
Mapping B-Cell Epitopes for Nonspecific Lipid Transfer Proteins of Legumes Consumed in India and Identification of Critical Residues Responsible for IgE Binding


Abstract
1. Introduction
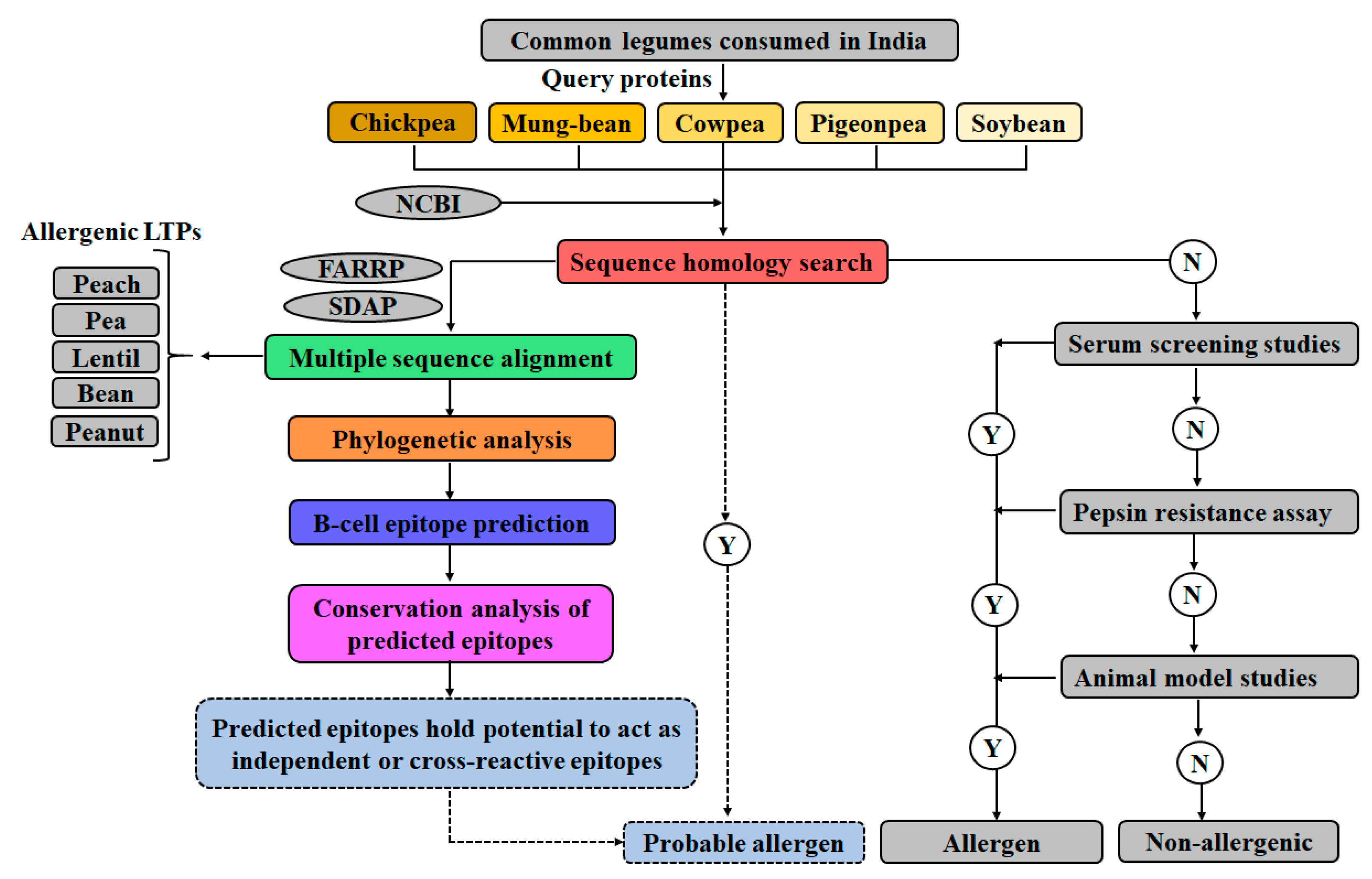
2. Materials and Methods
2.1. Sequence Retrieval
2.2. Sequence Homology Studies
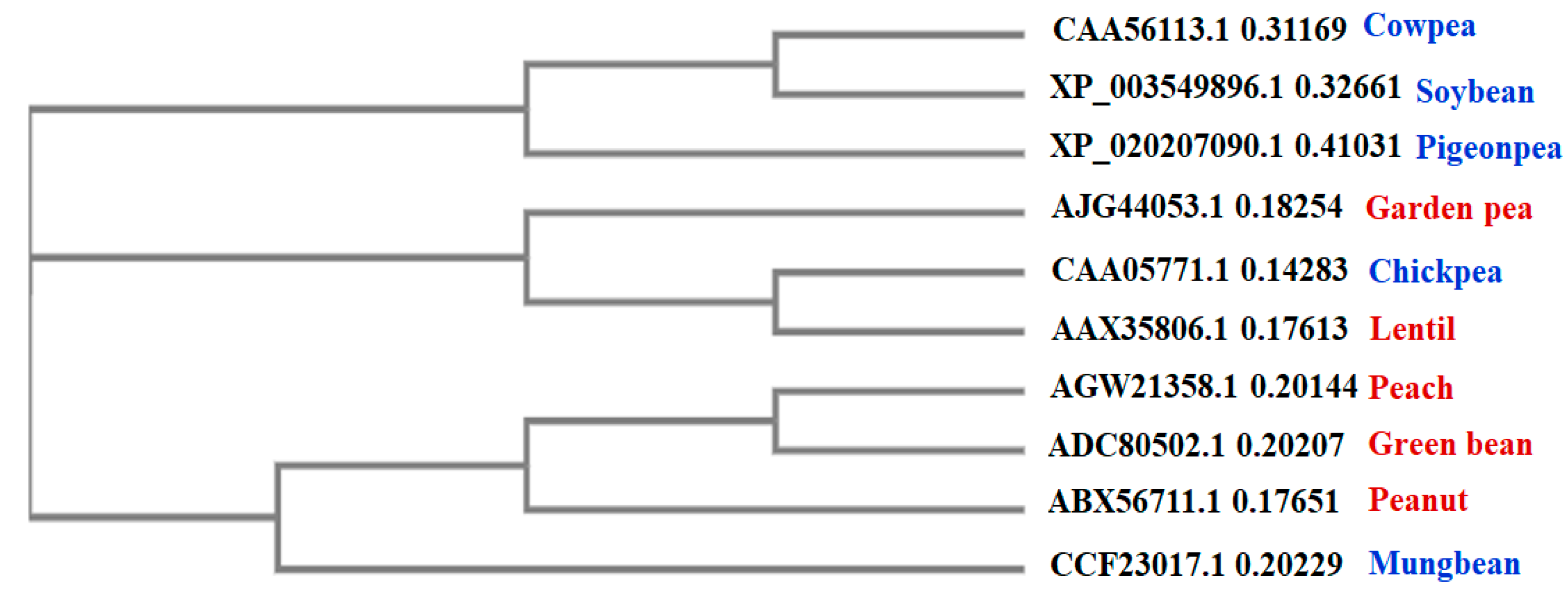
2.3. Multiple Sequence Alignment (MSA) and Phylogenetic Analysis
2.4. Mapping Linear and Conformational IgE Binding Epitopes
2.5. Secondary Structure Prediction
2.6. Conservation Analysis of Legume LTPs Employed in the Study
2.7. Property-Based Peptide Similarity Index (Property Distance Index—PD Values) of the Predicted Linear Epitopes
2.8. Conservation Analysis of the Residues, Experimentally Validated as B-Cell Epitopes of Peach LTP (Pur p 3), among Fifteen Allergenic LTPs
3. Results and Discussion
3.1. Sequence Homology Studies (SDAP and FARRP Databases)
3.2. Multiple Sequence Alignment (MSA) and Phylogenetic Analysis
3.3. Mapping of Consensus Linear and Conformational B-Cell Epitopes
3.3.1. Chickpea LTP
3.3.2. Mung-Bean LTP
3.3.3. Cowpea LTP
3.3.4. Pigeon-Pea LTP
3.3.5. Soybean LTP
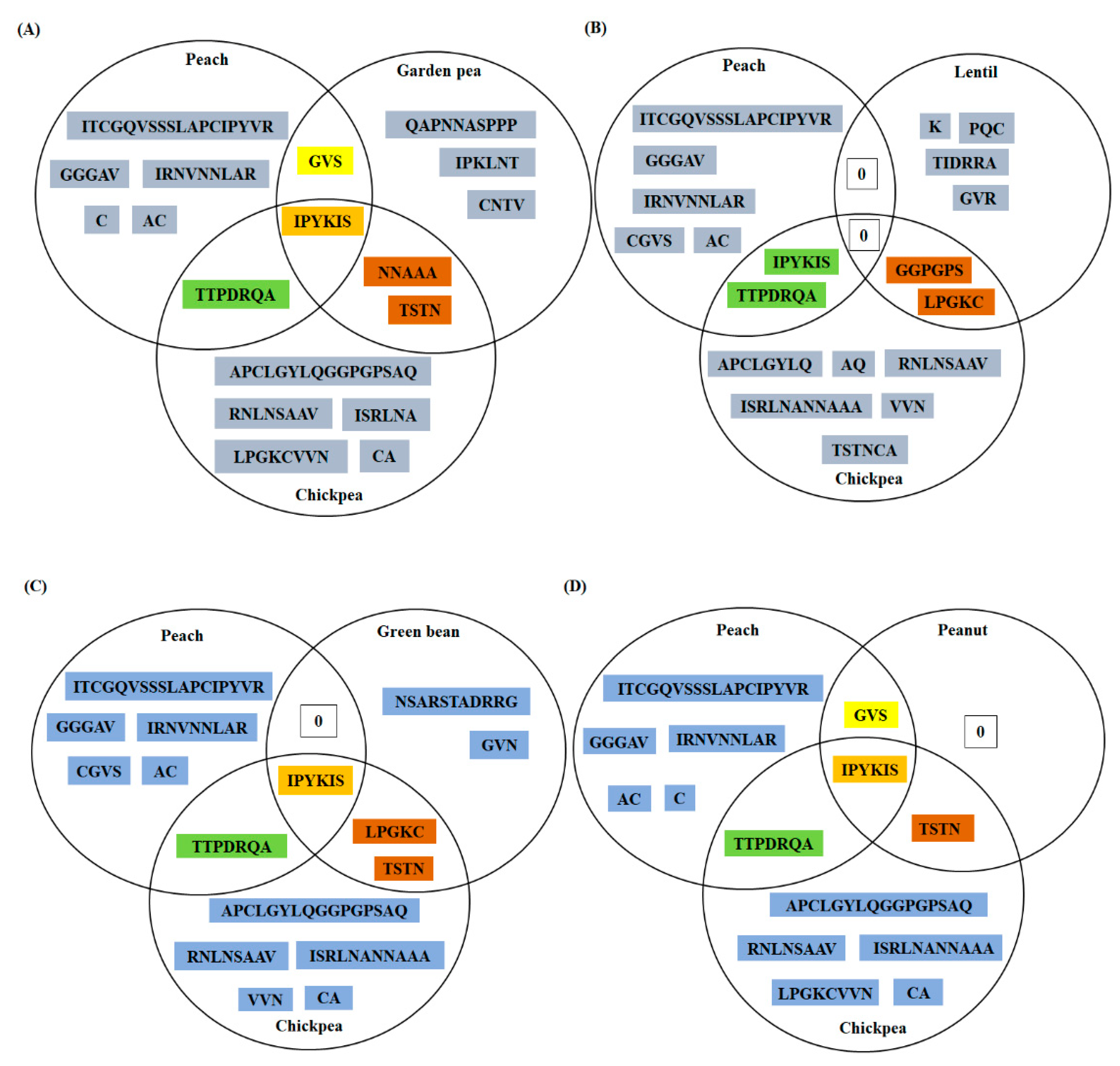
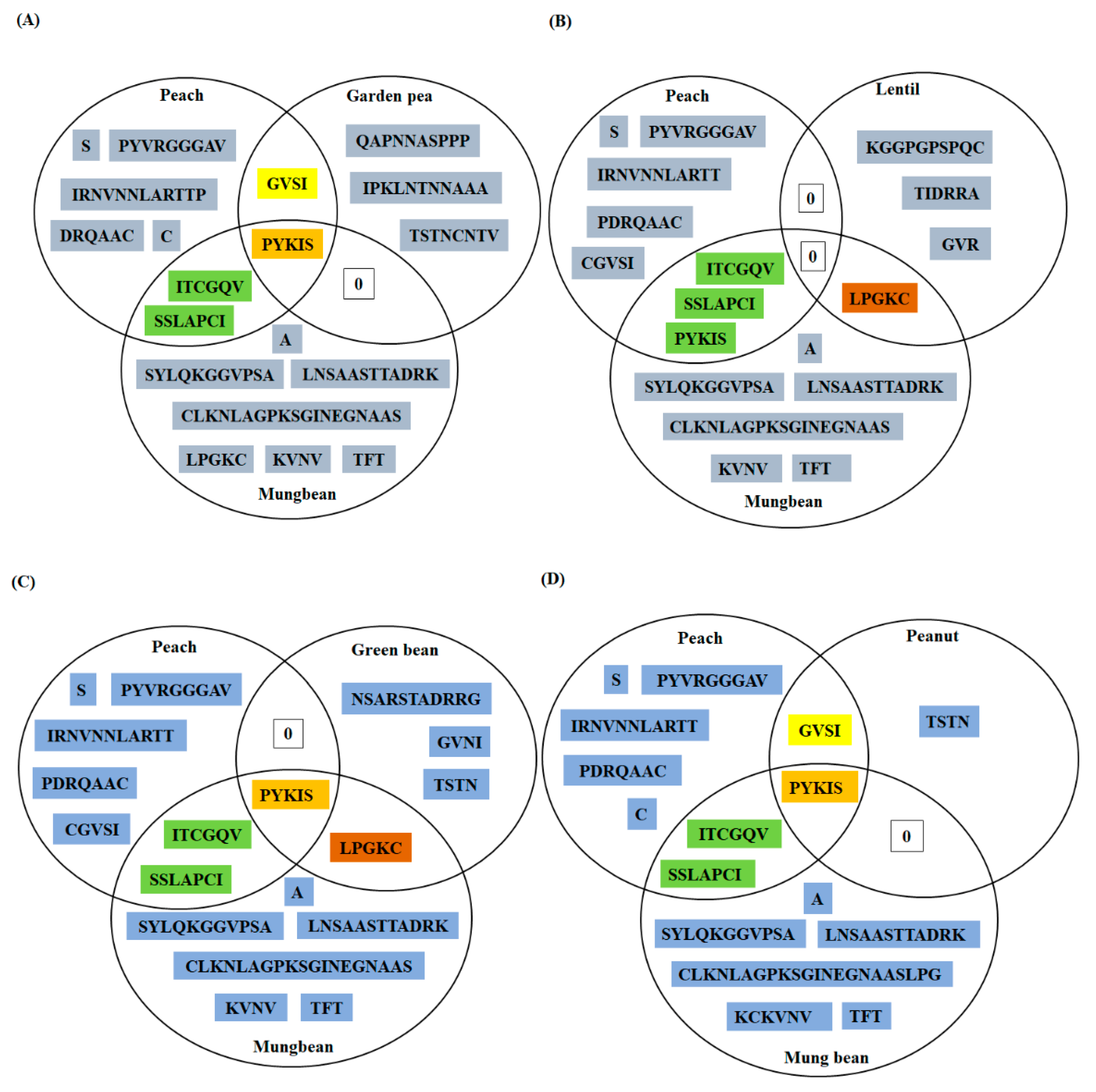
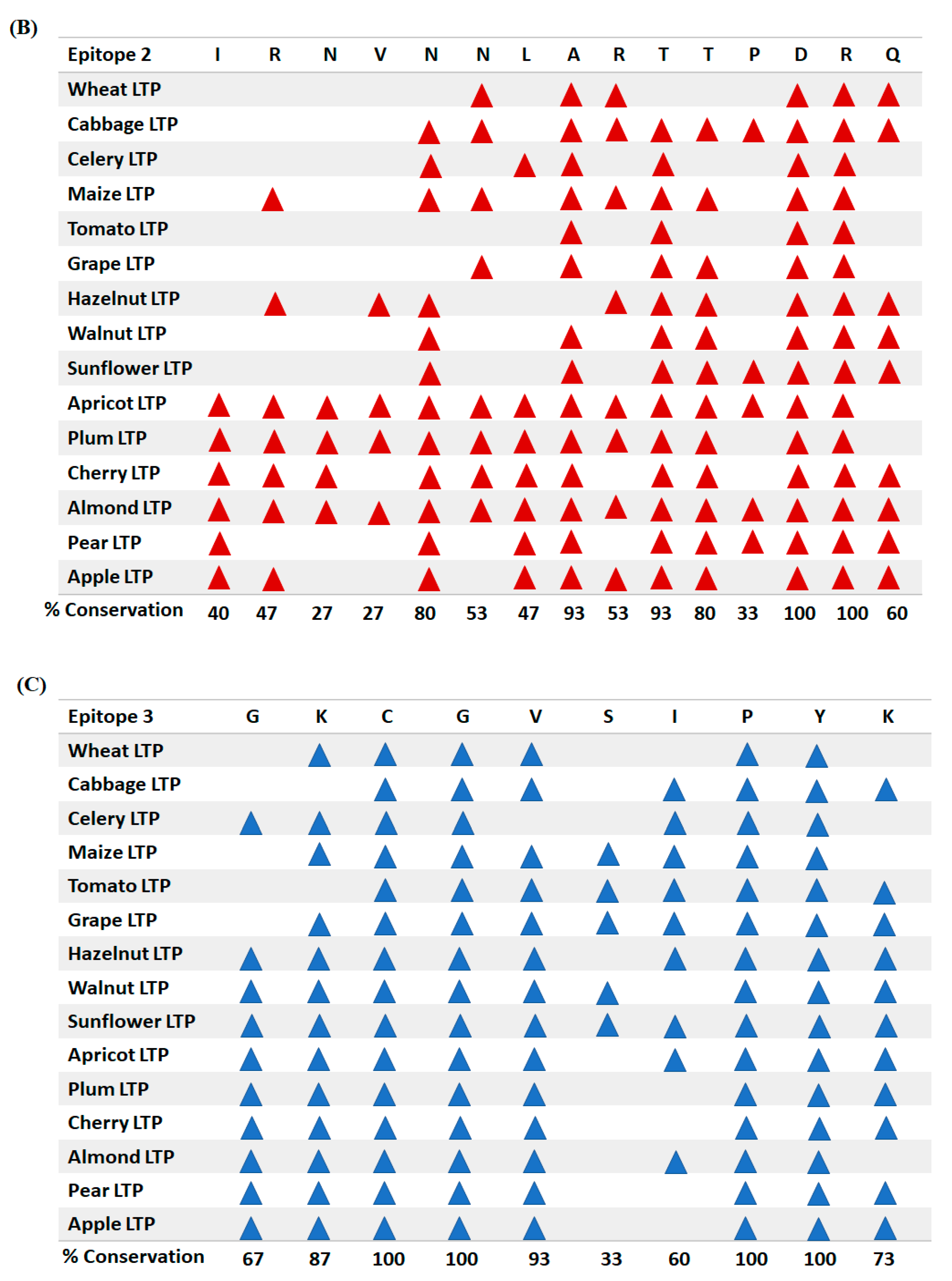
3.4. Conservation Analysis of Individual Amino Acid Residues in Different LTPs
3.5. Peptide Similarity Search of Predicted Linear Epitopes (PD Index)
3.6. Conservation Analysis of the Residues, Experimentally Validated as B-Cell Epitopes of Peach LTP (Pur p 3), among Fifteen Allergenic LTPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Huby, R.D.; Dearman, R.J.; Kimber, I. Why are some proteins allergens? Toxicol. Sci. 2000, 55, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Bannon, G.A. What makes a food protein an allergen? Curr. Allergy Asthma Rep. 2004, 4, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Freeland, D.M.H.; Nadeau, K.C. Food allergy: Immune mechanisms, diagnosis and immunotherapy. Nat. Rev. Immunol. 2016, 16, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; O’Mahony, L.; Burks, A.W.; Plaut, M.; Lack, G.; Akdis, C.A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018, 141, 11–19. [Google Scholar] [CrossRef]
- Looney, T.J.; Lee, J.Y.; Roskin, K.M.; Hoh, R.A.; King, J.; Glanville, J.; Liu, Y.; Pham, T.D.; Dekker, C.L.; Davis, M.M.; et al. Human B-cell isotype switching origins of IgE. J. Allergy Clin. Immunol. 2016, 137, 579–586.e7. [Google Scholar] [CrossRef]
- Gould, H.J.; Wu, Y.B. IgE repertoire and immunological memory: Compartmental regulation and antibody function. Int. Immunol. 2018, 30, 403–412. [Google Scholar] [CrossRef]
- Dougherty, J.A.; Wagner, J.D.; Stanton, M.C. Peanut allergen powder-dnfp: A novel oral immunotherapy to mitigate peanut allergy. Ann. Pharmacother. 2021, 55, 344–353. [Google Scholar] [CrossRef]
- Mishra, A.; Gaur, S.N.; Lavasa, S.; Arora, N. In vitro assessment of allergenicity features and localization of probable IgE binding regions. Food Chem. Toxicol. 2015, 84, 181–187. [Google Scholar] [CrossRef]
- Mishra, A.; Jain, A.; Arora, N. Mapping B-cell epitopes of major and minor peanut allergens and identifying residues contributing to IgE binding. J. Sci. Food Agric. 2016, 96, 539–547. [Google Scholar] [CrossRef]
- Barre, A.; Jacquet, G.; Sordet, C.; Culerrier, R.; Rougé, P. Homology modelling and conformational analysis of IgE-binding epitopes of Ara h 3 and other legumin allergens with a cupin fold from tree nuts. Mol. Immunol. 2007, 44, 3243–3255. [Google Scholar] [CrossRef]
- Pomés, A. Relevant B cell epitopes in allergic disease. Int. Arch. Allergy Immunol. 2010, 152, 1–11. [Google Scholar] [CrossRef]
- Teodorowicz, M.; van Neerven, J.; Savelkoul, H. Food processing: The influence of the maillard reaction on immunogenicity and allergenicity of food proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- McClain, S. Bioinformatic screening and detection of allergen cross-reactive IgE-binding epitopes. Mol. Nutr. Food Res. 2017, 61, 1600676. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.R.; Scheurer, S.; Vieths, S. Food allergens: Molecular and immunological aspects, allergen databases and cross-reactivity. Chem. Immunol. Allergy 2015, 101, 18–29. [Google Scholar] [PubMed]
- Jimenez-Lopez, J.C.; Gachomo, E.W.; Ariyo, O.A.; Baba-Moussa, L.; Kotchoni, S.O. Specific conformational epitope features of pathogenesis-related proteins mediating cross-reactivity between pollen and food allergens. Mol. Biol. Rep. 2012, 39, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Vieths, S.; Scheurer, S.; Ballmer-Weber, B. Current understanding of cross-reactivity of food allergens and pollen. Ann. N. Y. Acad. Sci. 2002, 964, 47–68. [Google Scholar] [CrossRef]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S.; Caldironi, G.; Barocci, F.; van Ree, R. Immunological cross-reactivity between lipid transfer proteins from botanically unrelated plant-derived foods: A clinical study. Allergy 2002, 57, 900–906. [Google Scholar] [CrossRef]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: Classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef] [PubMed]
- Pekar, J.; Ret, D.; Untersmayr, E. Stability of allergens. Mol. Immunol. 2018, 100, 14–20. [Google Scholar] [CrossRef]
- Wal, J.M. Thermal processing and allergenicity of foods. Allergy 2003, 58, 727–729. [Google Scholar] [CrossRef]
- Bhattacharya, K.; Sircar, G.; Dasgupta, A.; Gupta Bhattacharya, S. Spectrum of allergens and allergen biology in India. Int. Arch. Allergy Immunol. 2018, 177, 219–237. [Google Scholar] [CrossRef]
- Kumari, D.; Arora, N.; Kasera, R.; Sridhara, S.; Kumar, R.; Singh, B.P. Isolation and characterization of a 28 kDa major allergen from blackgram (Phaseolus mungo). Immunobiology 2012, 217, 895–904. [Google Scholar] [CrossRef]
- Breiteneder, H.; Radauer, C. A classification of plant food allergens. J. Allergy Clin. Immunol. 2004, 113, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Asero, R.; Mistrello, G.; Roncarolo, D.; de Vries, S.C.; Gautier, M.F.; Ciurana, C.L.; Verbeek, E.; Mohammadi, T.; Knul-Brettlova, V.; Akkerdaas, J.H.; et al. Lipid transfer protein: A pan-allergen in plant-derived foods that is highly resistant to pepsin digestion. Int. Arch. Allergy Immunol. 2001, 124, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S. Relationship between peach lipid transfer protein specific IgE levels and hypersensitivity to non-Rosaceae vegetable foods in patients allergic to lipid transfer protein. Ann. Allergy Asthma Immunol. 2004, 92, 268–272. [Google Scholar] [CrossRef]
- Mishra, A.; Arora, N. Allergenicity assessment of transgenic wheat lines in silico. Methods Mol. Biol. 2017, 1679, 97–111. [Google Scholar]
- Mishra, A.; Gaur, S.N.; Singh, B.P.; Arora, N. In silico assessment of the potential allergenicity of transgenes used for the development of GM food crops. Food Chem. Toxicol. 2012, 50, 1334–1339. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. BcePred: Prediction of continuous B-cell epitopes in antigenic sequences using physico-chemical properties. In Artificial Immune Systems, ICARIS 2004; Nicosia, G., Cutello, V., Bentley, P.J., Timis, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 197–204. [Google Scholar]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Ansari, H.R.; Raghava, G.P. Improved method for linear B-cell epitope prediction using antigen’s primary sequence. PLoS ONE 2013, 8, e62216. [Google Scholar] [CrossRef]
- Sweredoski, M.J.; Baldi, P. COBEpro: A novel system for predicting continuous B-cell epitopes. Protein Eng. Des. Sel. 2009, 22, 113–120. [Google Scholar] [CrossRef]
- Yao, B.; Zhang, L.; Liang, S.; Zhang, C. SVMTriP: A method to predict antigenic epitopes using support vector machine to integrate tri-peptide similarity and propensity. PLoS ONE 2012, 7, e45152. [Google Scholar] [CrossRef]
- Manavalan, B.; Govindaraj, R.G.; Shin, T.H.; Kim, M.O.; Lee, G. iBCE-EL: A new ensemble learning framework for improved linear B-cell epitope prediction. Front. Immunol. 2018, 9, 1695. [Google Scholar] [CrossRef]
- Ansari, H.R.; Raghava, G.P. Identification of conformational B-cell Epitopes in an antigen from its primary sequence. Immunome Res. 2010, 6, 1–9. [Google Scholar] [CrossRef]
- Sela-Culang, I.; Benhnia, M.R.; Matho, M.H.; Kaever, T.; Maybeno, M.; Schlossman, A.; Nimrod, G.; Li, S.; Xiang, Y.; Zajonc, D.; et al. Using a combined computational-experimental approach to predict antibody-specific B cell epitopes. Structure 2014, 22, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Ivanciuc, O.; Schein, C.H.; Braun, W. SDAP: Database and computational tools for allergenic proteins. Nucleic Acids Res. 2003, 31, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.E.; Ebisawa, M.; Ferreira, F.; Sampson, H.A.; van Ree, R.; Vieths, S.; Baumert, J.L.; Bohle, B.; Lalithambika, S.; Wise, J.; et al. AllergenOnline: A peer-reviewed, curated allergen database to assess novel food proteins for potential cross-reactivity. Mol. Nutr. Food Res. 2016, 60, 1183–1198. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization/World Health Organization. Evaluation of the Allergenicity of Genetically Modified foods: Report of a Joint FAO/WHO Expert Consultation on Allergenicity of Foods Derived from Biotechnology, 22–25 January 2001; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2001. [Google Scholar]
- Codex Alimentarius Commission. Alinorm 03/34: Joint FAO/WHO Food Standard Programme, Codex Alimentarius Commission, Appendix III, Guideline for the Conduct of Food Safety Assessment of Foods Derived from Recombinant DNA Plants and Appendix IV, Annex on the Assessment of Possible Allergenicity, 25th Session, Rome, Italy, 30 June–5 July 2003; Codex Alimentarius Commission: Rome, Italy, 5 July 2003; pp. 47–60. [Google Scholar]
- Goodman, R.E.; Vieths, S.; Sampson, H.A.; Hill, D.; Ebisawa, M.; Taylor, S.L.; van Ree, R. Allergenicity assessment of genetically modified crops—What makes sense? Nat. Biotechnol. 2008, 26, 73–81. [Google Scholar] [CrossRef]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S. Detection of some safe plant-derived foods for LTP-allergic patients. Int. Arch. Allergy Immunol. 2007, 144, 57–63. [Google Scholar] [CrossRef] [PubMed]
- García-Casado, G.; Pacios, L.F.; Díaz-Perales, A.; Sánchez-Monge, R.; Lombardero, M.; García-Selles, F.J.; Polo, F.; Barber, D.; Salcedo, G. Identification of IgE-binding epitopes of the major peach allergen Pru p 3. J. Allergy Clin. Immunol. 2003, 112, 599–605. [Google Scholar] [CrossRef]
- Eichhorn, S.; Steiner, M.; Laimer, J.; Lackner, P.; Zuidmeer-Jongejan, L.; Briza, P.; Mari, A.; van Ree, R.; Ferreira, F.; Gadermaier, G. Development of a hypoallergenic and immunogenic Pru p 3 proline variant for treatment of peach allergy. Clin. Transl. Allergy 2014, 4 (Suppl. 2), O22. [Google Scholar] [CrossRef]
- Skypala, I.J.; Cecchi, L.; Shamji, M.H.; Scala, E.; Till, S. Lipid transfer protein allergy in the United Kingdom: Characterization and comparison with a matched Italian cohort. Allergy 2019, 74, 1340–1351. [Google Scholar] [CrossRef]
- Bogdanov, I.V.; Shenkarev, Z.O.; Finkina, E.I.; Melnikova, D.N.; Rumynskiy, E.I.; Arseniev, A.S.; Ovchinnikova, T.V. A novel lipid transfer protein from the pea Pisum sativum: Isolation, recombinant expression, solution structure, antifungal activity, lipid binding, and allergenic properties. BMC Plant Biol. 2016, 16, 107. [Google Scholar] [CrossRef] [PubMed]
- Bastiaan-Net, S.; Reitsma, M.; Cordewener, J.; van der Valk, J.; America, T.; Dubois, A.; Gerth van Wijk, R.; Savelkoul, H.; de Jong, N.W.; Wichers, H.J. IgE cross-reactivity of cashew nut allergens. Int. Arch. Allergy Immunol. 2019, 178, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Asero, R.; Piantanida, M.; Pinter, E.; Pravettoni, V. The clinical relevance of lipid transfer protein. Clin. Exp. Allergy 2018, 48, 6–12. [Google Scholar] [CrossRef]
- Kulkarni, A.; Ananthanarayan, L.; Raman, K. Identification of putative and potential cross-reactive chickpea (Cicer arietinum) allergens through an in silico approach. Comput. Biol. Chem. 2013, 47, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Garino, C.; Coïsson, J.D.; Arlorio, M. In silico allergenicity prediction of several lipid transfer proteins. Comput. Biol. Chem. 2016, 60, 32–42. [Google Scholar] [CrossRef]
- Bernardi, M.L.; Giangrieco, I.; Camardella, L.; Ferrara, R.; Palazzo, P.; Panico, M.R.; Crescenzo, R.; Carratore, V.; Zennaro, D.; Liso, M.; et al. Allergenic lipid transfer proteins from plant-derived foods do not immunologically and clinically behave homogeneously: The kiwifruit LTP as a model. PLoS ONE 2011, 6, e27856. [Google Scholar] [CrossRef]
- Palacin, A.; Quirce, S.; Armentia, A.; Fernández-Nieto, M.; Pacios, L.F.; Asensio, T.; Sastre, J.; Diaz-Perales, A.; Salcedo, G. Wheat lipid transfer protein is a major allergen associated with baker’s asthma. J. Allergy Clin. Immunol. 2007, 120, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Kapingidza, A.B.; Dolamore, C.; Kowal, K. A robust method for the estimation and visualization of IgE cross-reactivity likelihood between allergens belonging to the same protein family. PLoS ONE 2018, 13, e0208276. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Homology Studies for Chickpea LTP Using FARRP Database | |||
|---|---|---|---|
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Lens culinaris | 71.80% | 81.30% |
| 2 | Pisum sativum | 70.60% | 76.20% |
| 3 | Phaseolus vulgaris | 62.70% | 72.80% |
| 4 | Malus x domestica | 60.30% | 70.40% |
| 5 | Arachis hypogaea | 67.00% | 70.00% |
| 6 | Prunus armeniaca | 57.40% | 68.80% |
| 7 | Hevea brasiliensis | 59.80% | 66.70% |
| Sequence Homology Studies for Chickpea LTP Using SDAP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Lens culinaris | 71.55% | 81.25% |
| 2 | Phaseolus vulgaris | 63.79% | -- |
| 3 | Hevea brasiliensis | 60.34% | -- |
| 4 | Arachis hypogaea | 62.07% | -- |
| 5 | Malus x domestica | 59.48% | -- |
| Sequence Homology Studies for Mungbean LTP Using FARRP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Malus x domestica | 62.10% | 68.80% |
| 2 | Arachis hypogaea | 67.00% | 67.55% |
| 3 | Phaseolus vulgaris | 62.40% | 63.70% |
| 4 | Fragaria x ananassa | 57.90% | 62.50% |
| 5 | Lens culinaris | 55.80% | 61.30% |
| Sequence Homology Studies for Mungbean LTP Using SDAP Database | |||
| S. no. | Allergen source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Phaseolus vulgaris | 62.93% | 62.50% |
| 2 | Malus x domestica | 61.21% | 67.50% |
| 3 | Arachis hypogaea | 62.07% | 67.50% |
| 4 | Lens culinaris | 54.31% | -- |
| Sequence Homology Studies for Cowpea LTP Using FARRP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Lens culinaris | 29.9% | -- |
| 2 | Solanum lycopersicum | 50% | 50% |
| Sequence Homology Studies for Cowpea LTP Using SDAP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Lens culinaris | 32.32% | 33.75% |
| Sequence Homology Studies for Pigeon-pea LTP Using FARRP Database | |||
| S. no. | Allergen source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Zea mays | 27% | -- |
| 2 | Triticum aestivum | 29.8% | -- |
| Sequence Homology Studies for Pigeon-pea LTP Using SDAP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Lens culinaris | -- | 31.25% |
| Sequence Homology Studies for Soybean LTP Using FARRP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Solanum lycopersicum | 32.70% | 35% |
| Sequence Homology Studies for Soybean LTP Using SDAP Database | |||
| S. no. | Allergen Source | Full Length FASTA Alignment | 80 Amino Acid Sliding Window |
| Identity Percentage | Identity Percentage | ||
| 1 | Arachis hypogaea | 23.18% | 28.75% |
| 2 | Hevea brasiliensis | -- | 36.25% |
| LTP Source | No. of Epitopes | Position | Predicted Epitope Sequence | Prediction Servers |
|---|---|---|---|---|
| Chickpea | 1 | 35–50 | APCLGYLQGGPGPSAQ | ABCpred BCEPred BepiPred 2.0 LBtope COBEpro SVMTriP iBCE-EL CBTOPE |
| 2 | 56–70 | RNLNSAAVTTPDRQA | ||
| 3 | 82–112 | ISRLNANNAAALPGKCVVNIPYKISTSTNCA | ||
| Mung-bean | No. of epitopes | Position | Predicted epitope sequence | |
| 1 | 26–50 | ITCGQVASSLAPCISYLQKGGVPSA | ||
| 2 | 59–70 | LNSAASTTADRK | ||
| 3 | 75–110 | CLKNLAGPKSGINEGNAASLPGKCKVNVPYKISTFT | ||
| Cowpea | No. of epitopes | Position | Predicted epitope sequence | |
| 1 | 29–35 | AEAVTCN | ||
| 2 | 37–50 | TELSSCVPAITGGS | ||
| 3 | 53–65 | SSTCCSKLKVQEP | ||
| 4 | 72–81 | KNPSLKQYVN | ||
| 5 | 93–99 | GVTYPNC | ||
| Pigeon-pea | No. of epitopes | Position | Predicted epitope sequence | |
| 1 | 25–39 | ASDIPATCNGDEPVL | ||
| 2 | 47–55 | VNKVPNPSS | ||
| 3 | 69–79 | MGDNTGQGIRD | ||
| 4 | 124–131 | LSNQEKNY | ||
| Soybean | No. of epitopes | Position | Predicted epitope sequence | |
| 1 | 3–10 | MGGGCKCL | ||
| 2 | 21–33 | RSLAEAQSGSSTT | ||
| 3 | 46–51 | NGTTTP | ||
| 4 | 59–66 | LKQTVENQ | ||
| 5 | 108–120 | NGSAPAPGSGPPP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, A.; Kumar, A. Mapping B-Cell Epitopes for Nonspecific Lipid Transfer Proteins of Legumes Consumed in India and Identification of Critical Residues Responsible for IgE Binding. Foods 2021, 10, 1269. https://doi.org/10.3390/foods10061269
Mishra A, Kumar A. Mapping B-Cell Epitopes for Nonspecific Lipid Transfer Proteins of Legumes Consumed in India and Identification of Critical Residues Responsible for IgE Binding. Foods. 2021; 10(6):1269. https://doi.org/10.3390/foods10061269
Chicago/Turabian StyleMishra, Ankita, and Ashok Kumar. 2021. "Mapping B-Cell Epitopes for Nonspecific Lipid Transfer Proteins of Legumes Consumed in India and Identification of Critical Residues Responsible for IgE Binding" Foods 10, no. 6: 1269. https://doi.org/10.3390/foods10061269
APA StyleMishra, A., & Kumar, A. (2021). Mapping B-Cell Epitopes for Nonspecific Lipid Transfer Proteins of Legumes Consumed in India and Identification of Critical Residues Responsible for IgE Binding. Foods, 10(6), 1269. https://doi.org/10.3390/foods10061269