
Administration of Ligilactobacillus salivarius MP101 in an Elderly Nursing Home during the COVID-19 Pandemic: Immunological and Nutritional Impact

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of the Probiotic Strain
2.2. Production and Quality Control of the Study Product
2.3. Study Design and Participants
2.4. Measurement of Functional, Cognitive and Nutritional Status of the Participants
2.5. Immunoprofiling of the Nasal and Fecal Samples
2.6. Statistical Analysis
3. Results
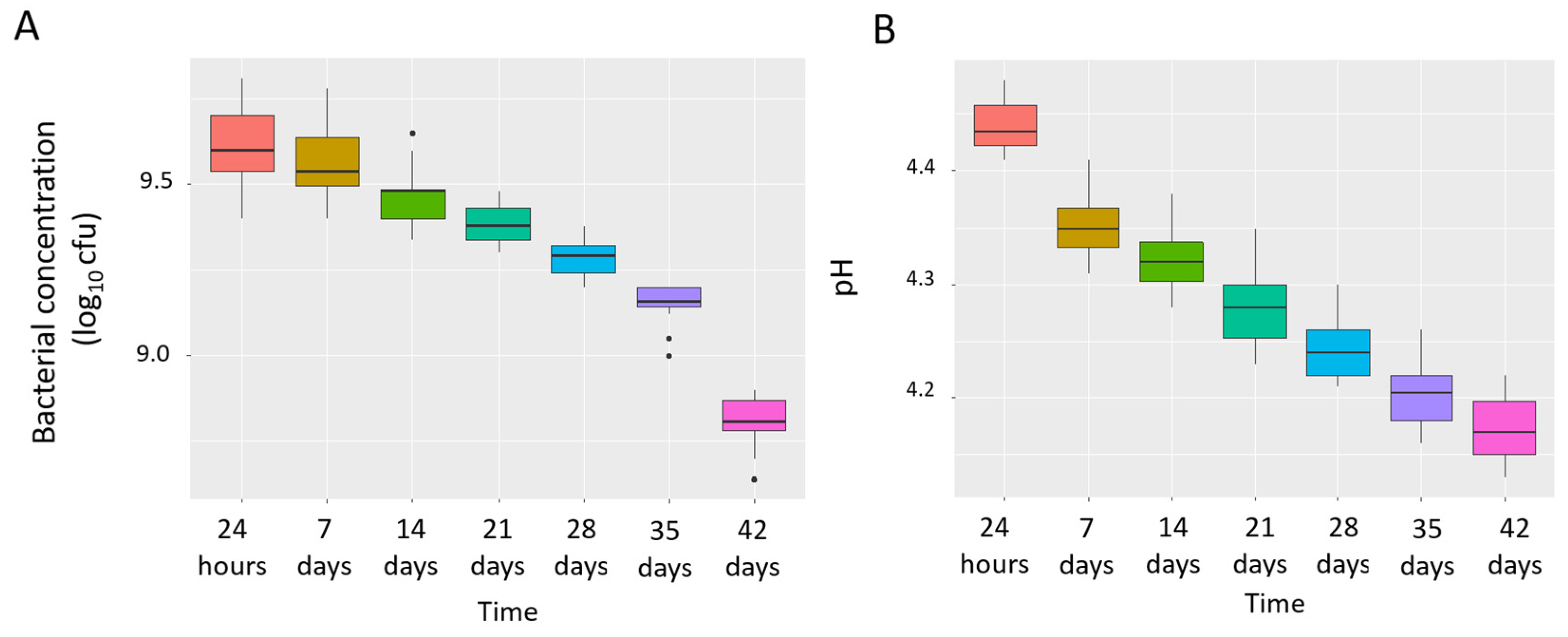
3.1. Concentration, Purity and Stability of L. salivarius MP101 in the Study Product
3.2. Evolution of the COVID-19, Functional, Cognitive and Nutritional Status of the Participants
3.3. Evolution of the Immunological Parameters in the Nasal Samples
3.4. Evolution of the Immunological Parameters in the Fecal Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, T.; Chakraborty, S.; Saha, P.; Mell, B.; Cheng, X.; Yeo, J.-Y.; Mei, X.; Zhou, G.; Mandal, J.; Golonka, R.; et al. Gnotobiotic Rats Reveal That Gut Microbiota Regulates Colonic mRNA of Ace2, the Receptor for SARS-CoV-2 Infectivity. Hypertension 2020, 76, e1–e3. [Google Scholar] [CrossRef]
- Viana, S.D.; Nunes, S.; Reis, F. ACE2 imbalance as a key player for the poor outcomes in COVID-19 patients with age-related comorbidities—Role of gut microbiota dysbiosis. Ageing Res. Rev. 2020, 62, 101123. [Google Scholar] [CrossRef]
- Khatiwada, S.; Subedi, A. Lung microbiome and coronavirus disease 2019 (COVID-19): Possible link and implications. Hum. Microbiome J. 2020, 17, 100073. [Google Scholar] [CrossRef]
- Dhar, D.; Mohanty, A. Gut microbiota and COVID-19 possible link and implications. Virus Res. 2020, 285, 198018. [Google Scholar] [CrossRef] [PubMed]
- McGillivary, G.; Mason, K.M.; Jurcisek, J.A.; Peeples, M.E.; Bakaletz, L.O. Respiratory syncytial virus-induced dysregulation of expression of a mucosal beta-defensin augments colonization of the upper airway by non-typeable Haemophilus influenzae. Cell. Microbiol. 2009, 11, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Mansbach, J.M.; Ajami, N.J.; Espinola, J.A.; Henke, D.M.; Petrosino, J.F.; Piedra, P.A.; Shaw, C.A.; Sullivan, A.F.; Camargo, C.A. Association of nasopharyngeal microbiota profiles with bronchiolitis severity in infants hospitalised for bronchiolitis. Eur. Respir. J. 2016, 48, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Piters, W.D.S.; Heinonen, S.; Hasrat, R.; Bunsow, E.; Smith, B.; Suarez-Arrabal, M.-C.; Chaussabel, D.; Cohen, D.M.; Sanders, E.A.M.; Ramilo, O.; et al. Nasopharyngeal Microbiota, Host Transcriptome, and Disease Severity in Children with Respiratory Syncytial Virus Infection. Am. J. Respir. Crit. Care Med. 2016, 194, 1104–1115. [Google Scholar] [CrossRef]
- He, L.H.; Ren, L.F.; Li, J.F.; Wu, Y.N.; Li, X.; Zhang, L. Intestinal flora as a potential strategy to fight SARS-CoV-2 infection. Front. Microbiol. 2020, 11, 1388. [Google Scholar] [CrossRef]
- He, Y.; Wang, J.; Li, F.; Shi, Y. Main Clinical Features of COVID-19 and Potential Prognostic and Therapeutic Value of the Microbiota in SARS-CoV-2 Infections. Front. Microbiol. 2020, 11, 1302. [Google Scholar] [CrossRef]
- Antunes, A.E.; Vinderola, G.; dos Santos, D.X.; Sivieri, K. Potential contribution of beneficial microbes to face the COVID-19 pandemic. Food Res. Int. 2020, 136, 109577. [Google Scholar] [CrossRef]
- Pagnini, C.; Di Paolo, M.C.; D’Alba, L.; Urgesi, R.; Pallotta, L.; Villotti, G.; D’Ambra, G.; Fave, G.D.; Graziani, M.G. Probiotics in COVID-19 pandemic: When evidence lacks, rationale can still ease the way. Minerva Gastroenterol. Dietol. 2020. [Google Scholar] [CrossRef]
- Sundararaman, A.; Ray, M.; Ravindra, P.V.; Halami, P.M. Role of probiotics to combat viral infections with emphasis on COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 1–16. [Google Scholar] [CrossRef]
- Bottari, B.; Castellone, V.; Neviani, E. Probiotics and COVID-19. Int. J. Food Sci. Nutr. 2021, 72, 293–299. [Google Scholar] [CrossRef] [PubMed]
- d’Ettorre, G.; Ceccarelli, G.; Marazzato, M.; Campagna, G.; Pinacchio, C.; Alessandri, F.; Ruberto, F.; Rossi, G.; Celani, L.; Scagnolari, C.; et al. Challenges in the management of SARS-CoV2 Infection: The role of oral bacteriotherapy as complementary therapeutic strategy to avoid the progression of COVID-19. Front. Med. 2020, 7, 389. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F.; et al. Oral Bacteriotherapy in Patients With COVID-19: A Retrospective Cohort Study. Front. Nutr. 2021, 7, 613928. [Google Scholar] [CrossRef]
- Angurana, S.K.; Bansal, A. Probiotics and COVID-19: Think about the link. Br. J. Nutr. 2020, 14, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Alba, C.; Aparicio, M.; González-Martínez, F.; González-Sánchez, M.I.; Pérez-Moreno, J.; del Castillo, B.T.; Rodríguez, J.M.; Rodríguez-Fernández, R.; Fernández, L. Nasal and Fecal Microbiota and Immunoprofiling of Infants with and Without RSV Bronchiolitis. Front. Microbiol. 2021, 12, 667832. [Google Scholar] [CrossRef] [PubMed]
- Kullen, M.J.; Sanozky-Dawes, R.B.; Crowell, D.C.; Klaenhammer, T.R. Use of the DNA sequence of variable regions of the 16S rRNA gene for rapid and accurate identification of bacteria in the Lactobacillus acidophilus complex. J. Appl. Microbiol. 2000, 89, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Barba, J.L.; Maldonado, A.; Jimenez-Diaz, R. Small-scale total DNA extraction from bacteria and yeast for PCR applications. Anal. Biochem. 2005, 347, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Mansbach, J.M.; Wong, M.C.; Ajami, N.J.; Petrosino, J.F.; Camargo, C.A.; Hasegawa, K. Associations of Nasopharyngeal Metabolome and Microbiome with Severity among Infants with Bronchiolitis. A Multiomic Analysis. Am. J. Respir. Crit. Care Med. 2017, 196, 882–891. [Google Scholar] [CrossRef]
- Mahoney, F.L.; Barthel, D.W. Functional evaluation: The Barthel Index. Md. State Med. J. 1965, 14, 61–65. [Google Scholar]
- Auer, S.; Reisberg, B. The GDS/FAST Staging System. Int. Psychogeriatr. 1997, 9, 167–171. [Google Scholar] [CrossRef]
- Bauer, J.M.; Kaiser, M.J.; Anthony, P.; Guigoz, Y.; Sieber, C.C. The Mini Nutritional Assessment—Its history, today’s practice, and future perspectives. Nutr. Clin. Pract. 2008, 23, 388–396. [Google Scholar] [CrossRef]
- Aparicio, M.; Alba, C.; CAM Public Health Area 6 PSGO; Rodríguez, J.M.; Fernández, L. Microbiological and immunological markers in milk and infant feces for common gastrointestinal disorders: A pilot study. Nutrients 2020, 12, 634. [Google Scholar] [CrossRef]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the gut microbiota in patients with COVID-19 or H1N1 influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.C.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 during Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Neville, B.A.; O’Toole, P.W. Probiotic properties of Lactobacillus salivarius and closely related Lactobacillus species. Future Microbiol. 2010, 5, 759–774. [Google Scholar]
- Fernández, L.; Cárdenas, N.; Arroyo, R.; Manzano, S.; Jiménez, E.; Martín, V.; Rodríguez, J.M. Prevention of Infectious Mastitis by Oral Administration of Lactobacillus salivariusPS2 during Late Pregnancy. Clin. Infect. Dis. 2016, 62, 568–573. [Google Scholar] [CrossRef]
- Martín, V.; Cárdenas, N.; Ocaña, S.; Marín, M.; Arroyo, R.; Beltrán, D.; Badiola, C.; Fernández, L.; Rodríguez, J.M. Rectal and Vaginal Eradication of Streptococcus agalactiae (GBS) in Pregnant Women by Using Lactobacillus salivarius CECT 9145, A Target-specific Probiotic Strain. Nutrients 2019, 11, 810. [Google Scholar] [CrossRef]
- Fernández, L.; Castro, I.; Arroyo, R.; Alba, C.; Beltrán, D.; Rodríguez, J.M. Application of Ligilactobacillus salivarius CECT5713 to Achieve Term Pregnancies in Women with Repetitive Abortion or Infertility of Unknown Origin by Microbiological and Immunological Modulation of the Vaginal Ecosystem. Nutrients 2021, 13, 162. [Google Scholar] [CrossRef]
- Pizarro-Pennarolli, C.; Sánchez-Rojas, C.; Torres-Castro, R.; Vera-Uribe, R.; Sanchez-Ramirez, D.C.; Vasconcello-Castillo, L.; Solís-Navarro, L.; Rivera-Lillo, G. Assessment of activities of daily living in patients post COVID-19: A systematic review. PeerJ 2021, 9, e11026. [Google Scholar] [CrossRef]
- Belli, S.; Balbi, B.; Prince, I.; Cattaneo, D.; Masocco, F.; Zaccaria, S.; Bertalli, L.; Cattini, F.; Lomazzo, A.; Negro, F.D.; et al. Low physical functioning and impaired performance of activities of daily life in COVID-19 patients who survived hospitalisation. Eur. Respir. J. 2020, 56, 2002096. [Google Scholar] [CrossRef]
- Curci, C.; Pisano, F.; Bonacci, E.; Camozzi, D.M.; Ceravolo, C.; Bergonzi, R.; De Franceschi, S.; Moro, P.; Guarnieri, R.; Ferrillo, M.; et al. Early rehabilitation in post-acute COVID-19 patients: Data from an Italian COVID-19 rehabilitation unit and proposal of a treatment protocol. A cross-sectional study. Eur. J. Phys. Rehab. Med. 2020, 56, 633–641. [Google Scholar] [CrossRef]
- Sakai, T.; Hoshino, C.; Yamaguchi, R.; Hirao, M.; Nakahara, R.; Okawa, A. Remote rehabilitation for patients with COVID-19. J. Rehabil. Med. 2020, 52, 00095. [Google Scholar] [CrossRef]
- Silva, D.; Lima, S.; Sena-Evangelista, K.; Marchioni, D.; Cobucci, R.; Andrade, F. Nutritional Risk Screening Tools for Older Adults with COVID-19: A Systematic Review. Nutrients 2020, 12, 2956. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Gong, C.; Wang, J.; Liu, B.; Shi, L.; Duan, J. Prevalence of malnutrition and analysis of related factors in elderly patients with COVID-19 in Wuhan, China. Eur. J. Clin. Nutr. 2020, 74, 871–875. [Google Scholar] [CrossRef]
- Neill, D.; Saint, G.; Bricio-Moreno, L.; Fothergill, J.; Southern, K.W.; Winstanley, C.; Christmas, S.; Slupsky, J.R.; McNamara, P.; Kadioglu, A.; et al. The B Lymphocyte Differentiation Factor (BAFF) Is Expressed in the Airways of Children with CF and in Lungs of Mice Infected with Pseudomonas aeruginosa. PLoS ONE 2014, 9, e95892. [Google Scholar] [CrossRef]
- Reed, J.L.; Welliver, T.P.; Sims, G.P.; McKinney, L.; Velozo, L.; Avendano, L.; Hintz, K.; Luma, J.; Coyle, A.J.; Welliver, S.R.C. Innate Immune Signals Modulate Antiviral and Polyreactive Antibody Responses during Severe Respiratory Syncytial Virus Infection. J. Infect. Dis. 2009, 199, 1128–1138. [Google Scholar] [CrossRef]
- McNamara, P.; Fonceca, A.M.; Howarth, D.; Correia, J.B.; Slupsky, J.R.; Trinick, R.E.; Al Turaiki, W.; Smyth, R.L.; Flanagan, B.F. Respiratory syncytial virus infection of airway epithelial cells, in vivo and in vitro, supports pulmonary antibody responses by inducing expression of the B cell differentiation factor BAFF. Thorax 2012, 68, 76–81. [Google Scholar] [CrossRef]
- Fodor, I.; Serban, O.; Serban, D.E.; Farcau, D.; Man, S.C.; Dumitrascu, D.L. B cell-activating factor (BAFF) in children with inflammatory bowel disease. Pediatr. Res. 2021, 89, 1798–1803. [Google Scholar] [CrossRef]
- Noah, T.L.; Ivins, S.S.; Murphy, P.; Kazachkova, I.; Moats-Staats, B.; Henderson, F.W. Chemokines and Inflammation in the Nasal Passages of Infants with Respiratory Syncytial Virus Bronchiolitis. Clin. Immunol. 2002, 104, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Breindahl, M.; Rieneck, K.; Nielsen, C.; Justesen, T.; Bendtzen, K.; Müller, K. Cytokine responses in infants infected with respiratory syncytial virus. Open J. Immunol. 2012, 2, 40–48. [Google Scholar] [CrossRef]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal Immunol. 2021, 14, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, A.; Pesce, F.; Laterza, R.; D’Alise, M.G.; Lovero, R.; Fontana, A.; Contino, R.; Di Serio, F. Pentraxin 3: Potential prognostic role in SARS-CoV-2 patients admitted to the emergency department. J. Infect. 2021, 82, 84–123. [Google Scholar] [CrossRef] [PubMed]
- Kamle, S.; Ma, B.; He, C.H.; Akosman, B.; Zhou, Y.; Lee, C.M.; El-Deiry, W.S.; Huntington, K.; Liang, O.; Machan, J.; et al. Chitinase 3-like-1 is a therapeutic target that mediates the effects of aging in COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]
- Sims, J.T.; Krishnan, V.; Chang, C.-Y.; Engle, S.M.; Casalini, G.; Rodgers, G.H.; Bivi, N.; Nickoloff, B.J.; Konrad, R.J.; de Bono, S.; et al. Characterization of the cytokine storm reflects hyperinflammatory endothelial dysfunction in COVID-19. J. Allergy Clin. Immunol. 2021, 147, 107–111. [Google Scholar] [CrossRef]
- Garlanda, C.; Bottazzi, B.; Magrini, E.; Inforzato, A.; Mantovani, A. PTX3, a Humoral Pattern Recognition Molecule, in Innate Immunity, Tissue Repair, and Cancer. Physiol. Rev. 2018, 98, 623–639. [Google Scholar] [CrossRef]


{kind=link}
| Participant Characteristic | Mean (95% CI) or n (%) |
|---|---|
| Age (years) | 84.95 (81.41–88.49) |
| Gender | data |
| Male | 11 (50%) |
| Female | 11 (50%) |
| Type of diet | |
| Normal | 10 (45%) |
| Normal/Diabetes | 7 (32%) |
| Pureed (turmix) foods | 5 (23%) |
| BMI (kg/m2) | |
| Day 0 | 24.82 (22.88–26.751) |
| Day 120 | 24.71 (23.27–26.16) |
| SARS-CoV-2 (positive PCR/total) | |
| Day 0 | 18/22 (81%) |
| Day 120 | 0/22 (0%) |
| Index or Score 1 | Day 0 | Day 120 | p-Value |
|---|---|---|---|
| BI | 36.00 [22.75–80.50] | 42.00 [25.25–84.25] | 0.022 |
| GDS/FAST | 4.00 [3.25–6.00] | 4.00 [3.25–6.00] | - |
| MNA | 20.70 (18.80–22.60) | 22.63 (21.20–24.06) | 0.001 |
| Day 0 | Day 120 | |||||
|---|---|---|---|---|---|---|
| Immune Factor | Frequency of Detection | Median (IQR) or Mean (95%CI) | Frequency of Detection | Median (IQR) or Mean (95%CI) | p-Value a | p-Value |
| APRIL/TNFSF13, μg/L | 22 (100%) | 7.35 [3.52–9.47] | 21 (96%) | 6.80 [3.00–8.35] | >0.99 | <0.001 n |
| BAFF/TNFSF13B, μg/L | 22 (100%) | 2.76 (2.35–3.16) | 22 (100%) | 0.89 (0.79–0.99) | >0.99 | <0.001 s |
| Chitinase 3-like 1, μg/L | 17 (77%) | 0.10 [0.68–1.31] | 17 (77%) | 0.95 [0.78–1.26] | >0.99 | 0.040 n |
| IFNα2, ng/L | 2 (9%) | 20.05 | 2 (9%) | 20.40 [19.92–20.87] | >0.99 | - |
| IFNβ, ng/L | 4 (18%) | 2.03 [1.90–2.28] | 1 (5%) | 1.47 | 0.4 | - |
| IL8, ng/L | 18 (82%) | 2.56 [2.42–3.30] | 17 (77%) | 0.49 [0.27–0.60] | >0.99 | <0.001 n |
| IL11, ng/L | 7 (32%) | 0.66 [0.52–0.75] | 0 (0%) | 0 | 0.008 | - |
| IL12p40, ng/L | 4 (18%) | 2.32 [2.14–2.65] | 0 (0%) | 0 | 0.1 | - |
| IL12p70, ng/L | 6 (27%) | 0.13 [0.12–0.13] | 0 (0%) | 0 | 0.02 | - |
| IL19, ng/L | 8 (36%) | 1.55 [0.96–2.45] | 15 (68%) | 4.76 [3.85–9.42] | 0.068 | 0.008 n |
| IL22, ng/L | 2 (9%) | 2.77 | 2 (9%) | 2.76 | >0.99 | - |
| IL26, ng/L | 0 (0%) | 0 | 1 (5%) | 0.46 | >0.99 | - |
| IL32, ng/L | 18 (82%) | 6.37 (5.32–7.43) | 17 (77%) | 3.10 (2.29–3.91) | >0.99 | <0.001 s |
| IL34, ng/L | 6 (27%) | 20.71 [18.72–22.88] | 4 (18%) | 12.5 [11.31–13.17] | 0.72 | - |
| IL35, ng/L | 8 (36%) | 0.64 [0.52–1.09] | 8 (36%) | 1.14 [1.07–2.28] | >0.99 | 0.008 n |
| LIGHT/TNFSF14, ng/L | 13 (59%) | 1.19 (0.93–1.44) | 5 (23%) | 0.63 (0.00–1.35) | 0.03 | 0.684 s |
| MMP-1, ng/L | 6 (27%) | 17.60 [15.00–18.97] | 0 (0%) | 0 | 0.02 | - |
| MMP-2, ng/L | 6 (27%) | 34.69 [24.06–50.05] | 2 (9%) | 14.54 | 0.24 | - |
| MMP-3, ng/L | 4 (18%) | 31.65 [30.05–34.12] | 0 (0%) | 0 | 0.1 | - |
| Osteocalcin, ng/L | 3 (14%) | 2.24 [1.52–2.25] | 1 (5%) | 2.97 | 0.61 | - |
| Osteopontin, ng/L | 22 (100%) | 81.22 (69.51–92.93) | 18 (82%) | 61.74 (49.02–74.47) | 0.1 | <0.001 s |
| Pentraxin 3, ng/L | 7 (77%) | 0.71 [0.63–2.44] | 22 (100%) | 2.75 [2.28–4.22] | <0.001 | 0.016 n |
| TSLP, ng/L | 17 (77%) | 0.73 (0.50–0.96) | 4 (18%) | 0.42 (0.00–1.34) | <0.001 | 0.075 s |
| TWEAK/TNFSF12, ng/L | 17 (77%) | 2.15 [0.72–3.41] | 19 (86%) | 2.28 [0.80–3.44] | 0.69 | 0.579 n |
| gp130/sIL-6Rb, ng/L | 22 (100%) | 155.88 (125.89–185.88) | 20 (91%) | 157.79 (123.62–191.96) | 0.48 | 0.154 s |
| sCD30/TNFRSF8, ng/L | 4 (18%) | 40.84 [14.45–65.42] | 4 (18%) | 44.02 [19.45–68.21] | >0.99 | - |
| sCD163, ng/L | 0 (0%) | 0 | 4 (18%) | 2.88 [2.22–3.69] | 0.1 | - |
| sIL-6Ra, ng/L | 4 (18%) | 10.75 (0–26.46) | 8 (36%) | 11.47 (1.17–21.76) | 0.31 | 0.351 s |
| sTNF-R1, ng/L | 21 (96%) | 34.64 (22.76–46.51) | 13 (59%) | 13.79 (2.58–25.01) | 0.009 | <0.001 s |
| sTNF-R2, ng/L | 21 (96%) | 12.82 (9.43–16.21) | 6 (27%) | 4.50 (1.62–7.38) | <0.001 | 0.212 s |
| Day 0 | Day 120 | |||||
|---|---|---|---|---|---|---|
| Immune Factor | Frequency of Detection | Median (IQR) or Mean (95%CI) | Frequency of Detection | Median (IQR) or Mean (95%CI) | p-Value a | p-Value |
| APRIL/TNFSF13, μg/L | 12 (55%) | 1.72 (1.22–2.23) | 11 (50%) | 1.48 (0.99–1.97) | >0.99 | 0.0111 s |
| BAFF/TNFSF13B, μg/L | 22 (100%) | 1.21 [0.96–1.51] | 22 (100%) | 0.49 [0.31–0.68] | >0.99 | <0.001 n |
| Chitinase 3-like 1, μg/L | 21 (95%) | 0.35 [0.21–0.56] | 22 (100%) | 0.31 [0.20–0.44] | >0.99 | 0.0822 n |
| IFNα2, ng/L | 0 (0%) | 0 | 5 (23%) | 15.56 [12.12–17.87] | 0.048 | - |
| IFNβ, ng/L | 6 (27%) | 2.72 (1.52–3.92) | 9 (41%) | 2.82 (1.81–3.83) | 0.52 | 0.407 s |
| IFNγ, ng/L | 4 (18%) | 5.42 [4.09–5.81] | 0 (0%) | 0 | 0.100 | - |
| IL8, ng/L | 8 (36%) | 3.79 [2.20–6.34] | 7 (32%) | 1.15 [0.51–2.27] | >0.99 | - |
| IL11, ng/L | 5 (23%) | 0.36 (0.24–0.49) | 5 (23%) | 0.36 (0.23–0.49) | >0.99 | 0.825 s |
| IL12p40, ng/L | 8 (36%) | 4.99 (1.89–8.09) | 9 (41%) | 4.29 (1.59–6.98) | >0.99 | 0.845 s |
| IL12p70, ng/L | 11 (50%) | 0.21 (0.11–0.31) | 10 (45%) | 0.22 (0.09–0.35) | >0.99 | 0.823 s |
| IL19, ng/L | 7 (32%) | 1.60 [1.19–2.37] | 14 (64%) | 4.05 [3.54–4.88] | 0.21 | 0.016 n |
| IL20, ng/L | 1 (5%) | 0.32 | 0 (0%) | 0 | >0.99 | - |
| IL22, ng/L | 8 (36%) | 2.37 (1.21–3.54) | 10 (45%) | 1.39 (0.66–2.12) | 0.75 | 0.079 s |
| IL26, ng/L | 11 (50%) | 1.28 (0.41–2.14) | 12 (55%) | 0.91 (0.41–1.41) | >0.99 | 0.675 s |
| IL27p28, ng/L | 9 (41%) | 0.55 [0.44–3.76] | 8 (36%) | 0.37 [0.21–0.87] | >0.99 | 0.813 n |
| IL28A/IFNλ2, ng/L | 3 (14%) | 4.52 [4.225–4.78] | 3 (14%) | 0.73 [0.565–1.91] | >0.99 | - |
| IL32, ng/L | 21 (95%) | 6.50 [5.89–8.09] | 20 (91%) | 3.47 [2.67–5.10] | >0.99 | <0.001 n |
| IL34, ng/L | 16 (73%) | 40.57 (30.55–50.59) | 16 (73%) | 19.19 (11.03–27.35) | >0.99 | 0.002 s |
| IL35, ng/L | 16 (73%) | 7.79 [4.23–10.23] | 16 (73%) | 10.33 [6.39–15.56] | >0.99 | 0.005 n |
| LIGHT/TNFSF14, ng/L | 12 (55%) | 1.39 (0.81–1.98) | 14 (64%) | 0.87 (0.43–1.31) | 0.750 | 0.228 s |
| MMP-1, ng/L | 7 (32%) | 26.85 [22.37–89.33] | 3 (14%) | 6.30 [3.96–20.34] | 0.280 | - |
| MMP-2, ng/L | 10 (45%) | 50.10 (23.35–76.86) | 6 (27%) | 70.08 (44.45–95.72) | 0.340 | 0.016 s |
| MMP-3, ng/L | 4 (18%) | 36.61 [29.21–59.21] | 1 (5%) | 23.39 | 0.340 | - |
| Osteocalcin, ng/L | 2 (9%) | 3.27 | 4 (18.18%) | 2.58 [1.99–3.71] | 0.660 | - |
| Osteopontin, ng/L | 22 (100%) | 88.61 [75.96–100.07] | 21 (95%) | 80.70 [74.87–94.65] | >0.99 | 0.128 n |
| Pentraxin 3, ng/L | 12 (55%) | 0.97 (0.39–1.55) | 22 (100%) | 2.84 (2.38–3.30) | 0.34 | <0.001 s |
| TSLP, ng/L | 19 (86%) | 0.94 [0.73- 1.94] | 16 (73%) | 0.86 [0.60–1.78] | 0.450 | 0.551 n |
| TWEAK/TNFSF12, ng/L | 21 (95%) | 3.19 [2.32–4.23] | 21 (95%) | 3.69 [2.62–5.01] | >0.99 | 0.063 n |
| gp130/sIL-6Rb, ng/L | 20 (91%) | 79.57 (67.48 91.65) | 20 (91%) | 71.80 (60.79–82.81) | >0.99 | <0.001 s |
| sCD30/TNFRSF8, ng/L | 11 (50%) | 22.41 [13.51–40.15] | 13 (59%) | 14.87 [4.45–37.20] | 0.76 | 0.831 n |
| sCD163, ng/L | 4 (18%) | 5.07(3.72–6.42) | 5 (23%) | 6.57 (5.72–7.42) | >0.99 | 0.037 s |
| sIL-6Ra, ng/L | 1 (5%) | 10.03 | 6 (27%) | 2.69 [2.35–3.2625] | 0.090 | - |
| sTNF-R1, ng/L | 22 (100%) | 29.62 [19.33–36.86] | 16 (73%) | 4.98 [1.13–9.14] | 0.020 | <0.001 n |
| sTNF-R2, ng/L | 22 (100%) | 19.72 (15.22–24.23) | 14 (64%) | 3.25 (1.18–5.33) | 0.003 | <0.001 s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mozota, M.; Castro, I.; Gómez-Torres, N.; Arroyo, R.; Lailla, Y.; Somada, M.; Alba, C.; Rodríguez, J.M. Administration of Ligilactobacillus salivarius MP101 in an Elderly Nursing Home during the COVID-19 Pandemic: Immunological and Nutritional Impact. Foods 2021, 10, 2149. https://doi.org/10.3390/foods10092149
Mozota M, Castro I, Gómez-Torres N, Arroyo R, Lailla Y, Somada M, Alba C, Rodríguez JM. Administration of Ligilactobacillus salivarius MP101 in an Elderly Nursing Home during the COVID-19 Pandemic: Immunological and Nutritional Impact. Foods. 2021; 10(9):2149. https://doi.org/10.3390/foods10092149
Chicago/Turabian StyleMozota, Marta, Irma Castro, Natalia Gómez-Torres, Rebeca Arroyo, Yolanda Lailla, Mario Somada, Claudio Alba, and Juan Miguel Rodríguez. 2021. "Administration of Ligilactobacillus salivarius MP101 in an Elderly Nursing Home during the COVID-19 Pandemic: Immunological and Nutritional Impact" Foods 10, no. 9: 2149. https://doi.org/10.3390/foods10092149
APA StyleMozota, M., Castro, I., Gómez-Torres, N., Arroyo, R., Lailla, Y., Somada, M., Alba, C., & Rodríguez, J. M. (2021). Administration of Ligilactobacillus salivarius MP101 in an Elderly Nursing Home during the COVID-19 Pandemic: Immunological and Nutritional Impact. Foods, 10(9), 2149. https://doi.org/10.3390/foods10092149