
Bacterial Community of Grana Padano PDO Cheese and Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling of Cheeses
2.2. Total DNA Extraction
2.3. DNA Metabarcoding
2.4. RAPD-PCR Metafingerprinting
2.5. Data Analysis and Bioinformatics Processing
3. Results
3.1. DNA Metabarcoding Analysis
3.1.1. Species Abundance
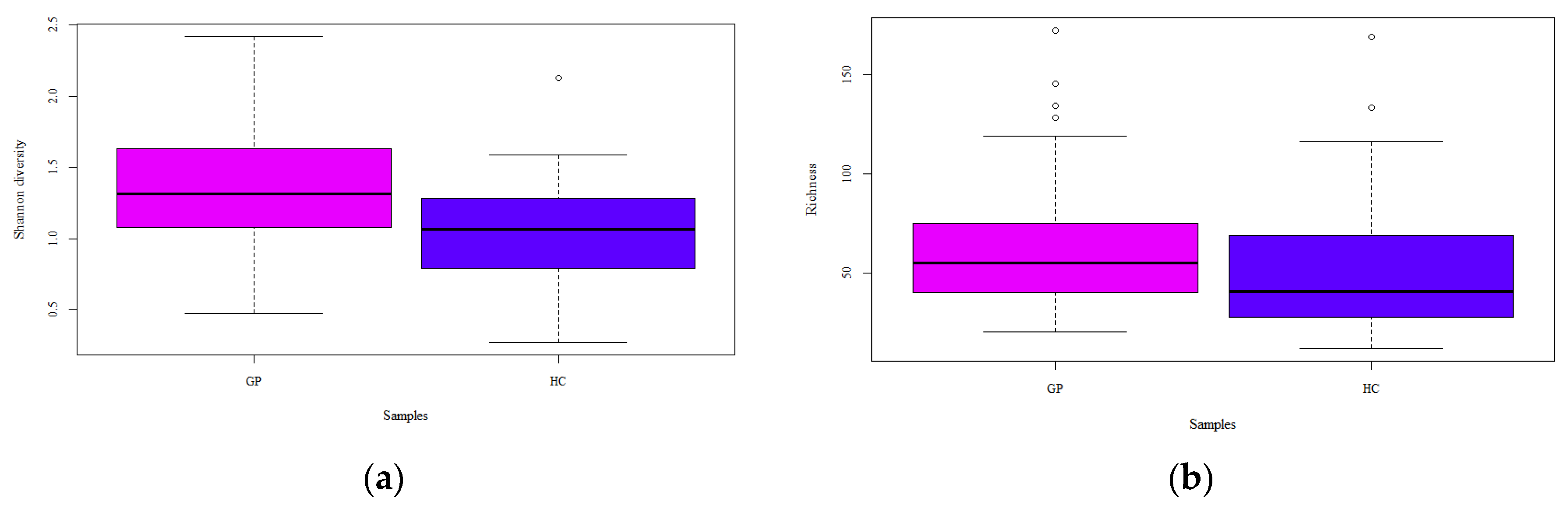
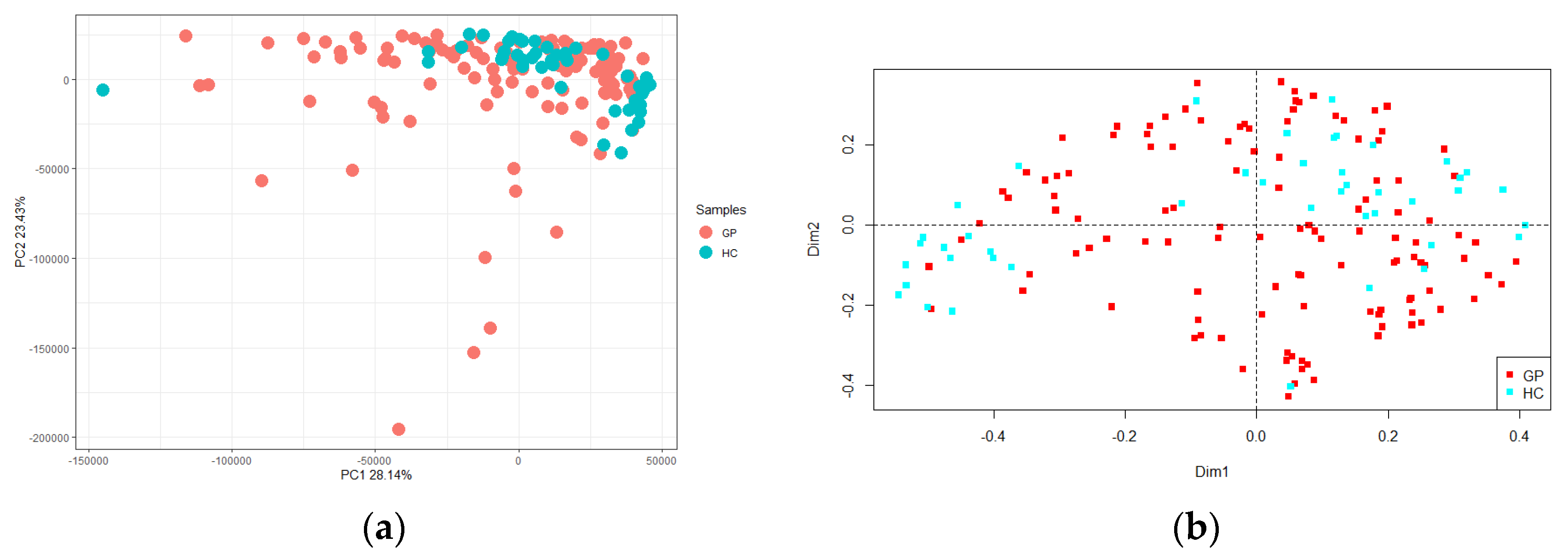
3.1.2. Alpha and Beta Diversity
3.2. RAPD-PCR Metafingerprinting Analysis
3.3. Implementation of a Classifier
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Regulation, E.C. Council Regulation EC no 1151 of November 21, 2012. Off. J. Eur. Union 2015, 343, 1–29. [Google Scholar]
- Gatti, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Mucchetti, G. Invited review: Microbial evolution in raw-milk, long-ripened cheeses produced using undefined natural whey starters. J. Dairy Sci. 2014, 97, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Santarelli, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Gatti, M. Survey on the community and dynamics of lactic acid bacteria in Grana Padano cheese. Syst. Appl. Microbiol. 2013, 36, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernandez-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Delcenserie, V.; Taminiau, B.; Delhalle, L.; Nezer, C.; Doyen, S.; Crevecoeur, S.; Roussey, D.; Korsak, N.; Daube, G. Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 2014, 97, 6046–6056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvetti, T.; Capra, E.; Morandi, S.; Cremonesi, P.; Decimo, M.; Gavazzi, F.; Giannico, R.; De Noni, I.; Brasca, M. Microbial population profile during ripening of Protected Designation of Origin (PDO) Silter cheese, produced with and without autochthonous starter culture. LWT Food Sci. Technol. 2017, 84, 821–831. [Google Scholar] [CrossRef]
- Bertani, G.; Levante, A.; Lazzi, C.; Bottari, B.; Gatti, M.; Neviani, E. Dynamics of a natural bacterial community under technological and environmental pressures: The case of natural whey starter for Parmigiano Reggiano cheese. Food Res. Int. 2020, 129, 108860. [Google Scholar] [CrossRef]
- Penland, M.; Falentin, H.; Parayre, S.; Pawtowski, A.; Maillard, M.-B.; Thierry, A.; Mounier, J.; Coton, M.; Deutsch, S.-M. Linking Pelardon artisanal goat cheese microbial communities to aroma compounds during cheese-making and ripening. Int. J. Food Microbiol. 2021, 345, 109130. [Google Scholar] [CrossRef]
- ICQRF Report 2021. In Department of Central Inspectorate for Fraud Repression and Quality Protection of the Agri-Food Products and Foodstuffs; Ministry of Agricultural, Food and Forestry Policies: Rome, Italy, 2021.
- Bava, L.; Bacenetti, J.; Gislon, G.; Pellegrino, L.; D’Incecco, P.; Sandrucci, A.; Tamburini, A.; Fiala, M.; Zucali, M. Impact assessment of traditional food manufacturing: The case of Grana Padano cheese. Sci Total Environ. 2018, 626, 1200–1209. [Google Scholar] [CrossRef]
- Summer, A.; Formaggioni, P.; Franceschi, P.; Di Frangia, F.; Righi, F.; Malacarne, M. Cheese as functional food: The example of Parmigiano Reggiano and Grana Padano. Food Technol Biotechnol. 2017, 55, 277–289. [Google Scholar] [CrossRef]
- Gobbetti, M.; Fox, P.; Neviani, E. The most traditional and popular Italian cheeses. In The Cheeses of Italy: Science and Technology; Gobbetti, M., Fox, P., Neviani, E., Eds.; Springer Int. Publishing AG: Cham, Switzerland, 2018; pp. 99–267. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Gallo, A.; Masoero, F.; Trevisan, M.; Giuberti, G. Untargeted metabolomics reveals differences in chemical fingerprints between PDO and non-PDO Grana Padano cheeses. Food Res. Int. 2018, 113, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Smit, G.; Smit, B.A.; Engels, W.J.M. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef] [PubMed]
- Barron, L.J.R.; Aldai, N.; Virto, M.; de Renobales, M. Cheeses with Protected land and tradition-related labels: Traceability and authentication. In Global Cheesemaking Technology: Cheese Quality and Characteristics, 1st ed.; Papademas, P., Bintsis, T., Eds.; John Wiley’s and Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 100–119. [Google Scholar] [CrossRef]
- Yeluri Jonnala, B.R.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Sequencing of the cheese microbiome and its relevance to industry. A review. Front Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamimura, B.A.; De Filippis, F.; Sant’Ana, A.S.; Ercolini, D. Large-scale mapping of microbial diversity in artisanal Brazilian cheeses. Food Microbiol. 2019, 80, 40–49. [Google Scholar] [CrossRef]
- Zago, M.; Bardelli, T.; Rossetti, L.; Nazzicari, N.; Carminati, D.; Galli, A.; Giraffa, G. Evaluation of bacterial communities of Grana Padano cheese by DNA metabarcoding and DNA metafingerprinting analysis. Food Microbiol. 2021, 93, 103613. [Google Scholar] [CrossRef] [PubMed]
- Probst, M.; Gómez-Brandón, M.; Bardelli, T.; Egli, M.; Insam, H.; Ascher-Jenull, J. Bacterial communities of decaying Norway spruce follow distinct slope exposure and time-dependent trajectories. Environ Microbiol. 2018, 20, 3657–3670. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package, Version 2.5-5; R Foundation: Vienna, Austria, 2019. [Google Scholar]
- De Mendiburu, F. Package ‘Agricolae.’ R Package Version 1.2-8. Available online: http://CRAN.R-project.org/package=agricolae (accessed on 4 July 2019).
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Meth. 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Reshape2: Flexibly Reshape Data: A Reboot of the Reshape Package; R Package Version; R Foundation: Vienna, Austria, 2012. [Google Scholar]
- Chollet, F. Keras. Available online: https://github.com/fchollet/keras. (accessed on 10 April 2015).
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn Res. 2011, 12, 2825–2830. [Google Scholar]
- Marino, M.; Dubsky de Wittenau, G.; Saccà, E.; Cattonaro, F.; Spadotto, A.; Innocente, N.; Radovic, S.; Piasentier, E.; Marroni, F. Metagenomic profiles of different types of Italian high-moisture Mozzarella cheese. Food Microbiol. 2019, 79, 123–131. [Google Scholar] [CrossRef]
- Pellegrino, L.; Resmini, P. Cheesemaking conditions and compositive characteristics supporting the safety of the raw milk cheese Italian grana. Sci. Tecn. Latt. Cas. 2001, 52, 105–114. [Google Scholar]
- Panari, G.; Perini, S.; Guidetti, R.; Pecorari, M.; Merialdi, G.; Albertini, A. Indagine sul comportamento di germi potenzialmente patogeni nella tecnologia del formaggio Parmigiano-Reggiano. Sci. Tecn. Latt. Cas. 2001, 52, 13–22. [Google Scholar]
- Bachmann, H.P.; Spahr, U. The fate of potentially pathogenic bacteria in Swiss hard and semihard cheeses made from raw milk. J. Dairy Sci. 1995, 78, 476–483. [Google Scholar] [CrossRef]
- Ercolini, D. Secrets of the cheese microbiome. Nat. Foods 2020, 1, 466–467. [Google Scholar] [CrossRef]
- Avila, C.L.S.; Carvalho, B.F. Silage fermentation-updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2019, 128, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Pogačić, T.; Mancini, A.; Santarelli, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Gatti, M. Diversity and dynamic of lactic acid bacteria strains during aging of a long ripened hard cheese produced from raw milk and undefined natural starter. Food Microbiol. 2013, 36, 207–215. [Google Scholar] [CrossRef]
- Bava, L.; Zucali, M.; Tamburini, A.; Morandi, S.; Brasca, M. Effect of different farming practices on lactic acid bacteria content in cow milk. Animals 2021, 11, 522. [Google Scholar] [CrossRef]
- Cattaneo, S.; Hogenboom, J.A.; Masotti, F.; Rosi, V.; Pellegrino, L.; Resmini, P. Grated Grana Padano cheese: New hints on how to control quality and recognize imitations. Dairy Sci. Technol. 2008, 88, 595–605. [Google Scholar] [CrossRef]
- Franklin, R.B.; Taylor, D.R.; Mills, A.L. Characterization of microbial communities using randomly amplified polymorphic DNA (RAPD). J. Microbiol. Meth. 1999, 35, 225–235. [Google Scholar] [CrossRef]
- Khodakova, A.S.; Smith, R.J.; Burgoyne, L.; Abarno, D.; Linacre, A. Random whole metagenomic sequencing for forensic discrimination of soils. PLoS ONE 2014, 9, e104996. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistics | Metabarcoding | Fingerprinting | Fingerprinting Weight Adjusted |
|---|---|---|---|
| FN | 3.952 | 2.188 | 0.840 |
| FP | 8.022 | 0.262 | 2.320 |
| TN | 15.978 | 23.738 | 21.680 |
| TP | 5.048 | 7.812 | 9.160 |
| AUC | 0.657 | 0.934 | 0.972 |
| Loss | 1.147 | 0.338 | 0.312 |
| Precision | 0.386 | 0.968 | 0.798 |
| TPR | 0.561 | 0.781 | 0.916 |
| TNR | 0.666 | 0.989 | 0.903 |
| Accuracy | 0.637 | 0.928 | 0.907 |
| Balanced Accuracy | 0.613 | 0.885 | 0.910 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zago, M.; Rossetti, L.; Bardelli, T.; Carminati, D.; Nazzicari, N.; Giraffa, G. Bacterial Community of Grana Padano PDO Cheese and Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences. Foods 2021, 10, 1826. https://doi.org/10.3390/foods10081826
Zago M, Rossetti L, Bardelli T, Carminati D, Nazzicari N, Giraffa G. Bacterial Community of Grana Padano PDO Cheese and Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences. Foods. 2021; 10(8):1826. https://doi.org/10.3390/foods10081826
Chicago/Turabian StyleZago, Miriam, Lia Rossetti, Tommaso Bardelli, Domenico Carminati, Nelson Nazzicari, and Giorgio Giraffa. 2021. "Bacterial Community of Grana Padano PDO Cheese and Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences" Foods 10, no. 8: 1826. https://doi.org/10.3390/foods10081826
APA StyleZago, M., Rossetti, L., Bardelli, T., Carminati, D., Nazzicari, N., & Giraffa, G. (2021). Bacterial Community of Grana Padano PDO Cheese and Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences. Foods, 10(8), 1826. https://doi.org/10.3390/foods10081826