
The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Pepsin Activity
2.3. Trypsin Activity
2.4. Amylase Activity
2.5. Lipase Activity
2.6. In Vitro Digestion
2.7. HR-NMR Spectroscopy
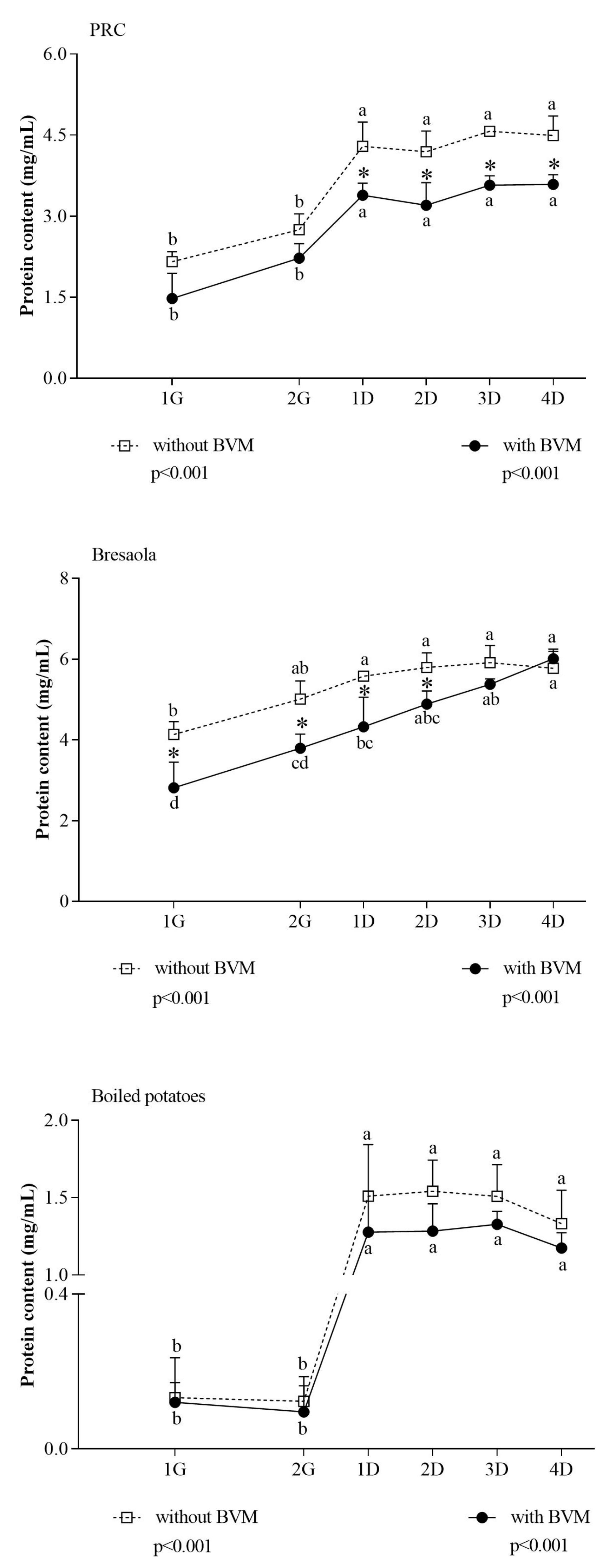
2.8. Protein Quantitative Analysis
2.9. Statistical Analysis
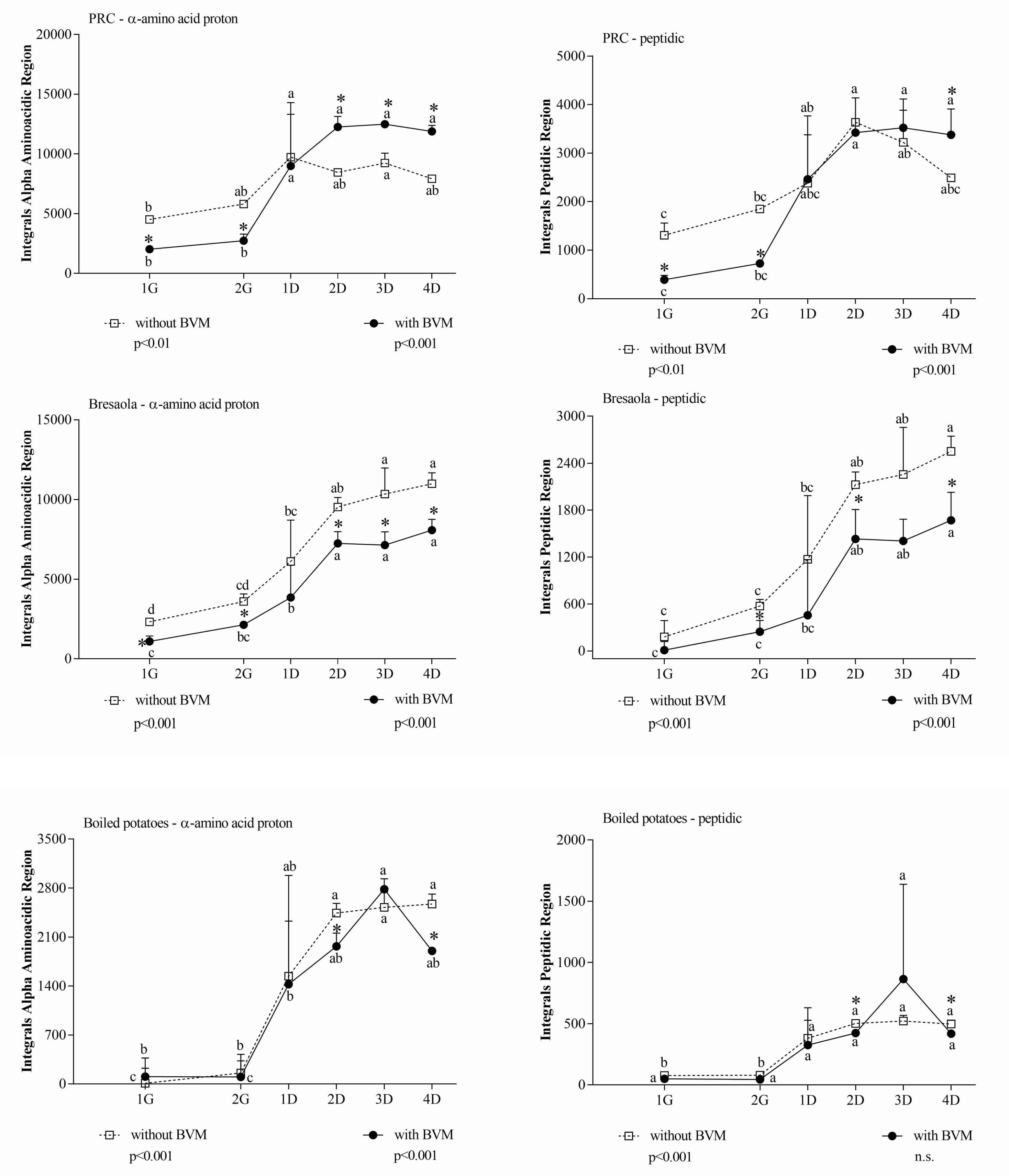
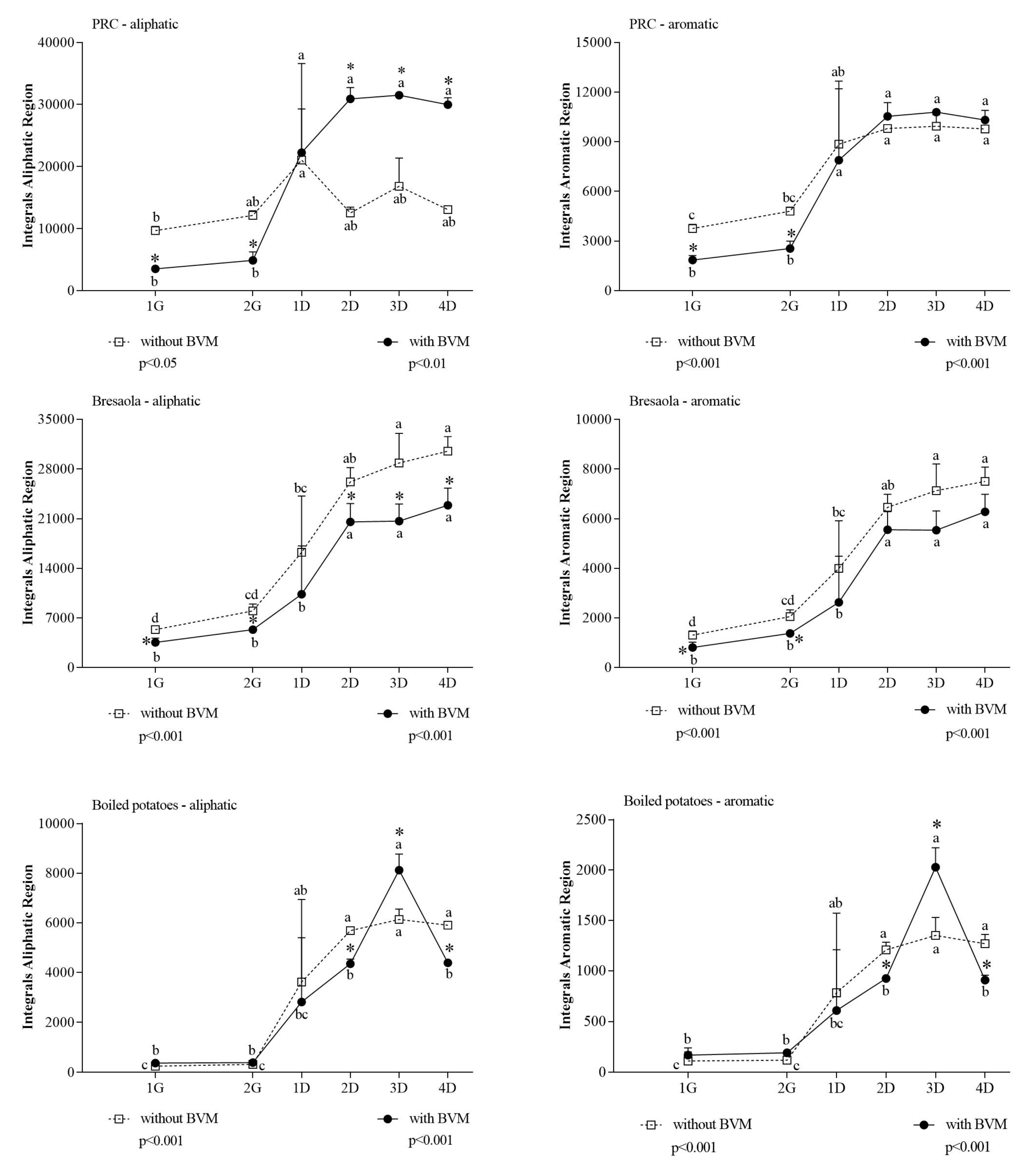
3. Results and Discussion
- (i)
- The aromatic region, where the signals of tyrosine, phenylalanine, and tryptophan resonate;
- (ii)
- The aliphatic region, where the signals of all the branched chain amino acids (BCAAs) fall;
- (iii)
- The α-amino acid proton region, which includes the signals of all the amino acids (AAs), including those mentioned above, both in the free state and bound to peptides or in soluble proteins;
- (iv)
- The peptide proton region, collecting the amide signals that are not accessible to water. They are part of structured proteins or macropeptides that hide the peptide bonds inside the globular structure. Therefore, small peptides, which necessarily expose this group, and single AAs, which do not have the peptide bond, are not providing signals in this region.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenzer, H.; Büsser, S.; Sadeghi, L. Essentials of Bioaccessibility and Bioavailability for Applied Nutritional Practices. J. Mol. Biomark. Diagn. 2016, 7, 307–309. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Dima, S.; Jafari, S.M. Bioavailability and bioaccessibility of food bioactive compounds; overview and assessment by in vitro methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2862–2884. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.; Ramos, R.; Luís, Â.; Rocha, S.; Rosado, T.; Gallardo, E.; Duarte, A.P. Assessment of the Bioaccessibility and Bioavailability of the Phenolic Compounds of Prunus avium L. by in Vitro Digestion and Cell Model. ACS Omega 2019, 4, 7605–7613. [Google Scholar] [CrossRef]
- Fardet, A.; Dupont, D.; Rioux, L.E.; Turgeon, S.L. Influence of food structure on dairy protein, lipid and calcium bioavailability: A narrative review of evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 1987–2010. [Google Scholar] [CrossRef] [PubMed]
- Hiolle, M.; Lechevalier, V.; Floury, J.; Boulier-Monthéan, N.; Prioul, C.; Dupont, D.; Nau, F. In vitro digestion of complex foods: How microstructure influences food disintegration and micronutrient bioaccessibility. Food Res. Int. 2020, 128, 108817. [Google Scholar] [CrossRef] [PubMed]
- Marcolini, E.; Babini, E.; Bordoni, A.; Di Nunzio, M.; Laghi, L.; Maczó, A.; Picone, G.; Szerdahelyi, E.; Valli, V.; Capozzi, F. Bioaccessibility of the Bioactive Peptide Carnosine during in Vitro Digestion of Cured Beef Meat. J. Agric. Food Chem. 2015, 63, 4973–4978. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Busconi, M.; Marocco, A.; Trevisan, M.; Lucini, L. Pigmented sorghum polyphenols as potential inhibitors of starch digestibility: An in vitro study combining starch digestion and untargeted metabolomics. Food Chem. 2020, 312, 126077. [Google Scholar] [CrossRef]
- Ballot, D.; Baynes, R.D.; Bothwell, T.H.; Gillooly, M.; MacFarlane, B.J.; MacPhail, A.P.; Lyons, G.; Derman, D.P.; Bezwoda, W.R.; Torrance, J.D.; et al. The effects of fruit juices and fruits on the absorption of iron from a rice meal. Br. J. Nutr. 1987, 57, 331–343. [Google Scholar] [CrossRef]
- Bothwell, T.H.; Baynes, R.D.; MacFarlane, B.J.; MacPhail, A.P. Nutritional iron requirements and food iron absorption. J. Internal Med. 1989, 226, 357–365. [Google Scholar] [CrossRef]
- Bugianesi, R.; Salucci, M.; Leonardi, C.; Ferracane, R.; Catasta, G.; Azzini, E.; Maiani, G. Effect of domestic cooking on human bioavailability of naringenin, chlorogenic acid, lycopene and β-carotene in cherry tomatoes. Eur. J. Nutr. 2004, 43, 360–366. [Google Scholar] [CrossRef]
- Fielding, J.M.; Rowley, K.G.; Cooper, P.; O’Dea, K. Increases in plasma lycopene concentration after consumption of tomatoes cooked with olive oil. Asia Pac. J. Clin. Nutr. 2005, 14, 131–136. [Google Scholar]
- Budak, N.H.; Aykin, E.; Seydim, A.C.; Greene, A.K.; Guzel-Seydim, Z.B. Functional properties of vinegar. J. Food Sci. 2014, 79, R757–R764. [Google Scholar] [CrossRef]
- Ho, C.W.; Lazim, A.M.; Fazry, S.; Zaki, U.; Lim, S.J. Varieties, production, composition and health benefits of vinegars: A review. Food Chem. 2017, 221, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Steplewska, I.; Long, C.A.; Harris, L.N.; Ryals, R.H. Examination of the antiglycemic properties of vinegar in healthy adults. Ann. Nutr. Metab. 2010, 56, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Seok, H.; Lee, J.Y.; Park, E.M.; Park, S.E.; Lee, J.H.; Lim, S.; Lee, B.W.; Kang, E.S.; Lee, H.C.; Cha, B.S. Balsamic Vinegar Improves High Fat-Induced Beta Cell Dysfunction via Beta Cell ABCA1. Diabetes Metab. J. 2012, 36, 275–279. [Google Scholar] [CrossRef]
- Zhao, Y.; He, Z.; Hao, W.; Zhu, H.; Liang, N.; Liu, J.; Zhang, C.; Ma, K.Y.; He, W.S.; Yang, Y.; et al. Vinegars but not acetic acid are effective in reducing plasma cholesterol in hamsters fed a high-cholesterol diet. Food Funct. 2020, 11, 2163–2172. [Google Scholar] [CrossRef]
- Petsiou, E.I.; Mitrou, P.I.; Raptis, S.A.; Dimitriadis, G.D. Effect and mechanisms of action of vinegar on glucose metabolism, lipid profile, and body weight. Nutr. Rev. 2014, 72, 651–661. [Google Scholar] [CrossRef]
- Ahmadniaye Motlagh, H.; Javadmanesh, A.; Safari, O. Improvement of non-specific immunity, growth, and activity of digestive enzymes in Carassius auratus as a result of apple cider vinegar administration to diet. Fish. Physiol. Biochem. 2020, 46, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.; Le Feunteun, S. Acid induced reduction of the glycaemic response to starch-rich foods: The salivary α-amylase inhibition hypothesis. Food Funct. 2018, 9, 5096–5102. [Google Scholar] [CrossRef]
- Deng, R.; Janssen, A.E.M.; Vergeldt, F.J.; Van As, H.; de Graaf, C.; Mars, M.; Smeets, P.A.M. Exploring in vitro gastric digestion of whey protein by time-domain nuclear magnetic resonance and magnetic resonance imaging. Food Hydrocoll. 2020, 99, 105348. [Google Scholar] [CrossRef]
- Ferranti, P.; Nitride, C.; Nicolai, M.A.; Mamone, G.; Picariello, G.; Bordoni, A.; Valli, V.; Di Nunzio, M.; Babini, E.; Marcolini, E.; et al. In vitro digestion of Bresaola proteins and release of potential bioactive peptides. Food Res. Int. 2014, 63, 157–169. [Google Scholar] [CrossRef]
- Marze, S. Bioavailability of Nutrients and Micronutrients: Advances in Modeling and In Vitro Approaches. Annu. Rev. Food Sci. Technol. 2017, 8, 35–55. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. A method based on 1H NMR spectral data useful to evaluate the hydrolysis level in complex lipid mixtures. Food Res. Int. 2014, 66, 379–387. [Google Scholar] [CrossRef]
- Nieva-Echevarria, B.; Goicoechea, E.; Manzanos, M.J.; Guillen, M.D. Usefulness of (1)H NMR in assessing the extent of lipid digestion. Food Chem. 2015, 179, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Nieva-Echevarria, B.; Goicoechea, E.; Manzanos, M.J.; Guillen, M.D. (1)H NMR and SPME-GC/MS study of hydrolysis, oxidation and other reactions occurring during in vitro digestion of non-oxidized and oxidized sunflower oil. Formation of hydroxy-octadecadienoates. Food Res. Int. 2017, 91, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Roque, M.J.; de Ancos, B.; Sánchez-Moreno, C.; Cano, M.P.; Elez-Martínez, P.; Martín-Belloso, O. Impact of food matrix and processing on the in vitro bioaccessibility of vitamin C, phenolic compounds, and hydrophilic antioxidant activity from fruit juice-based beverages. J. Funct. Foods 2015, 14, 33–43. [Google Scholar] [CrossRef]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, and Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Hummel, B.C.W. A modified spectrophotometric determination of chymotrypsin, trypsin, and thrombin. Can. J. Biochem. Physiol. 1959, 37, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Marie, A.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in-vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Picone, G.; De Noni, I.; Ferranti, P.; Nicolai, M.A.; Alamprese, C.; Trimigno, A.; Brodkorb, A.; Portmann, R.; Pihlanto, A.; El, S.N.; et al. Monitoring molecular composition and digestibility of ripened bresaola through a combined foodomics approach. Food Res. Int. 2019, 115, 360–368. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Picone, G.; Pasini, F.; Caboni, M.F.; Gianotti, A.; Bordoni, A.; Capozzi, F. Olive oil industry by-products. Effects of a polyphenol-rich extract on the metabolome and response to inflammation in cultured intestinal cell. Food Res. Int. 2018, 113, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, A.; Laghi, L.; Babini, E.; Di Nunzio, M.; Picone, G.; Ciampa, A.; Valli, V.; Danesi, F.; Capozzi, F. The foodomics approach for the evaluation of protein bioaccessibility in processed meat upon in vitro digestion. Electrophoresis 2014, 35, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, A.; Picone, G.; Babini, E.; Vignali, M.; Danesi, F.; Valli, V.; Di Nunzio, M.; Laghi, L.; Capozzi, F. NMR comparison of in vitro digestion of Parmigiano Reggiano cheese aged 15 and 30 months. Magn. Reson. Chem. 2011, 49, S61–S70. [Google Scholar] [CrossRef] [PubMed]
- Valli, V.; Danesi, F.; Gianotti, A.; Di Nunzio, M.; Taneyo Saa, D.L.; Bordoni, A. Antioxidative and anti-inflammatory effect of in vitro digested cookies baked using different types of flours and fermentation methods. Food Res. Int. 2016, 88, 256–262. [Google Scholar] [CrossRef]
- Noh, Y.H.; Lee, D.B.; Lee, Y.W.; Pyo, Y.H. In Vitro Inhibitory Effects of Organic Acids Identified in Commercial Vinegars on α-Amylase and α-Glucosidase. Prev. Nutr. Food Sci. 2020, 25, 319–324. [Google Scholar] [CrossRef]
- Cirkovic Velickovic, T.D.; Stanic-Vucinic, D.J. The Role of Dietary Phenolic Compounds in Protein Digestion and Processing Technologies to Improve Their Antinutritive Properties. Compr. Rev. Food Sci. Food Saf. 2018, 17, 82–103. [Google Scholar] [CrossRef]
- Li, Y.; Lu, F.Q.; Feng, Y.; He, Z.D.; Wu, X.L. Binding interaction between (−)-epigallocatechin-3-gallate (EGCG) of green tea and pepsin. Acta Aliment. 2016, 45, 129–140. [Google Scholar] [CrossRef][Green Version]
- Liu, Z.; Wang, L.; Shi, L.; Chen, X.; Chang, Y.; Cao, Y.; Zhao, L. Investigation on the Interaction Behavior Between Oenothein B and Pepsin by Isothermal Titration Calorimetry and Spectral Studies. J. Food Sci. 2019, 84. [Google Scholar] [CrossRef]
- McDougall, G.; Kulkarni, N.; Stewart, D. Current developments on the inhibitory effects of berry polyphenols on digestive enzymes. Biofactors 2008, 34, 73–80. [Google Scholar] [CrossRef]
- Sinanoglou, V.; Zoumpoulakis, P.; Fotakis, C.; Kalogeropoulos, N.; Sakellari, A.; Karavoltsos, S.; Strati, I. On the Characterization and Correlation of Compositional, Antioxidant and Colour Profile of Common and Balsamic Vinegars. Antioxidants 2018, 7, 139. [Google Scholar] [CrossRef]
- Łoś, J.; Podsędek, A. Tannins from different foodstuffs as trypsin inhibitors. Pol. J. Food Nutr. Sci. 2004, 54, 5. [Google Scholar]
- Liu, Q.; Tang, G.-Y.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Antioxidant Activities, Phenolic Profiles, and Organic Acid Contents of Fruit Vinegars. Antioxidants 2019, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Burokerkilgore, M.; Wang, K.K.W. A Coomassie Brilliant Blue G-250-Based Colorimetric Assay for Measuring Activity of Calpain and Other Proteases. Anal. Biochem. 1993, 208, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cano, I.; Rocha-Mendoza, D.; Ortega-Anaya, J.; Wang, K.; Kosmerl, E.; Jiménez-Flores, R. Lactic acid bacteria isolated from dairy products as potential producers of lipolytic, proteolytic and antibacterial proteins. Appl. Microbiol. Biotechnol. 2019, 103, 5243–5257. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.; Moraes, W.M.; Silva, G.; Prestes, J.; Schoenfeld, B. Vinegar (acetic acid) intake on glucose metabolism: A narrative review. Clin. Nutr. Espen 2019, 32. [Google Scholar] [CrossRef] [PubMed]
- Masino, F.; Chinnici, F.; Bendini, A.; Montevecchi, G.; Antonelli, A. A study on relationships among chemical, physical, and qualitative assessment in traditional balsamic vinegar. Food Chem. 2008, 106, 90–95. [Google Scholar] [CrossRef]
- Vitali Čepo, D.; Radić, K.; Turčić, P.; Anić, D.; Komar, B.; Šalov, M. Food (Matrix) Effects on Bioaccessibility and Intestinal Permeability of Major Olive Antioxidants. Foods 2020, 9, 1831. [Google Scholar] [CrossRef]
- Turgeon, S.L.; Brisson, G. Symposium review: The dairy matrix-Bioaccessibility and bioavailability of nutrients and physiological effects. J. Dairy Sci. 2020, 103, 6727–6736. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; de Ancos, B.; Sánchez-Vega, R.; Sánchez-Moreno, C.; Cano, M.P.; Elez-Martínez, P.; Martín-Belloso, O. Food matrix and processing influence on carotenoid bioaccessibility and lipophilic antioxidant activity of fruit juice-based beverages. Food Funct. 2016, 7, 380–389. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pepsin | Pepsin with acetic acid ( 0.6% v/v) | Pepsin with BVM ( 10% v/v) | |
|---|---|---|---|
| Pepsin activity (U/mg) | 1926.61 ± 642.70 a | 1945.22 ± 615.57 a | 958.5 ± 526.13 b |
| Pancreatin | Pancreatin and acetic acid (f.c. 0.3% v/v) | Pancreatin and BVM (f.c. 5% v/v) | |
| Trypsin activity (U/mg) | 16.36 ± 2.62 a | 16.72 ± 2.16 a | 15.88 ± 1.73 a |
| Amylase activity (U/mg) | 27.25 ± 1.55 a | 3.14 ± 0.45 c | 8.12 ± 1.9 b |
| Lipase activity (mU/mg) | 1.37 ± 0.09 b | 1.34 ± 0.39 b | 5.96 ± 0.6 a |
| Region | Without BVM | With BVM | p |
|---|---|---|---|
| PRC | |||
| Carbohydrate | 18,928 ± 3008 | 22,754 ± 194 | 0.092 |
| Anomeric | 8357 ± 746 | 2439 ± 407 | 0.0003 |
| Bresaola | |||
| Carbohydrate | 50,725 ± 3755 | 39,106 ± 4310 | 0.024 |
| Anomeric | 2374 ± 244 | 1210 ± 575 | 0.032 |
| Boiled potatoes | |||
| Carbohydrate | 40,369 ± 5376 | 22,398 ± 2508 | 0.006 |
| Anomeric | 3416 ± 707 | 1760 ± 323 | 0.021 |
| Region | Without BVM | With BVM | p |
|---|---|---|---|
| PRC | |||
| α-Amino acid proton | 3443 ± 242 | 10,433 ± 935 | 0.0002 |
| Peptide | 851 ± 136 | 3212 ± 457 | 0.001 |
| Bresaola | |||
| α-Amino acid proton | 7295 ± 843 | 6368 ± 340 | 0.153 |
| Peptide | 2220 ± 123 | 1751 ± 307 | 0.070 |
| Boiled potatoes | |||
| α-Amino acid proton | 2546 ± 33.76 | 1975 ± 432 | 0.080 |
| Peptidic | 429 ± 16 | 488 ± 108 | 0.625 |
| Region | Without BVM | With BVM | p |
|---|---|---|---|
| PRC | |||
| Aliphatic | 3758 ± 1084 | 28,001 ± 2546 | 0.0001 |
| Aromatic | 6107 ± 386 | 9018 ± 771 | 0.004 |
| Bresaola | |||
| Aliphatic | 25,644 ± 1536 | 20,311 ± 2556 | 0.036 |
| Aromatic | 6368 ± 527 | 5718 ± 718 | 0.275 |
| Boiled potatoes | |||
| Aliphatic | 5975 ± 693 | 5110 ± 2028 | 0.523 |
| Aromatic | 1264 ± 281 | 1049 ± 602 | 0.605 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbinati, E.; Di Nunzio, M.; Picone, G.; Chiarello, E.; Bordoni, A.; Capozzi, F. The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix. Foods 2021, 10, 411. https://doi.org/10.3390/foods10020411
Urbinati E, Di Nunzio M, Picone G, Chiarello E, Bordoni A, Capozzi F. The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix. Foods. 2021; 10(2):411. https://doi.org/10.3390/foods10020411
Chicago/Turabian StyleUrbinati, Eleonora, Mattia Di Nunzio, Gianfranco Picone, Elena Chiarello, Alessandra Bordoni, and Francesco Capozzi. 2021. "The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix" Foods 10, no. 2: 411. https://doi.org/10.3390/foods10020411
APA StyleUrbinati, E., Di Nunzio, M., Picone, G., Chiarello, E., Bordoni, A., & Capozzi, F. (2021). The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix. Foods, 10(2), 411. https://doi.org/10.3390/foods10020411