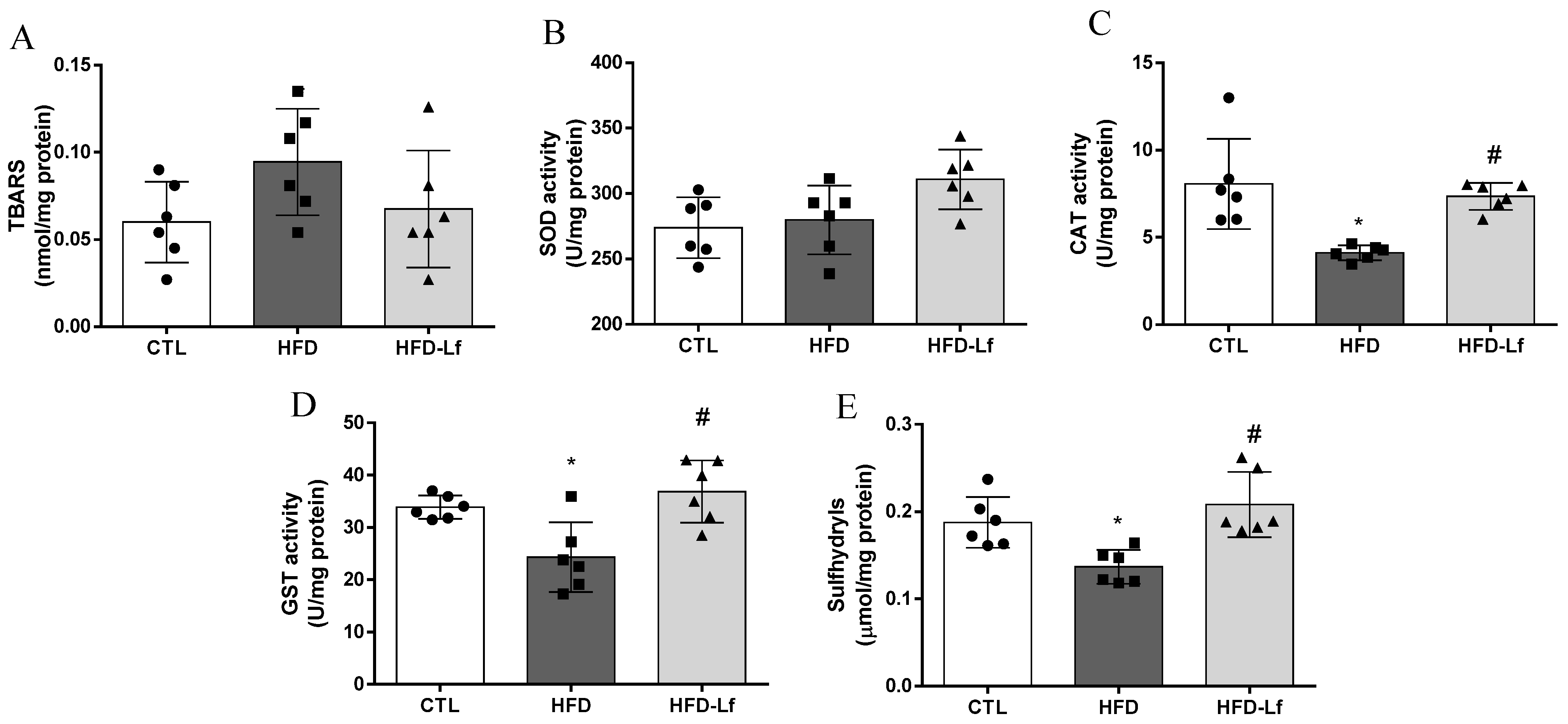
3.4. Oxidative Stress Biomarkers in Heart Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madan, S.; Mehra, M.R. The heart-gut microbiome axis in advanced heart failure. J. Heart Lung Transpl. 2020, 39, 891–893. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Liong, M.T.; Tsai, Y.C. New perspectives of Lactobacillus plantarum as a probiotic: The gut-heart-brain axis. J. Microbiol. 2018, 56, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef]
- Schulz, M.D.; Atay, C.; Heringer, J.; Romrig, F.K.; Schwitalla, S.; Aydin, B.; Ziegler, P.K.; Varga, J.; Reindl, W.; Pommerenke, C.; et al. High-fat-diet-mediated dysbiosis promotes intestinal carcinogenesis independently of obesity. Nature 2014, 514, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, X.; Sun, Y.; Du, J.; Li, X.; Xun, Z.; Li, Y.C. High-fat diet promotes experimental colitis by inducing oxidative stress in the colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G453–G462. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Apaijai, N.; Arinno, A.; Palee, S.; Pratchayasakul, W.; Kerdphoo, S.; Jaiwongkam, T.; Chunchai, T.; Chattipakorn, S.C.; Chattipakorn, N. High-Saturated Fat High-Sugar Diet Accelerates Left-Ventricular Dysfunction Faster than High-Saturated Fat Diet Alone via Increasing Oxidative Stress and Apoptosis in Obese-Insulin Resistant Rats. Mol. Nutr. Food Res. 2019, 63, e1800729. [Google Scholar] [CrossRef]
- Cavalcante, R.G.S.; de Albuquerque, T.M.R.; de Luna Freire, M.O.; Ferreira, G.A.H.; Carneiro Dos Santos, L.A.; Magnani, M.; Cruz, J.C.; Braga, V.A.; de Souza, E.L.; de Brito Alves, J.L. The probiotic Lactobacillus fermentum 296 attenuates cardiometabolic disorders in high fat diet-treated rats. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1408–1417. [Google Scholar] [CrossRef]
- Nagatomo, Y.; Tang, W.H. Intersections Between Microbiome and Heart Failure: Revisiting the Gut Hypothesis. J. Card Fail. 2015, 21, 973–980. [Google Scholar] [CrossRef]
- Cavalcanti Neto, M.P.; Aquino, J.S.; Romao da Silva, L.F.; de Oliveira Silva, R.; Guimaraes, K.S.L.; de Oliveira, Y.; de Souza, E.L.; Magnani, M.; Vidal, H.; de Brito Alves, J.L. Gut microbiota and probiotics intervention: A potential therapeutic target for management of cardiometabolic disorders and chronic kidney disease? Pharmacol. Res. 2018, 130, 152–163. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O'Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Lomax, A.R.; Calder, P.C. Probiotics, immune function, infection and inflammation: A review of the evidence from studies conducted in humans. Curr. Pharm. Des. 2009, 15, 1428–1518. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.F.; Luciano, W.A.; Xavier, D.E.; da Costa, W.C.; de Sousa Oliveira, K.; Franco, O.L.; de Morais Junior, M.A.; Lucena, B.T.; Picao, R.C.; Magnani, M.; et al. Identification of Lactic Acid Bacteria in Fruit Pulp Processing Byproducts and Potential Probiotic Properties of Selected Lactobacillus Strains. Front. Microbiol. 2016, 7, 1371. [Google Scholar] [CrossRef]
- de Albuquerque, T.M.R.; Garcia, E.F.; de Oliveira Araujo, A.; Magnani, M.; Saarela, M.; de Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Proteins 2018, 10, 704–716. [Google Scholar] [CrossRef]
- de Oliveira, Y.; Cavalcante, R.G.S.; Cavalcanti Neto, M.P.; Magnani, M.; Braga, V.A.; de Souza, E.L.; de Brito Alves, J.L. Oral administration of Lactobacillus fermentum post-weaning improves the lipid profile and autonomic dysfunction in rat offspring exposed to maternal dyslipidemia. Food Funct. 2020, 11, 5581–5594. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Bezerra, M.L.R.; de Souza, E.L.; de Sousa, J.M.B.; Lima, M.D.S.; Alves, A.F.; Almeida, M.D.G.; Coutinho Alves, R.; Verissimo de Araujo, E.; Soares, N.L.; da Silva, G.A.; et al. Effects of honey from Mimosa quadrivalvis L. (malicia) produced by the Melipona subnitida D. (jandaira) stingless bee on dyslipidaemic rats. Food Funct. 2018, 9, 4480–4492. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Martirosyan, D.M.; Miroshnichenko, L.A.; Kulakova, S.N.; Pogojeva, A.V.; Zoloedov, V.I. Amaranth oil application for coronary heart disease and hypertension. Lipids Health Dis. 2007, 6, 1. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dobiasova, M. Atherogenic index of plasma [log(triglycerides/HDL-cholesterol)]: Theoretical and practical implications. Clin. Chem. 2004, 50, 1113–1115. [Google Scholar] [CrossRef] [PubMed]
- Castelli, W.P.; Abbott, R.D.; McNamara, P.M. Summary estimates of cholesterol used to predict coronary heart disease. Circulation 1983, 67, 730–734. [Google Scholar] [CrossRef]
- Pedroza, A.; Ferreira, D.S.; Santana, D.F.; da Silva, P.T.; de Aguiar Junior, F.C.A.; Sellitti, D.F.; Lagranha, C.J. A maternal low-protein diet and neonatal overnutrition result in similar changes to glomerular morphology and renal cortical oxidative stress measures in male Wistar rats. Appl. Physiol. Nutr. Metab. 2019, 44, 164–171. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Palaniyandi, S.A.; Damodharan, K.; Suh, J.W.; Yang, S.H. Probiotic Characterization of Cholesterol-Lowering Lactobacillus fermentum MJM60397. Probiotics Antimicrob. Proteins 2020, 12, 1161–1172. [Google Scholar] [CrossRef]
- Yadav, R.; Khan, S.H.; Mada, S.B.; Meena, S.; Kapila, R.; Kapila, S. Consumption of Probiotic Lactobacillus fermentum MTCC: 5898-Fermented Milk Attenuates Dyslipidemia, Oxidative Stress, and Inflammation in Male Rats Fed on Cholesterol-Enriched Diet. Probiotics Antimicrob. Proteins 2019, 11, 509–518. [Google Scholar] [CrossRef]
- Kullisaar, T.; Zilmer, K.; Salum, T.; Rehema, A.; Zilmer, M. The use of probiotic L. fermentum ME-3 containing Reg'Activ Cholesterol supplement for 4 weeks has a positive influence on blood lipoprotein profiles and inflammatory cytokines: An open-label preliminary study. Nutr. J. 2016, 15, 93. [Google Scholar] [CrossRef]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martin-Gari, M.; Sanchez, V.; Riart Solans, M.; Berdun, R.; Ludwig, I.A.; Rubio, L.; Vilaprinyo, E.; Portero-Otin, M.; Serrano, J.C.E. Faecal bacterial and short-chain fatty acids signature in hypercholesterolemia. Sci. Rep. 2019, 9, 1772. [Google Scholar] [CrossRef] [PubMed]
- Weitkunat, K.; Schumann, S.; Nickel, D.; Kappo, K.A.; Petzke, K.J.; Kipp, A.P.; Blaut, M.; Klaus, S. Importance of propionate for the repression of hepatic lipogenesis and improvement of insulin sensitivity in high-fat diet-induced obesity. Mol. Nutr. Food Res. 2016, 60, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. Int. J. Mol. Sci. 2020, 21, 3026. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Cani, P.D. Is colonic propionate delivery a novel solution to improve metabolism and inflammation in overweight or obese subjects? Gut 2019, 68, 1352–1353. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Ding, Y.; Le, G.; Shi, Y. Alterations of the gut microbiota in high-fat diet mice is strongly linked to oxidative stress. Appl. Microbiol. Biotechnol. 2013, 97, 1689–1697. [Google Scholar] [CrossRef]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.M.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef]
- Pussinen, P.J.; Havulinna, A.S.; Lehto, M.; Sundvall, J.; Salomaa, V. Endotoxemia is associated with an increased risk of incident diabetes. Diabetes Care 2011, 34, 392–397. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Tan, F.; Zhou, X.; Mu, J.; Zhao, X. Lactobacillus fermentum CQPC07 attenuates obesity, inflammation and dyslipidemia by modulating the antioxidant capacity and lipid metabolism in high-fat diet induced obese mice. J. Inflamm. Lond. 2021, 18, 5. [Google Scholar] [CrossRef]
- Mikelsaar, M.; Zilmer, M. Lactobacillus fermentum ME-3—An antimicrobial and antioxidative probiotic. Microb. Ecol. Health Dis. 2009, 21, 1–27. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; de la Visitacion, N.; Sanchez, M.; Romero, M.; Olivares, M.; Jimenez, R.; Duarte, J. The Probiotic Lactobacillus fermentum Prevents Dysbiosis and Vascular Oxidative Stress in Rats with Hypertension Induced by Chronic Nitric Oxide Blockade. Mol. Nutr. Food Res. 2018, 62, e1800298. [Google Scholar] [CrossRef]
- Wang, A.N.; Cai, C.J.; Zeng, X.F.; Zhang, F.R.; Zhang, G.L.; Thacker, P.A.; Wang, J.J.; Qiao, S.Y. Dietary supplementation with Lactobacillus fermentum I5007 improves the anti-oxidative activity of weanling piglets challenged with diquat. J. Appl. Microbiol. 2013, 114, 1582–1591. [Google Scholar] [CrossRef]
- Jang, Y.J.; Kim, W.K.; Han, D.H.; Lee, K.; Ko, G. Lactobacillus fermentum species ameliorate dextran sulfate sodium-induced colitis by regulating the immune response and altering gut microbiota. Gut Microbes 2019, 10, 696–711. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In vitro immunomodulatory activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two probiotic strains isolated from human breast milk. Immunobiology 2010, 215, 996–1004. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Singh, B.P.; Sandhu, N.; Singh, N.; Kaur, R.; Rokana, N.; Singh, K.S.; Chaudhary, V.; Panwar, H. Probiotic mediated NF-kappaB regulation for prospective management of type 2 diabetes. Mol. Biol. Rep. 2020, 47, 2301–2313. [Google Scholar] [CrossRef]
- Campese, V.M.; Shaohua, Y.; Huiquin, Z. Oxidative stress mediates angiotensin II-dependent stimulation of sympathetic nerve activity. Hypertension 2005, 46, 533–539. [Google Scholar] [CrossRef]
- Levick, S.P.; Murray, D.B.; Janicki, J.S.; Brower, G.L. Sympathetic nervous system modulation of inflammation and remodeling in the hypertensive heart. Hypertension 2010, 55, 270–276. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut-heart axis in atherosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Ke, B.; Du, J. TMAO: How gut microbiota contributes to heart failure. Transl. Res. 2021, 228, 109–125. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/100 g) | Diets | |
|---|---|---|
| Control (AIN-93M) * | HFD ** | |
| Corn starch | 39.75 | 33.09 |
| Dextrinized corn starch | 13.20 | 15.50 |
| Casein # | 20.00 | 19.86 |
| Sucrose | 10.00 | 6.00 |
| Soybean oil | 7.00 | 3.00 |
| Animal fat (lard) | 0.00- | 6.00 |
| Non-hydrolyzed vegetable fat | 0.00 | 5.00 |
| Sigma cholesterol | 0.00 | 1.00 |
| Sigma colic acid | 0.00 | 0.50 |
| Cellulose | 5.00 | 5.00 |
| Mineral mix 93M | 3.50 | 3.50 |
| Vitamin mix | 1.00 | 1.00 |
| L-cystine | 0.30 | 0.30 |
| Choline bitartrate | 0.25 | 0.25 |
| t-BHQ *** | 0.014 | 0.014 |
| Nutritional composition | ||
| Calories (Kj/100 g) | 16.46 | 18.05 |
| Carbohydrate (%) | 63.8 | 50.5 |
| Protein (%) | 20.3 | 18.3 |
| Lipids (%) | 15.9 | 31.2 |
| CTL (n = 6) | HFD (n = 6) | HFD-Lf (n = 6) | F | p-Value | |
|---|---|---|---|---|---|
| % Weight gain | 12.1 ± 2.9 | 6.6 ± 3.6 | 7.6 ± 4.4 | 2.9 | 0.09 |
| Biochemical parameters | |||||
| Glucose (mmol/L) | 5.8 ± 0.5 | 11.2 ± 2.0 * | 8.2 ± 0.9 * # | 25.45 | <0.0001 |
| Triglycerides (mmol/L) | 0.9 ± 0.03 | 1.6 ± 0.12 * | 1.1 ± 0.06 * # | 69.05 | <0.0001 |
| Total cholesterol (mmol/L) | 36.1 ± 1.4 | 113.5 ± 7.6 * | 43.6 ± 5.6 * # | 360.3 | <0.0001 |
| LDL-cholesterol (mmol/L) | 17.4 ± 1.4 | 62.5 ± 7.3 * | 27.3 ± 5.0 * # | 128.4 | <0.0001 |
| HDL-cholesterol (mmol/L) | 14.5 ± 1.0 | 10.6 ± 2.7 * | 17.5 ± 1.6 * # | 17.47 | 0.0001 |
| Atherogenic indices | |||||
| CRR | 2.5 ± 0.2 | 11.3 ± 2.9 * | 2.5 ± 0.5 # | 53.11 | <0.0001 |
| AIP | 1.5 ± 0.1 | 3.5 ± 0.7 * | 1.4 ± 0.2 # | 54.86 | <0.0001 |
| CRI-II | 1.2 ± 0.2 | 6.2 ± 1.5 * | 1.6 ± 0.3 # | 63.93 | <0.0001 |
| Cytokine levels | |||||
| TNF-α (pg/mL) † | 78.4 (70.7–80.6) | 139.3 (134.8–179.7) * | 137.2 (98.8–158.0) * | 11.79 | 0.0003 |
| IL-6 (pg/mL) | 63.8 ± 3.9 | 43.3 ± 5.8 * | 49.3 ± 5.4 * | 25.65 | <0.0001 |
| IL-1β (pg/mL) | 52.6 ± 1.7 | 139.3 ± 4.4 * | 96.7 ± 9.2 * # | 313.0 | <0.0001 |
| IL-10 (pg/mL) | 66.4 ± 2.5 | 25.2 ± 3.6 * | 49.4 ± 11.4 * # | 51.23 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Luna Freire, M.O.; do Nascimento, L.C.P.; de Oliveira, K.Á.R.; de Oliveira, A.M.; Napoleão, T.H.; Lima, M.d.S.; Lagranha, C.J.; de Souza, E.L.; de Brito Alves, J.L. Effects of a Mixed Limosilactobacillus fermentum Formulation with Claimed Probiotic Properties on Cardiometabolic Variables, Biomarkers of Inflammation and Oxidative Stress in Male Rats Fed a High-Fat Diet. Foods 2021, 10, 2202. https://doi.org/10.3390/foods10092202
de Luna Freire MO, do Nascimento LCP, de Oliveira KÁR, de Oliveira AM, Napoleão TH, Lima MdS, Lagranha CJ, de Souza EL, de Brito Alves JL. Effects of a Mixed Limosilactobacillus fermentum Formulation with Claimed Probiotic Properties on Cardiometabolic Variables, Biomarkers of Inflammation and Oxidative Stress in Male Rats Fed a High-Fat Diet. Foods. 2021; 10(9):2202. https://doi.org/10.3390/foods10092202
Chicago/Turabian Stylede Luna Freire, Micaelle Oliveira, Luciana Caroline Paulino do Nascimento, Kataryne Árabe Rimá de Oliveira, Alisson Macário de Oliveira, Thiago Henrique Napoleão, Marcos dos Santos Lima, Cláudia Jacques Lagranha, Evandro Leite de Souza, and José Luiz de Brito Alves. 2021. "Effects of a Mixed Limosilactobacillus fermentum Formulation with Claimed Probiotic Properties on Cardiometabolic Variables, Biomarkers of Inflammation and Oxidative Stress in Male Rats Fed a High-Fat Diet" Foods 10, no. 9: 2202. https://doi.org/10.3390/foods10092202
APA Stylede Luna Freire, M. O., do Nascimento, L. C. P., de Oliveira, K. Á. R., de Oliveira, A. M., Napoleão, T. H., Lima, M. d. S., Lagranha, C. J., de Souza, E. L., & de Brito Alves, J. L. (2021). Effects of a Mixed Limosilactobacillus fermentum Formulation with Claimed Probiotic Properties on Cardiometabolic Variables, Biomarkers of Inflammation and Oxidative Stress in Male Rats Fed a High-Fat Diet. Foods, 10(9), 2202. https://doi.org/10.3390/foods10092202