
Intensive Environmental Surveillance Plan for Listeria monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity

 , , , , , , , , , , and
, , , , , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling and L. monocytogenes Detection
2.2. Statistical Analysis
2.3. Strains Collection
2.4. Whole Genome Sequencing and Bioinformatics Analysis
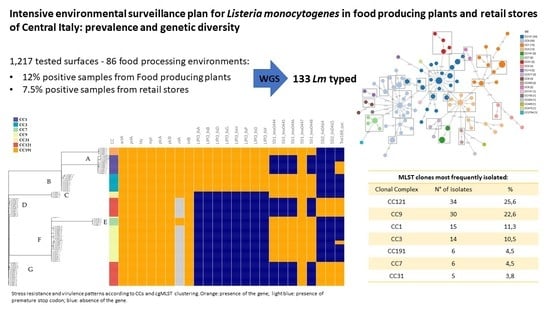
3. Results
3.1. Sampling and L. monocytogenes Detection
3.2. Strains Collection
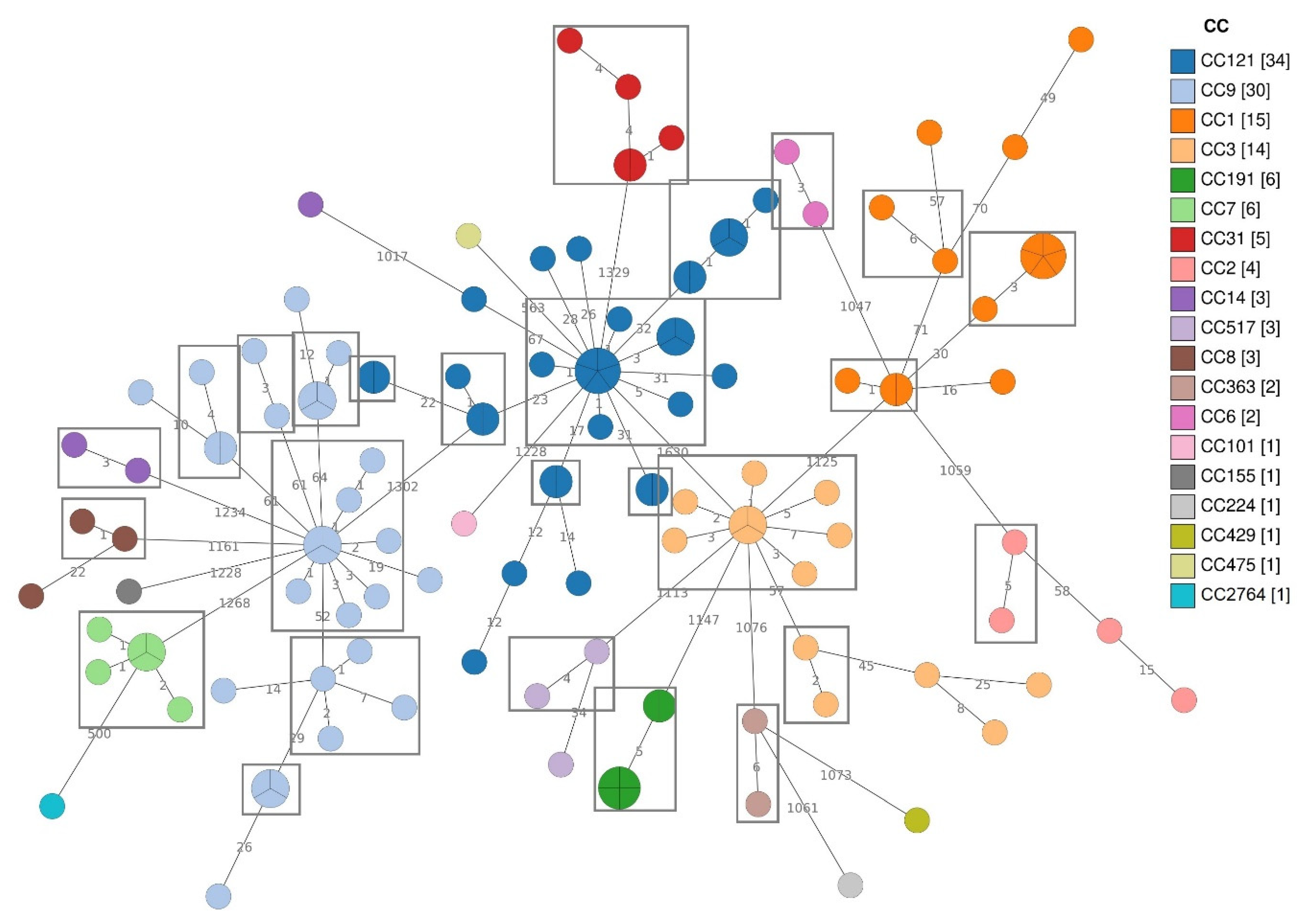
3.3. WGS and Bioinformatics Analysis
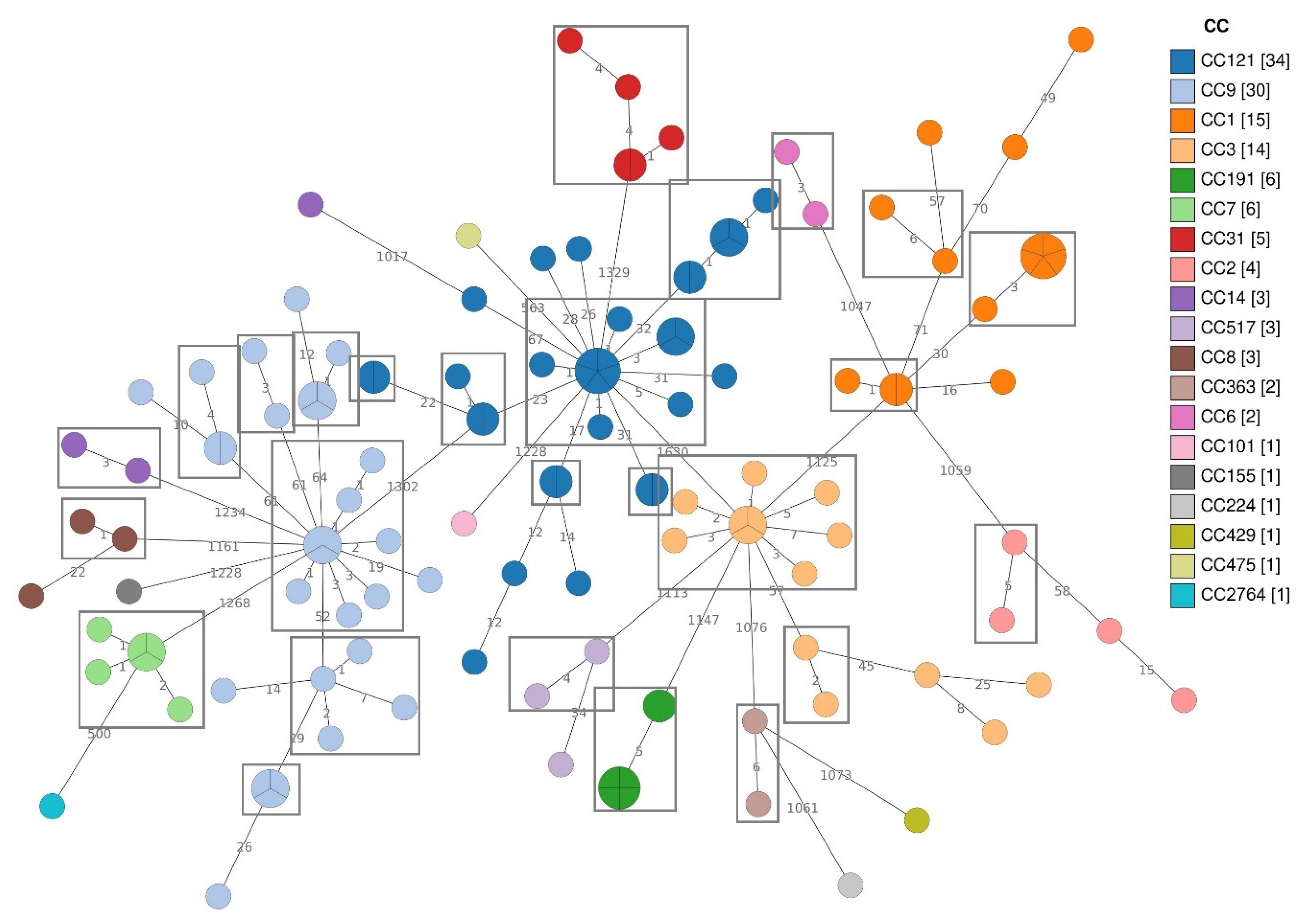
3.3.1. Distribution of CCs and cg-MLST Clusters
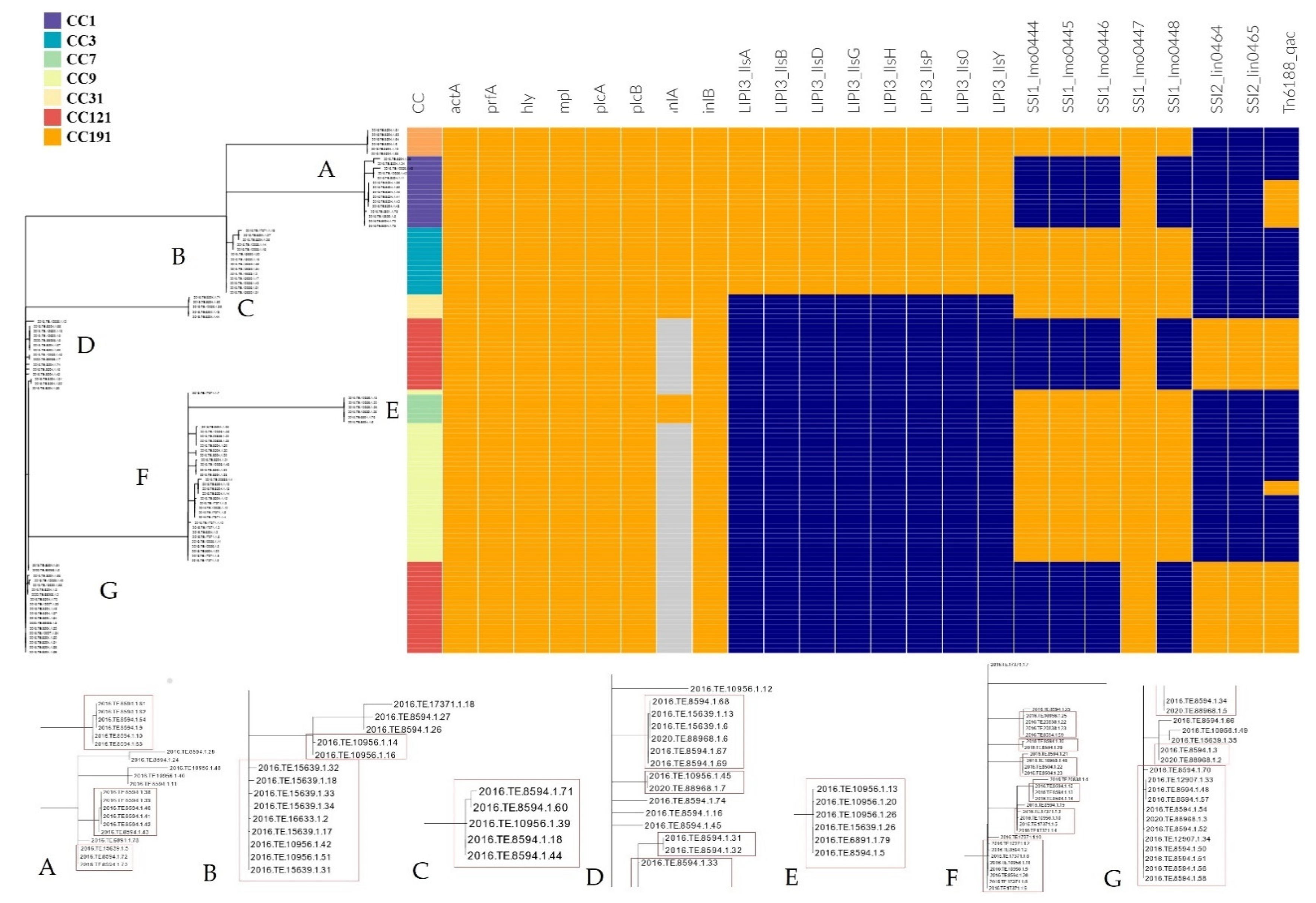
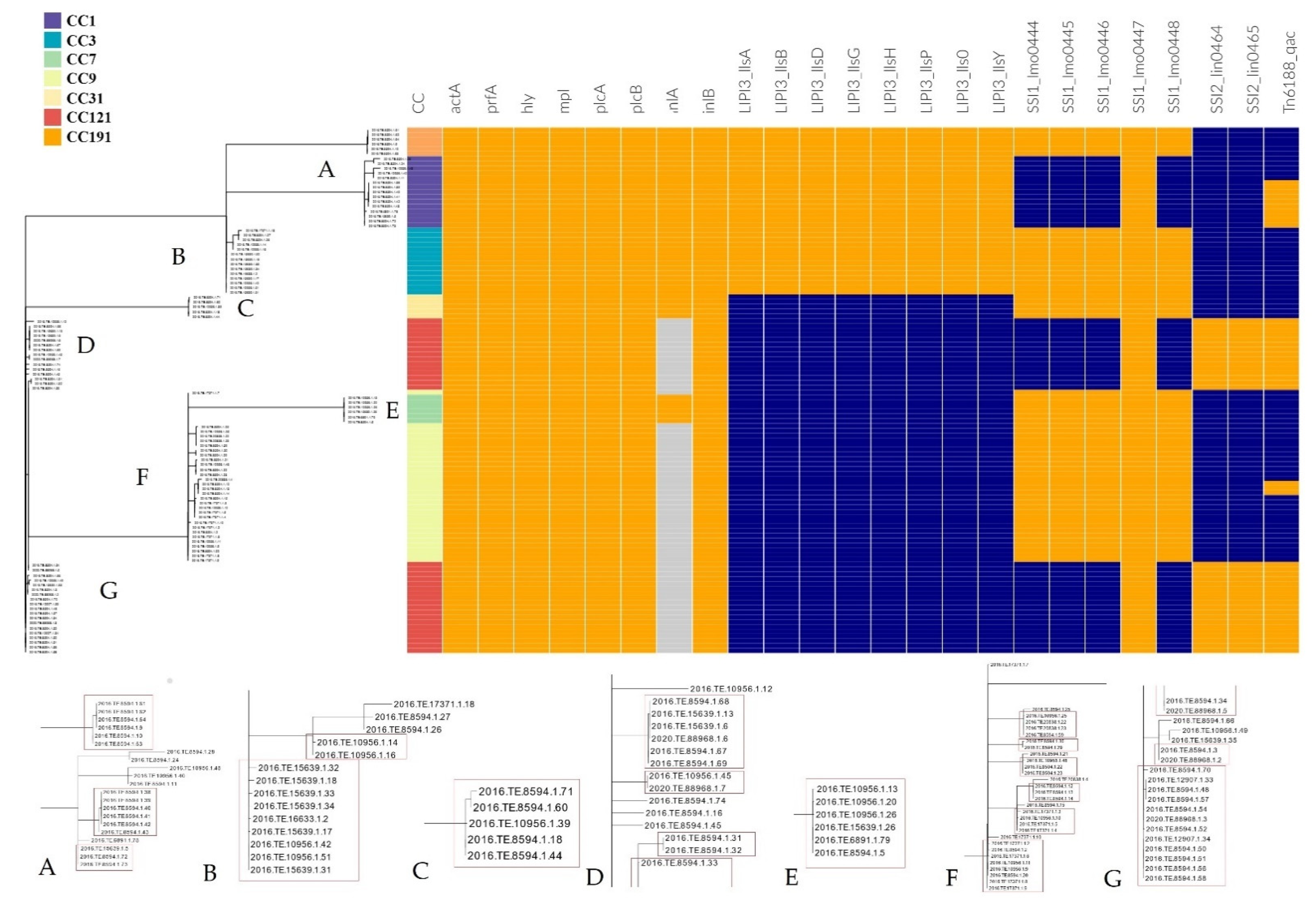
3.3.2. Detection of Stress Resistance and Virulence Genes
4. Discussion
4.1. Sampling and L. monocytogenes Detection
4.2. Distribution of CCs and cg-MLST Clusters and Their Virulence and Stress Resistance Profiles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camargo, A.C.; Moura, A.; Avillan, J.; Herman, N.; McFarland, A.P.; Sreevatsan, S.; Call, D.R.; Woodward, J.J.; Lecuit, M.; Nero, L.A. Whole-genome Sequencing Reveals Listeria monocytogenes Diversity and Allows Identification of Long-term Persistent Strains in Brazil. Environ. Microbiol. 2019, 21, 4478–4487. [Google Scholar] [CrossRef]
- Disson, O.; Moura, A.; Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria Monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Stoller, A.; Stevens, M.; Stephan, R.; Guldimann, C. Characteristics of Listeria Monocytogenes Strains Persisting in a Meat Processing Facility over a 4-Year Period. Pathogens 2019, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Paul, M. Monitoring and Analytics of Listeria Monocytogenes: Key Facts for Authorities, Food Manufacturers, and Retailers. J. Consum. Prot. Food Saf. 2020, 15, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Guidi, F.; Orsini, M.; Chiaverini, A.; Torresi, M.; Centorame, P.; Acciari, V.A.; Salini, R.; Palombo, B.; Brandi, G.; Amagliani, G.; et al. Hypo- and Hyper-Virulent Listeria Monocytogenes Clones Persisting in Two Different Food Processing Plants of Central Italy. Microorganisms 2021, 9, 376. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria Monocytogenes Contamination of Ready-to-eat Foods and the Risk for Human Health in the EU. EFSA J. 2018, 16, 5134. [Google Scholar] [CrossRef]
- Demaître, N.; Van Damme, I.; De Zutter, L.; Geeraerd, A.H.; Rasschaert, G.; De Reu, K. Occurrence, Distribution and Diversity of Listeria Monocytogenes Contamination on Beef and Pig Carcasses after Slaughter. Meat Sci. 2020, 169, 108177. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (EFSA BIOHAZ Panel); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Whole Genome Sequencing and Metagenomics for Outbreak Investigation, Source Attribution and Risk Assessment of Food-borne Microorganisms. EFSA J. 2019, 17, e05898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maury, M.M.; Tsai, Y.-H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria Monocytogenes Hypervirulence by Harnessing Its Biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Kallipolitis, B.; Gahan, C.G.; Piveteau, P. Factors Contributing to Listeria Monocytogenes Transmission and Impact on Food Safety. Curr. Opin. Food Sci. 2020, 36, 9–17. [Google Scholar] [CrossRef]
- Duranti, A.; Sabbatucci, M.; Blasi, G.; Acciari, V.A.; Ancora, M.; Bella, A.; Busani, L.; Centorame, P.; Cammà, C.; Conti, F.; et al. A Severe Outbreak of Listeriosis in Central Italy with a Rare Pulsotype Associated with Processed Pork Products. J. Med. Microbiol. 2018, 67, 1351–1360. [Google Scholar] [CrossRef]
- Carpentier, B.; Barre, L. Guidelines on Sampling the Food Processing Area and Equipment for the Detection of Listeria Monocytogenes. Fr. Agency Food Environ. Occup. Health Saf. 2012, 1–15. [Google Scholar]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the Major Listeria Monocytogenes Serovars by Multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [Green Version]
- Kérouanton, A.; Marault, M.; Petit, L.; Grout, J.; Dao, T.T.; Brisabois, A. Evaluation of a Multiplex PCR Assay as an Alternative Method for Listeria Monocytogenes Serotyping. J. Microbiol. Methods 2010, 80, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Cito, F.; Di Pasquale, A.; Cammà, C.; Cito, P. The Italian Information System for the Collection and Analysis of Complete Genome Sequence of Pathogens Isolated from Animal, Food and Environment. Int. J. Infect. Dis. 2018, 73, 296–297. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Salcedo, C.; Arreaza, L.; Alcalá, B.; de la Fuente, L.; Vázquez, J.A. Development of a Multilocus Sequence Typing Method for Analysis of Listeria monocytogenes Clones. J. Clin. Microbiol. 2003, 41, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. ChewBBACA: A Complete Suite for Gene-by-Gene Schema Creation and Strain Identification. Microbial. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef]
- Moura, A.; Tourdjman, M.; Leclercq, A.; Hamelin, E.; Laurent, E.; Fredriksen, N.; Van Cauteren, D.; Bracq-Dieye, H.; Thouvenot, P.; Vales, G.; et al. Real-Time Whole-Genome Sequencing for Surveillance of Listeria monocytogenes, France. Emerg. Infect. Dis. 2017, 23, 1462–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of Core Genomic Relationships among 100,000 Bacterial Pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An Interactive Viewer for Bacterial Population Genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q. Heterogeneity, Characteristics, and Public Health Implications of Listeria Monocytogenes in Ready-to-Eat Foods and Pasteurized Milk in China. Front. Microbiol. 2020, 11, 10. [Google Scholar]
- Hoelzer, K.; Sauders, B.D.; Sanchez, M.D.; Olsen, P.T.; Pickett, M.M.; Mangione, K.J.; Rice, D.H.; Corby, J.; Stich, S.; Fortes, E.D.; et al. Prevalence, Distribution, and Diversity of Listeria Monocytogenes in Retail Environments, Focusing on Small Establishments and Establishments with a History of Failed Inspections. J. Food Prot. 2011, 74, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.; Wu, S.T.; den Bakker, H.C.; Cook, P.W.; Veenhuizen, D.R.; Hammons, S.R.; Singh, M.; Oliver, H.F. Listeria Monocytogenes Is Prevalent in Retail Produce Environments but Salmonella Enterica Is Rare. Food Control 2020, 113, 107173. [Google Scholar] [CrossRef]
- Reinhard, R.G.; Kalinowski, R.M.; Bodnaruk, P.W.; Eifert, J.D.; Boyer, R.R.; Duncan, S.E.; Bailey, R.H. Incidence of Listeria Spp. in Ready-to-Eat Food Processing Plant Environments Regulated by the U.S. Food Safety and Inspection Service and the U.S. Food and Drug Administration. J. Food Prot. 2018, 81, 1063–1067. [Google Scholar] [CrossRef]
- Antoci, S.; Iannetti, L.; Centorotola, G.; Acciari, V.A.; Pomilio, F.; Daminelli, P.; Romanelli, C.; Ciorba, A.B.; Santini, N.; Torresi, M.; et al. Monitoring Italian Establishments Exporting Food of Animal Origin to Third Countries: SSOP Compliance and Listeria Monocytogenes and Salmonella Spp. Contamination. Food Control 2021, 121, 107584. [Google Scholar] [CrossRef]
- Li, H.; Wang, P.; Lan, R.; Luo, L.; Cao, X.; Wang, Y.; Wang, Y.; Li, H.; Zhang, L.; Ji, S.; et al. Risk Factors and Level of Listeria Monocytogenes Contamination of Raw Pork in Retail Markets in China. Front. Microbiol. 2018, 9, 1090. [Google Scholar] [CrossRef]
- Bērziņš, A.; Hellström, S.; Siliņš, I.; Korkeala, H. Contamination Patterns of Listeria Monocytogenes in Cold-Smoked Pork Processing. J. Food Prot. 2010, 73, 2103–2109. [Google Scholar] [CrossRef] [PubMed]
- Forauer, E.; Wu, S.T.; Etter, A.J. Listeria Monocytogenes in the Retail Deli Environment: A Review. Food Control 2021, 119, 107443. [Google Scholar] [CrossRef]
- Sauders, B.D.; Sanchez, M.D.; Rice, D.H.; Corby, J.; Stich, S.; Fortes, E.D.; Roof, S.E.; Wiedmann, M. Prevalence and Molecular Diversity of Listeria Monocytogenes in Retail Establishments. J. Food Prot. 2009, 72, 2337–2349. [Google Scholar] [CrossRef] [PubMed]
- Etter, A.J.; Hammons, S.R.; Roof, S.; Simmons, C.; Wu, T.; Cook, P.W.; Katubig, A.; Stasiewicz, M.J.; Wright, E.; Warchocki, S.; et al. Enhanced Sanitation Standard Operating Procedures Have Limited Impact on Listeria Monocytogenes Prevalence in Retail Delis. J. Food Prot. 2017, 80, 1903–1912. [Google Scholar] [CrossRef]
- D’Arrigo, M.; Mateo-Vivaracho, L.; Guillamón, E.; Fernández-León, M.F.; Bravo, D.; Peirotén, Á.; Medina, M.; García-Lafuente, A. Characterization of Persistent Listeria Monocytogenes Strains from Ten Dry-Cured Ham Processing Facilities. Food Microbiol. 2020, 92, 103581. [Google Scholar] [CrossRef]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Zeng, H.; Xue, L.; Lei, T.; Pang, R.; Wu, S.; Wu, H.; et al. Isolation, Potential Virulence, and Population Diversity of Listeria Monocytogenes From Meat and Meat Products in China. Front. Microbiol. 2019, 10, 946. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Niu, Y.; Liu, Y.; Lu, Z.; Wang, D.; Cui, X.; Chen, Q.; Ma, X. Isolation and Characterization of Clinical Listeria Monocytogenes in Beijing, China, 2014–2016. Front. Microbiol. 2019, 10, 981. [Google Scholar] [CrossRef]
- Leong, D.; Alvarez-Ordóñez, A.; Jordan, K. Monitoring Occurrence and Persistence of Listeria Monocytogenes in Foods and Food Processing Environments in the Republic of Ireland. Front. Microbiol. 2014, 5, 436. [Google Scholar] [CrossRef] [PubMed]
- Martín, B.; Perich, A.; Gómez, D.; Yangüela, J.; Rodríguez, A.; Garriga, M.; Aymerich, T. Diversity and Distribution of Listeria Monocytogenes in Meat Processing Plants. Food Microbiol. 2014, 44, 119–127. [Google Scholar] [CrossRef]
- Meloni, D. Presence of Listeria Monocytogenes in Mediterranean-Style Dry Fermented Sausages. Foods 2015, 4, 34–50. [Google Scholar] [CrossRef] [Green Version]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria Monocytogenes Clones’ Adaption to Mammalian Gut Accounts for Their Association with Dairy Products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef] [Green Version]
- Parisi, A.; Latorre, L.; Normanno, G.; Miccolupo, A.; Fraccalvieri, R.; Lorusso, V.; Santagada, G. Amplified Fragment Length Polymorphism and Multi-Locus Sequence Typing for High-Resolution Genotyping of Listeria Monocytogenes from Foods and the Environment. Food Microbiol. 2010, 27, 101–108. [Google Scholar] [CrossRef]
- Wieczorek, K.; Bomba, A.; Osek, J. Whole-Genome Sequencing-Based Characterization of Listeria Monocytogenes from Fish and Fish Production Environments in Poland. IJMS 2020, 21, 9419. [Google Scholar] [CrossRef] [PubMed]
- Harter, E.; Wagner, E.M.; Zaiser, A.; Halecker, S.; Wagner, M.; Rychli, K. Stress Survival Islet 2, Predominantly Present in Listeria Monocytogenes Strains of Sequence Type 121, Is Involved in the Alkaline and Oxidative Stress Responses. Appl. Environ. Microbiol. 2017, 83, e00827-17. [Google Scholar] [CrossRef] [Green Version]
- Félix, B. Population Genetic Structure of Listeria Monocytogenes Strains Isolated From the Pig and Pork Production Chain in France. Front. Microbiol. 2018, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Luo, L.; Zhang, Z.; Deng, J.; Wang, Y.; Miao, Y.; Zhang, L.; Chen, X.; Liu, X.; Sun, S.; et al. Prevalence and Molecular Characteristics of Listeria Monocytogenes in Cooked Products and Its Comparison with Isolates from Listeriosis Cases. Front. Med. 2018, 12, 104–112. [Google Scholar] [CrossRef]
- Matle, I.; Mafuna, T.; Madoroba, E.; Mbatha, K.R.; Magwedere, K.; Pierneef, R. Population Structure of Non-ST6 Listeria Monocytogenes Isolated in the Red Meat and Poultry Value Chain in South Africa. Microorganisms 2020, 8, 1152. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Haendiges, J.; Keller, E.N.; Myers, R.; Kim, A.; Lombard, J.E.; Karns, J.S.; Van Kessel, J.A.S.; Haley, B.J. Genetic Diversity and Virulence Profiles of Listeria Monocytogenes Recovered from Bulk Tank Milk, Milk Filters, and Milking Equipment from Dairies in the United States (2002 to 2014). PLoS ONE 2018, 13, e0197053. [Google Scholar] [CrossRef] [PubMed]
- Papić, B.; Pate, M.; Félix, B.; Kušar, D. Genetic Diversity of Listeria Monocytogenes Strains in Ruminant Abortion and Rhombencephalitis Cases in Comparison with the Natural Environment. BMC Microbiol. 2019, 19, 299. [Google Scholar] [CrossRef] [Green Version]
- Orsini, M.; Torresi, M.; Patavino, C.; Centorame, P.; Rinaldi, A.; Acciari, V.A.; Centorotola, G.; Marcacci, M.; Ancora, M.; Di Domenico, M.; et al. Whole-Genome Sequence of a Reemerging Listeria Monocytogenes Serovar 1/2a Strain in Central Italy. Microbiol. Resour. Announc. 2018, 7, e01069-18. [Google Scholar] [CrossRef] [Green Version]
- Kurpas, M.; Osek, J.; Moura, A.; Leclercq, A.; Lecuit, M.; Wieczorek, K. Genomic Characterization of Listeria Monocytogenes Isolated From Ready-to-Eat Meat and Meat Processing Environments in Poland. Front. Microbiol. 2020, 11, 1412. [Google Scholar] [CrossRef]
- Cardenas-Alvarez, M.X.; Townsend Ramsett, M.K.; Malekmohammadi, S.; Bergholz, T.M. Evidence of Hypervirulence in Listeria Monocytogenes Clonal Complex 14. J. Med. Microbiol. 2019, 68, 1677–1685. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, W.; Wang, J.; Xu, B.; Liu, H.; Dong, Q.; Zhang, X. 10-Year Molecular Surveillance of Listeria Monocytogenes Using Whole-Genome Sequencing in Shanghai, China, 2009–2019. Front. Microbiol. 2020, 11, 551020. [Google Scholar] [CrossRef]
- Ziegler, M.; Jang, H.; Gopinath, G.; Horlbog, J.A.; Stephan, R.; Guldimann, C. Whole-Genome Shotgun Sequencing of Three Listeria Monocytogenes Strains Isolated from a Ready-to-Eat Salad-Producing Facility in Switzerland. Genome Announc. 2018, 6, e00547-18. [Google Scholar] [CrossRef] [Green Version]
- Painset, A.; Björkman, J.T.; Kiil, K.; Guillier, L.; Mariet, J.-F.; Félix, B.; Amar, C.; Rotariu, O.; Roussel, S.; Perez-Reche, F.; et al. LiSEQ–Whole-Genome Sequencing of a Cross-Sectional Survey of Listeria Monocytogenes in Ready-to-Eat Foods and Human Clinical Cases in Europe. Microbial. Genom. 2019, 5, e000257. [Google Scholar] [CrossRef] [PubMed]
- Gelbicova, T.; Florianova, M.; Hluchanova, L.; Kalova, A.; Korena, K.; Strakova, N.; Karpiskova, R. Comparative Analysis of Genetic Determinants Encoding Cadmium, Arsenic, and Benzalkonium Chloride Resistance in Listeria Monocytogenes of Human, Food, and Environmental Origin. Front. Microbiol. 2021, 11, 599882. [Google Scholar] [CrossRef] [PubMed]
- Hilliard, A.; Leong, D.; O’Callaghan, A.; Culligan, E.; Morgan, C.; DeLappe, N.; Hill, C.; Jordan, K.; Cormican, M.; Gahan, C. Genomic Characterization of Listeria Monocytogenes Isolates Associated with Clinical Listeriosis and the Food Production Environment in Ireland. Genes 2018, 9, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupp, S.; Bärtschi, M.; Frey, J.; Oevermann, A. Hyperinvasiveness and Increased Intercellular Spread of Listeria Monocytogenes Sequence Type 1 Are Independent of Listeriolysin S, Internalin F and Internalin J1. J. Med. Microbiol. 2017, 66, 1053–1062. [Google Scholar] [CrossRef]
- Su, X.; Cao, G.; Zhang, J.; Pan, H.; Zhang, D.; Kuang, D.; Yang, X.; Xu, X.; Shi, X.; Meng, J. Characterization of Internalin Genes in Listeria Monocytogenes from Food and Humans, and Their Association with the Invasion of Caco-2 Cells. Gut Pathog. 2019, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Manuel, C.S.; Van Stelten, A.; Wiedmann, M.; Nightingale, K.K.; Orsi, R.H. Prevalence and Distribution of Listeria Monocytogenes InlA Alleles Prone to Phase Variation and InlA Alleles with Premature Stop Codon Mutations among Human, Food, Animal, and Environmental Isolates. Appl. Environ. Microbiol. 2015, 81, 8339–8345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Melo Tavares, R.; da Silva, D.A.L.; Camargo, A.C.; Yamatogi, R.S.; Nero, L.A. Interference of the Acid Stress on the Expression of LlsX by Listeria Monocytogenes Pathogenic Island 3 (LIPI-3) Variants. Food Res. Int. 2020, 132, 109063. [Google Scholar] [CrossRef] [PubMed]
- Vilchis-Rangel, R.E.; del Rosario Espinoza-Mellado, M.; Salinas-Jaramillo, I.J.; Martinez-Peña, M.D.; Rodas-Suárez, O.R. Association of Listeria Monocytogenes LIPI-1 and LIPI-3 Marker LlsX with Invasiveness. Curr. Microbiol. 2019, 76, 637–643. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Surfaces | FCS | NFCS | ||||||
|---|---|---|---|---|---|---|---|---|
| Food Producing Environments | n | Lm+ | % | CI 95% | n | Lm+ | % | CI 95% |
| FPP | 379 | 49 | 12.9 | 9.9–16.7 | 222 | 23 | 10.4 | 7.0–15.1 |
| RS | 409 | 27 | 6.6 | 4.6–9.4 | 207 | 19 | 9.2 | 6.0–13.9 |
| Total | 788 | 76 | 9.6 | 7.8–11.9 | 429 | 42 | 9.8 | 7.3–13.0 |
| FCS | NFCS | |||||||
|---|---|---|---|---|---|---|---|---|
| Surface Category | n | Lm+ | % | CI 95% | n | Lm+ | % | CI 95% |
| Equipment | 346 | 39 | 11.3 | 8.4–15.0 | 83 | 6 | 7.2 | 3.4–14.9 |
| Industrial systems | 64 | 5 | 7.8 | 3.4–17.0 | 267 | 29 | 10.9 | 7.7–15.2 |
| Machines | 259 | 20 | 7.7 | 5.0–11.6 | 64 | 4 | 6.3 | 2.5–15.0 |
| Clothing | 5 | 1 | 20.0 | 3.6–62.5 | 9 | 3 | 33.3 | 12.1–64.6 |
| Cleaning tools | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Tools | 114 | 11 | 9.6 | 5.5–16.5 | 3 | 0 | 0 | 0 |
| Not classifiable surface | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Total | 788 | 76 | 9.6 | 7.8–11.9 | 429 | 42 | 9.8 | 7.3–12.9 |
| FPP | RS | |||||||
|---|---|---|---|---|---|---|---|---|
| Surface Category | n | Lm+ | % | CI 95% | n | Lm+ | % | CI 95% |
| Equipment | 221 | 24 | 10.9 | 7.4–15.7 | 208 | 21 | 10.1 | 6.7–14.9 |
| Industrial systems | 156 | 22 | 14.1 | 9.5–20.4 | 175 | 12 | 6.9 | 4.0–11.6 |
| Machines | 162 | 16 | 9.9 | 6.2–15.4 | 161 | 8 | 5.0 | 2.5–9.5 |
| Clothing | 7 | 1 | 14.2 | 2.6–51.3 | 7 | 3 | 42.9 | 15.8–75.0 |
| Tools | 52 | 9 | 17.3 | 9.4–29.7 | 65 | 2 | 3.1 | 0.9–10.5 |
| Total | 598 | 72 | 12.0 | 9.7–14.9 | 616 | 46 | 7.5 | 5.7–9.8 |
| CC | cgMLST Clusters | No. of Isolates | No. of Singleton Strains |
|---|---|---|---|
| CC121 | 6 | 12 | 7 |
| 6 | |||
| 3 | |||
| 2 | |||
| 2 | |||
| 2 | |||
| CC9 | 6 | 9 | 5 |
| 4 | |||
| 4 | |||
| 3 | |||
| 3 | |||
| 2 | |||
| CC1 | 3 | 6 | 4 |
| 3 | |||
| 2 | |||
| CC3 | 2 | 9 | 3 |
| 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Centorotola, G.; Guidi, F.; D’Aurizio, G.; Salini, R.; Di Domenico, M.; Ottaviani, D.; Petruzzelli, A.; Fisichella, S.; Duranti, A.; Tonucci, F.; et al. Intensive Environmental Surveillance Plan for Listeria monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity. Foods 2021, 10, 1944. https://doi.org/10.3390/foods10081944
Centorotola G, Guidi F, D’Aurizio G, Salini R, Di Domenico M, Ottaviani D, Petruzzelli A, Fisichella S, Duranti A, Tonucci F, et al. Intensive Environmental Surveillance Plan for Listeria monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity. Foods. 2021; 10(8):1944. https://doi.org/10.3390/foods10081944
Chicago/Turabian StyleCentorotola, Gabriella, Fabrizia Guidi, Guglielmo D’Aurizio, Romolo Salini, Marco Di Domenico, Donatella Ottaviani, Annalisa Petruzzelli, Stefano Fisichella, Anna Duranti, Franco Tonucci, and et al. 2021. "Intensive Environmental Surveillance Plan for Listeria monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity" Foods 10, no. 8: 1944. https://doi.org/10.3390/foods10081944
APA StyleCentorotola, G., Guidi, F., D’Aurizio, G., Salini, R., Di Domenico, M., Ottaviani, D., Petruzzelli, A., Fisichella, S., Duranti, A., Tonucci, F., Acciari, V. A., Torresi, M., Pomilio, F., & Blasi, G. (2021). Intensive Environmental Surveillance Plan for Listeria monocytogenes in Food Producing Plants and Retail Stores of Central Italy: Prevalence and Genetic Diversity. Foods, 10(8), 1944. https://doi.org/10.3390/foods10081944