
Listeria monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR


Abstract
1. Introduction
2. Materials and Methods
2.1. Ready-to-Eat (RTE) Salad Production Process
2.2. Ready-to-Eat Salad Sampling and Storage during Shelf-Life Testing
2.3. Physicochemical Analyses
2.3.1. Water Activity (aw) Determination
2.3.2. Potential of Hydrogen (pH) Determination
2.4. Microbiological Analyses
2.5. L. monocytogenes Confirmation and Serogrouping
2.6. Pulsed-Field Gel Electrophoresis Typing
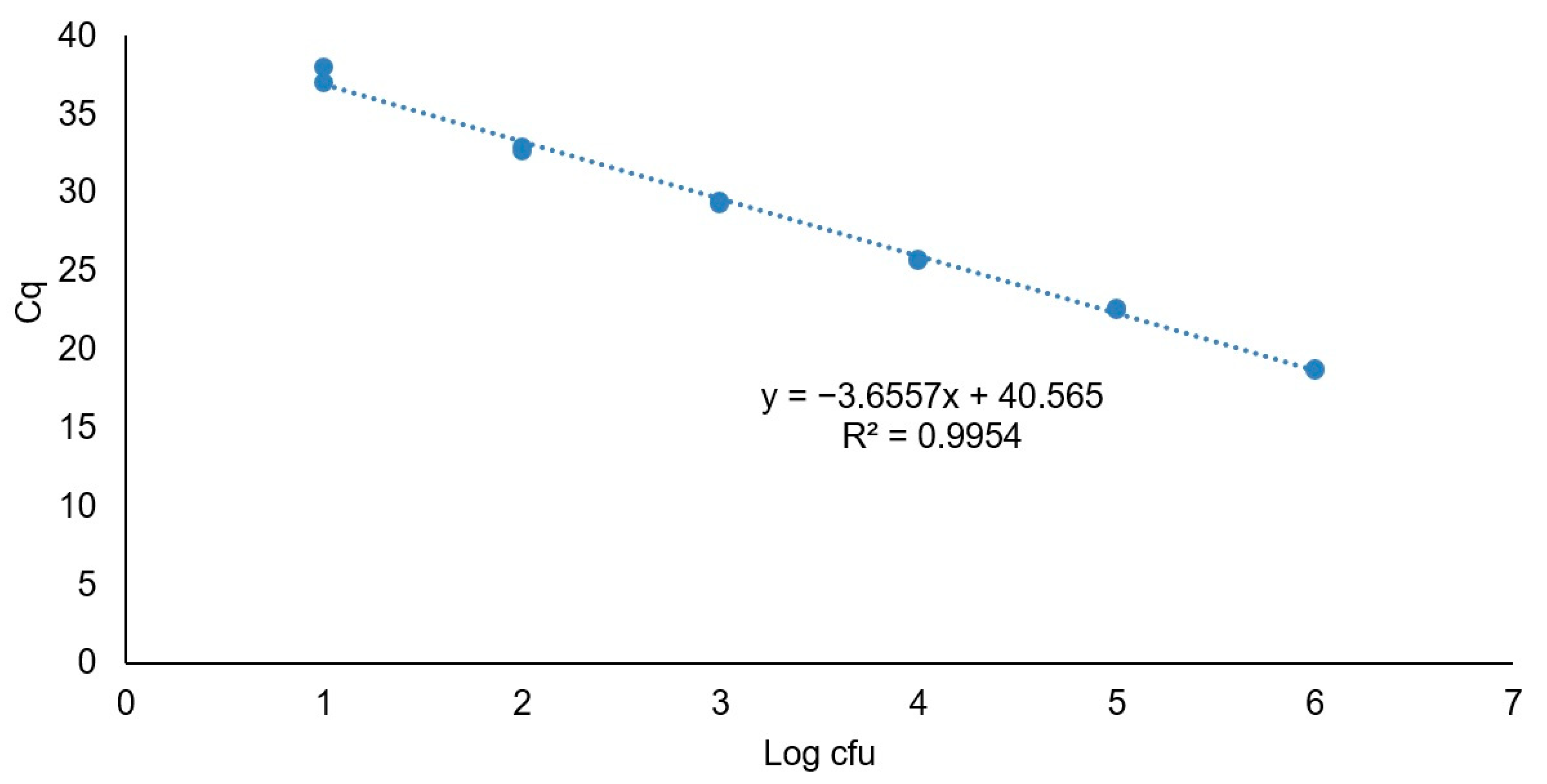
2.7. PMAxx-qPCR Assay
2.7.1. Bacterial Strains Preparation
2.7.2. PMAxx Treatment and Genomic DNA Extraction
2.8. Statistical Analyses
3. Results
3.1. Physicochemical Analyses—aw and pH Measurements
3.2. Microbiological Analyses
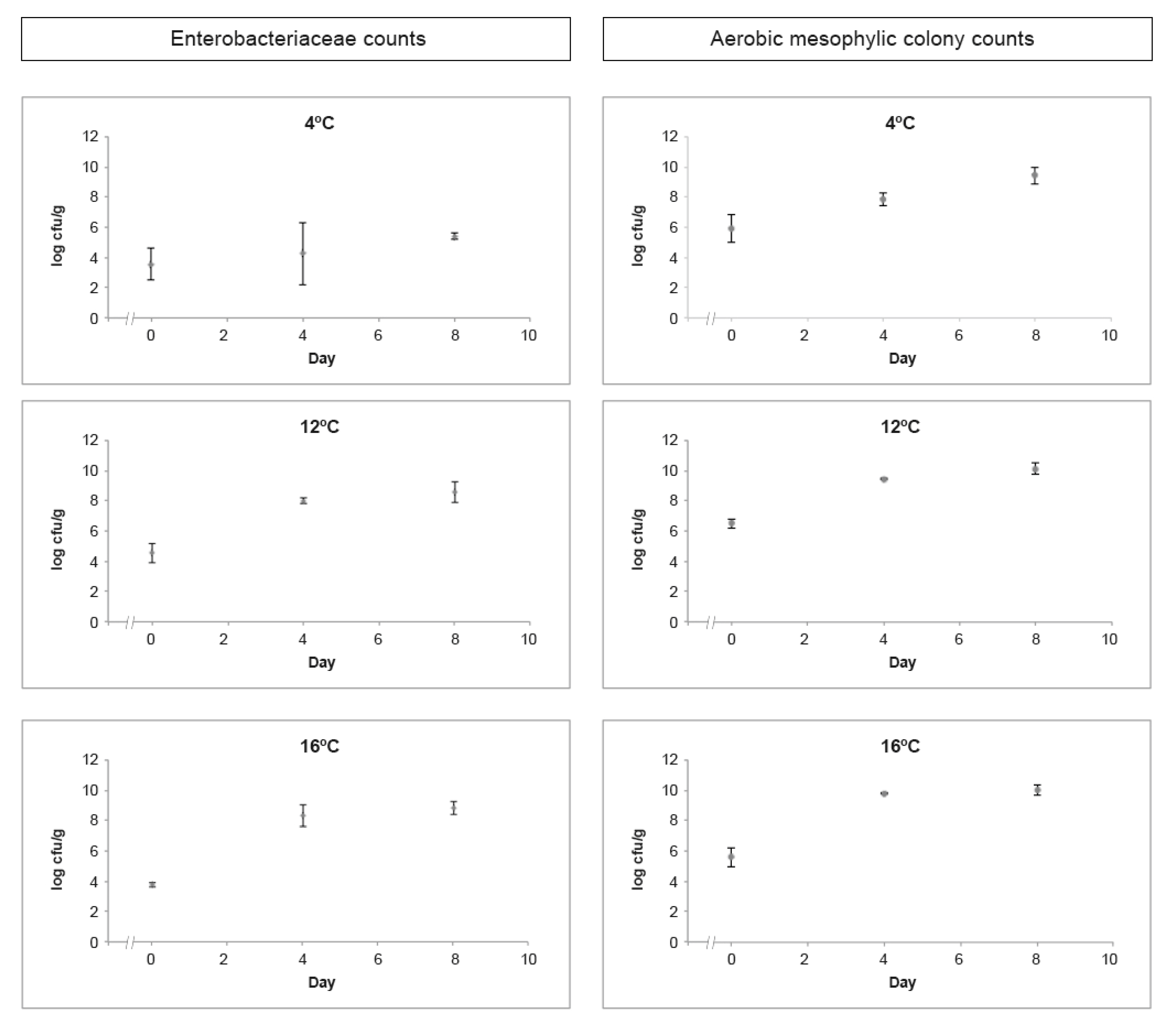
3.2.1. Hygiene Indicators
3.2.2. Detection and Enumeration of L. monocytogenes, Confirmation, and Serogrouping
3.2.3. PFGE Typing
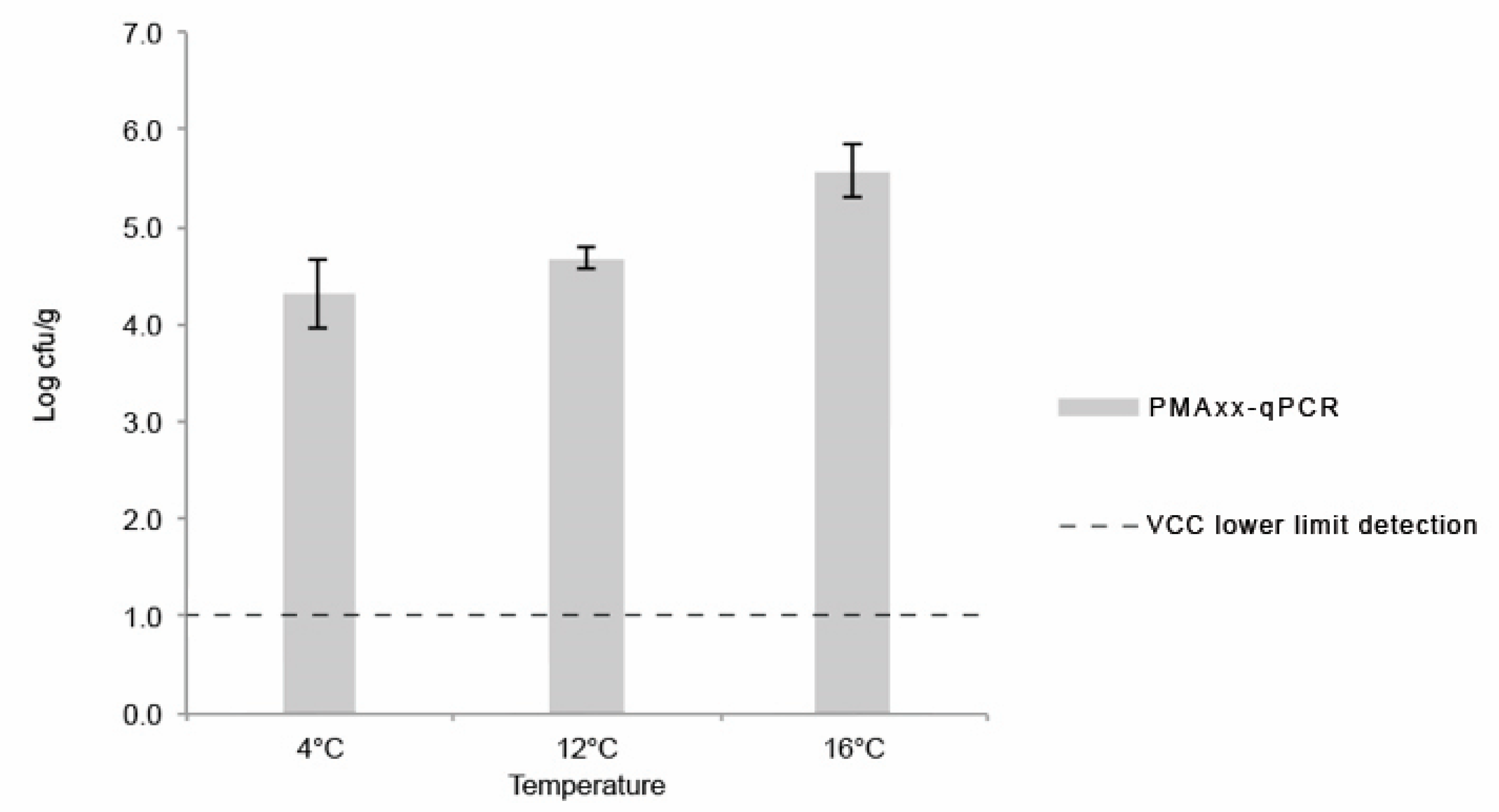
3.2.4. PMAxx-qPCR Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gullino, M.L.; Gilardi, G.; Garibaldi, A. Ready-to-eat salad crops: A plant pathogen’s heaven. Plant Dis. 2019, 103, 2153–2170. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.A.; Germano, P.M.L. Listeria monocytogenes in ready-to-eat, sliced, cooked ham and salami products, marketed in the city of São Paulo, Brazil: Occurrence, quantification, and serotyping. Food Control 2011, 22, 297–302. [Google Scholar] [CrossRef]
- Soderqvist, K. Is your lunch salad safe to eat? Occurrence of bacterial pathogens and potential for pathogen growth in pre-packed ready-to-eat mixed-ingredient salads. Infect. Ecol. Epidemiol. 2017, 7, 1407216. [Google Scholar] [CrossRef]
- Tirloni, E.; Stella, S.; de Knegt, L.V.; Gandolfi, G.; Bernardi, C.; Nauta, M.J. A quantitative microbial risk assessment model for Listeria monocytogenes in RTE sandwiches. Microb. Risk Anal. 2018, 9, 11–21. [Google Scholar] [CrossRef]
- Ziegler, M.; Kent, D.; Stephan, R.; Guldimann, C. Growth potential of Listeria monocytogenes in twelve different types of RTE salads: Impact of food matrix, storage temperature and storage time. Int. J. Food Microbiol. 2019, 296, 83–92. [Google Scholar] [CrossRef]
- Chau, M.L.; Aung, K.T.; Hapuarachchi, H.C.; Lee, P.S.; Lim, P.Y.; Kang, J.S.; Ng, Y.; Yap, H.M.; Yuk, H.G.; Gutiérrez, R.A.; et al. Microbial survey of ready-to-eat salad ingredients sold at retail reveals the occurrence and the persistence of Listeria monocytogenes Sequence Types 2 and 87 in pre-packed smoked salmon. BMC Microbiol. 2017, 17, 1–13. [Google Scholar] [CrossRef]
- Sahu, S.N.; Kim, B.; Ferguson, M.S.; Zink, D.L.; Datta, A.R. Growth potential of Listeria monocytogenes in artificially contaminated celery and chicken salad. Food Control 2016, 73, 1229–1236. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. Available online: https://www.ecdc.europa.eu/en/publications-data/european-union-one-health-2018-zoonoses-report (accessed on 2 August 2020).
- United States of America Centers for Disease Control and Prevention. Listeria (listeriosis) Prevention. Available online: https://www.cdc.gov/listeria/prevention.html (accessed on 4 August 2020).
- European Commission. Commmission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, 1–25. [Google Scholar] [CrossRef]
- Auvolat, A.; Besse, N.G. The challenge of enumerating Listeria monocytogenes in food. Food Microbiol. 2016, 53, 135–149. [Google Scholar] [CrossRef]
- Barre, L.; Brasseur, E.; Doux, C.; Lombard, B.; Besse, N.G. Sensitive enumeration of Listeria monocytogenes and other Listeria species in various naturally contaminated matrices using a membrane filtration method. Food Microbiol. 2015, 48, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Maestu, A.; Chapela, M.-J.; Peñaranda, E.; Cabado, A.G. Re-evaluation of enhanced qPCR prevalidated method for next-day detection of Salmonella spp., Shigella spp., Escherichia coli O157 and Listeria monocytogenes. Food Biotechnol. 2015, 29, 317–335. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Azinheiro, S.; Fuciños, P.; Carvalho, J.; Prado, M. Comparative study of multiplex real-time recombinase polymerase amplification and ISO 11290-1 methods for the detection of Listeria monocytogenes in dairy products. Food Microbiol. 2020, 92, 1–9. [Google Scholar] [CrossRef]
- He, P.; Zhu, G.; Luo, J.; Wang, H.; Yan, Y.; Chen, L.; Gao, W.; Chen, Z. Development and application of a one-tube multiplex real-time PCR with melting curve analysis for simultaneous detection of five foodborne pathogens in food samples. J. Food Saf. 2017, 37, e12297. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Martínez-Garcia, M.; Capellas, M.; Yuste, J.; Fung, D.Y.C.; Rodríguez-Jerez, J.-J. From hazard analysis to risk control using rapid methods in microbiology: A practical approach for the food industry. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1877–1907. [Google Scholar] [CrossRef]
- Xu, Y.-G.; Liu, Z.-M.; Zhang, B.-Q.; Qu, M.; Mo, C.-S.; Luo, J.; Li, S.-L. Development of a novel target-enriched multiplex PCR (Tem-PCR) assay for simultaneous detection of five foodborne pathogens. Food Control 2016, 64, 54–59. [Google Scholar] [CrossRef]
- Agustí, G.; Fittipaldi, M.; Codony, F. Optimization of a viability PCR method for the detection of Listeria monocytogenes in food samples. Curr. Microbiol. 2018, 75, 779–785. [Google Scholar] [CrossRef]
- Molinos, A.C.; Abriouel, H.; Ben-Omar, N.; Martinez-Canamero, M.; Gálvez, A. A quantitative real-time PCR assay for quantification of viable Listeria monocytogenes cells after bacteriocin injury in food-first insights. Curr. Microbiol. 2010, 61, 515–519. [Google Scholar] [CrossRef]
- Postollec, F.; Falentin, H.; Pavan, S.; Combrisson, J.; Sohier, D. Recent advances in quantitative PCR (qPCR) applications in food microbiology. Food Microbiol. 2011, 28, 848–861. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Azinheiro, S.; Carvalho, J.; Prado, M. Rapid and sensitive detection of viable Listeria monocytogenes in food products by a filtration-based protocol and qPCR. Food Microbiol. 2018, 73, 254–263. [Google Scholar] [CrossRef]
- Truchado, P.; Gil, M.I.; Kostic, T.; Allende, A. Optimization and validation of a PMA qPCR method for Escherichia coli quantification in primary production. Food Control 2016, 62, 150–156. [Google Scholar] [CrossRef]
- D’Urso, O.F.; Poltronieri, P.; Marsigliante, S.; Storelli, C.; Hernández, M.; Rodríguez-Lázaro, D. A filtration-based real-time PCR method for the quantitative detection of viable Salmonella enterica and Listeria monocytogenes in food samples. Food Microbiol. 2009, 26, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Cheung, C.Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods. 2006, 67, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tong, T.; Zeng, S.; Lin, Y.; Wu, S.; He, M. Quantification of viable bacteria in wastewater treatment plants by using propidium monoazide combined with quantitative PCR (PMA-qPCR). J. Environ. Sci. (China) 2014, 26, 299–306. [Google Scholar] [CrossRef]
- Elizaquível, P.; Sánchez, G.; Aznar, R. Quantitative detection of viable foodborne E. coli O157:H7, Listeria monocytogenes and Salmonella in fresh-cut vegetables combining propidium monoazide and real-time PCR. Food Control 2012, 25, 704–708. [Google Scholar] [CrossRef]
- Chen, J.-Q.; Regan, P.; Laksanalamai, P.; Healey, S.; Hu, Z. Prevalence and methodologies for detection, characterization and subtyping of Listeria monocytogenes and L. ivanovii in foods and environmental sources. Food Sci. Hum. Well. 2017, 6, 97–120. [Google Scholar] [CrossRef]
- Halpin, J.L.; Garrett, N.M.; Ribot, E.M.; Graves, L.M.; Cooper, K.L. Re-evaluation, optimization, and multilaboratory validation of the PulseNet-standardized pulsed-field gel electrophoresis protocol for Listeria monocytogenes. Foodborne Pathog. Dis. 2010, 7, 293–298. [Google Scholar] [CrossRef]
- Jadhav, S.; Bhave, M.; Palombo, E.A. Methods used for the detection and subtyping of Listeria monocytogenes. J. Microbiol. Methods. 2012, 88, 327–341. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Foodstuffs-Determination of Water Activity; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Norma Portuguesa (NP). Carnes, Derivados e Produtos Cárneos—Determinação do pH. Método de Referência; Instituto Portugês da Qualidade: Lisboa, Portugal, 1990. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs—Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination—Part 2: Specific Rules for the Preparation of Meat and Meat Products; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Enterobacteriaceae—Part 2: Colony-Count Technique; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain—Horizontal Method for the Enumeration of Microorganisms—Part 1: Colony Count at 30 °C by the Pour Plate Technique; International Organization for Standardization: Geneva, Switzerland, 2019. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and of Listeria spp.—Part 1: Detection Method; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- International Organization for Standardization (ISO). Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria—Part 2: Enumeration Method; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Kérouanton, A.; Marault, M.; Petit, L.; Grout, J.; Dao, T.T.; Brisabois, A. Evaluation of a multiplex PCR assay as an alternative method for Listeria monocytogenes serotyping. J. Microbiol. Methods 2010, 80, 134–137. [Google Scholar] [CrossRef]
- Graves, L.M.; Swaminathan, B. PulseNet standardized protocol for subtyping Listeria monocytogenes by macrorestriction and pulsed-field gel electrophoresis. Int. J. Food Microbiol. 2001, 65, 55–62. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, L.; Xu, H.; Aguilar, Z.P.; Liu, C.; Gan, B.; Xiong, Y.; Lai, W.; Xu, F.; Wei, H. Detection of non-emetic and emetic Bacillus cereus by propidium monoazide multiplex PCR (PMA-mPCR) with internal amplification control. Food Control 2014, 35, 401–406. [Google Scholar] [CrossRef]
- Josefsen, M.H.; Löfström, C.; Hansen, T.B.; Christensen, L.S.; Olsen, J.E.; Hoorfar, J. Rapid quantification of viable Campylobacter bacteria on chicken carcasses, using real-time PCR and propidium monoazide treatment, as a tool for quantitative risk assessment. Appl. Environ. Microbiol. 2010, 76, 5097–5104. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.; Saunders, N.; Owen, R. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol. 1989, 8, 151–156. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. Primer sequence disclosure: A clarification of the MIQE guidelines. Clin. Chem. 2011, 57, 919–921. [Google Scholar] [CrossRef] [PubMed]
- European Union Reference Laboratory for Listeria monocytogenes. Technical Guidance Document for Conducting Shelf-Life Studies on Listeria monocytogenes in Ready-to-Eat Foods. Available online: https://ec.europa.eu/food/sites/food (accessed on 4 December 2020).
- Manios, S.G.; Konstantinidis, N.; Gounadaki, A.S.; Skandamis, P.N. Dynamics of low (1–4 cells) vs high populations of Listeria monocytogenes and Salmonella Typhimurium in fresh-cut salads and their sterile liquid or solidified extracts. Food Control 2013, 29, 318–327. [Google Scholar] [CrossRef]
- Santos, T.; Campos, F.; Padovani, N.; Dias, M.; Mendes, M.; Maffei, D. Assessment of the microbiological quality and safety of minimally processed vegetables sold in Piracicaba, SP, Brazil. Lett. Appl. Microbiol. 2020, 71, 187–194. [Google Scholar] [CrossRef]
- Health Protection Agency. Guidelines for Assessing the Microbiological Safety of Ready-to-Eat Foods; HPA: London, UK, 2009.
- European Commission. Risk Profile on the Microbiological Contamination of Fruits and Vegetables Eaten Raw—Report of the Scientific Committee on Food (SCF/CS/FMH/SURF/Final–29 April 2002). Available online: https://www.adiveter.com/ftp_public/articulo358.pdf (accessed on 17 July 2020).
- Food Safety Authority of Ireland. Guidelines for the Interpretation of Results of Microbiological Testing of Ready-to-Eat Foods Placed on the Market. Available online: https://www.fsai.ie/publications_GN3_microbiological_limits/ (accessed on 1 August 2020).
- Omac, B.; Moreira, R.G.; Castell-Perez, E. Quantifying growth of cold-adapted Listeria monocytogenes and Listeria innocua on fresh spinach leaves at refrigeration temperatures. J. Food Eng. 2018, 224, 17–26. [Google Scholar] [CrossRef]
- Skalina, L.; Nikolajeva, V. Growth potential of Listeria monocytogenes strains in mixed ready-to-eat salads. Int. J. Food Microbiol. 2010, 144, 317–321. [Google Scholar] [CrossRef]
- Kotzekidou, P. Microbiological examination of ready-to-eat foods and ready-to-bake frozen pastries from university canteens. Food Microbiol. 2013, 34, 337–343. [Google Scholar] [CrossRef]
- Moravkova, M.; Verbikova, V.; Michna, V.; Babak, V.; Cahlikova, H.; Karpiskova, R.; Kralik, P. Detection and quantification of Listeria monocytogenes in ready-to-eat vegetables, frozen vegetables and sprouts examined by culture methods and real-time PCR. J. Food Nutr. Res. 2017, 5, 832–837. [Google Scholar] [CrossRef]
- Hamidiyan, N.; Salehi-Abargouei, A.; Rezaei, Z.; Dehghani-Tafti, R.; Akrami-Mohajeri, F. The prevalence of Listeria spp. food contamination in Iran: A systematic review and meta-analysis. Food Res. Int. 2018, 107, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Public Health England. Detection and Enumeration of Listeria monocytogenes and other Listeria Species National Infection Service Food Water and Environmental Microbiology Standard Method. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/768777/detection_and_enumeration_of_listeria_monocytogenes_and_other_listeria_species.pdf (accessed on 17 September 2020).
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Maćkiw, E.; Modzelewska, M.; Mąka, Ł.; Ścieżyńska, H.; Pawłowska, K.; Postupolski, J.; Korsak, D. Antimicrobial resistance profiles of Listeria monocytogenes isolated from ready-to-eat products in Poland in 2007–2011. Food Control 2016, 59, 7–11. [Google Scholar] [CrossRef]
- Amajoud, N.; Leclercq, A.; Soriano, J.M.; Bracq-Dieye, H.; Maadoudi, M.; Skali-Senhaji, N.; Kounnoun, A.; Moura, A.; Lecuit, M.; Abrini, J. Prevalence of Listeria spp. and characterization of Listeria monocytogenes isolated from food products in Tetouan, Morocco. Food Control 2017, 84, 436–441. [Google Scholar] [CrossRef]
- Eskhan, A.O.; Abu-Lail, N.I. Cellular and molecular investigations of the adhesion and mechanics of Listeria monocytogenes lineages I and II environmental and epidemic strains. J. Colloid Interface Sci. 2013, 394, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef]
- Véghová, A.; Minarovičová, J.; Koreňová, J.; Drahovská, H.; Kaclíková, E. Prevalence and tracing of persistent Listeria monocytogenes strains in meat processing facility production chain. J. Food Saf. 2017, 37, e12315. [Google Scholar] [CrossRef]
- Henriques, A.R.; Telo da Gama, L.; Fraqueza, M.J. Tracking Listeria monocytogenes contamination and virulence-associated characteristics in the ready-to-eat meat-based food products industry according to the hygiene level. Int. J. Food Microbiol. 2017, 242, 101–106. [Google Scholar] [CrossRef]
- Montero, D.; Bodero, M.; Riveros, G.; Lapierre, L.; Gaggero, A.; Vidal, R.M.; Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 1–8. [Google Scholar] [CrossRef]
- Yu, T.; Jiang, X. Prevalence and characterization of Listeria monocytogenes isolated from retail food in Henan, China. Food Control 2014, 37, 228–231. [Google Scholar] [CrossRef]
- Scariot, M.C.; Venturelli, G.L.; Prudêncio, E.S.; Arisi, A.C.M. Quantification of Lactobacillus paracasei viable cells in probiotic yoghurt by propidium monoazide combined with quantitative PCR. Int. J. Food Microbiol. 2018, 264, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Gil, M.I.; Larrosa, M.; Allende, A. Detection and quantification methods for viable but non-culturable (VBNC) cells in process wash water of fresh-cut produce: Industrial validation. Front. Microbiol. 2020, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Barretta, C.; Verruck, S.; Maran, B.M.; Maurício, L.S.; Miotto, M.; Vieira, C.R.W.; Prudencio, E.S. Listeria monocytogenes survival in raw Atlantic salmon (Salmo salar) fillet under in vitro simulated gastrointestinal conditions by culture, qPCR and PMA-qPCR detection methods. LWT 2019, 107, 132–137. [Google Scholar] [CrossRef]
- Nocker, A.; Camper, A.K. Novel approaches toward preferential detection of viable cells using nucleic acid amplification techniques. FEMS Microbiol. Lett. 2009, 291, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Overney, A.; Chassaing, D.; Carpentier, B.; Guillier, L.; Firmesse, O. Development of synthetic media mimicking food soils to study the behaviour of Listeria monocytogenes on stainless steel surfaces. Int. J. Food Microbiol. 2016, 238, 7–14. [Google Scholar] [CrossRef]
- Alessandria, V.; Rantsiou, K.; Dolci, P.; Cocolin, L. Molecular methods to assess Listeria monocytogenes route of contamination in a dairy processing plant. Int. J. Food Microbiol. 2010, 141, S156–S162. [Google Scholar] [CrossRef]
- Muhterem-Uyar, M.; Dalmasso, M.; Bolocan, A.S.; Hernandez, M.; Kapetanakou, A.E.; Kuchta, T.; Manios, S.G.; Melero, B.; Minarovičová, J.; Nicolau, A.I.; et al. Environmental sampling for Listeria monocytogenes control in food processing facilities reveals three contamination scenarios. Food Control 2015, 51, 94–107. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Anchor Nucleotide | Amplicon Length (bp) |
|---|---|---|---|
| iap | AF500174.1 | 285 | 124 |
| Storage Temperature | Day 0 | Day 4 | Day 8 | |
|---|---|---|---|---|
| aw | 4 °C | 0.955 ± 0.001 | 0.957 ± 0.006 | 0.962 ± 0.009 |
| 12 °C | 0.966 ± 0.001 | 0.957 ± 0.002 | 0.971 ± 0.008 | |
| 16 °C | 0.980 ± 0.002 | 0.958 ± 0.001 | 0.963 ± 0.007 | |
| pH | 4 °C | 5.930 ± 0.033 | 6.113 ± 0.191 | 6.251 ± 0.561 |
| 12 °C | 6.021 ± 0.080 | 6.427 ± 0.400 | 6.632 ± 0.554 | |
| 16 °C | 6.647 ± 0.491 | 7.309 ± 0.325 | 7.625 ± 0.167 | |
| Salad Code | Incubation Temperature | Presumptive Colonies (n) Collected throughout Shelf-Life | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 4 | Day 8 | ||||||||
| A a | B a | C a | A a | B a | C a | A a | B a | C a | ||
| 1 | 4 °C | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| 2 | 4 °C | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 3 | 4 °C | 1 | 1 | 0 | 3 | 1 | 0 | 0 | 0 | 0 |
| 4 | 12 °C | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 5 | 12 °C | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 |
| 6 | 12 °C | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 | 16 °C | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 2 | 0 |
| 8 | 16 °C | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 0 | 0 |
| 9 | 16 °C | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 1 | 0 |
| Serogroup | Proportion | Isolate Code |
|---|---|---|
| IIa | 1 (10%) | i108 |
| IIb | 1 (10%) | i187 |
| IVb | 8 (80%) | i380, i301, i341, i342, i343, i543, i546, i782 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardo, R.; Duarte, A.; Tavares, L.; Barreto, A.S.; Henriques, A.R. Listeria monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR. Foods 2021, 10, 235. https://doi.org/10.3390/foods10020235
Bernardo R, Duarte A, Tavares L, Barreto AS, Henriques AR. Listeria monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR. Foods. 2021; 10(2):235. https://doi.org/10.3390/foods10020235
Chicago/Turabian StyleBernardo, Rita, Ana Duarte, Luís Tavares, António Salvador Barreto, and Ana Rita Henriques. 2021. "Listeria monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR" Foods 10, no. 2: 235. https://doi.org/10.3390/foods10020235
APA StyleBernardo, R., Duarte, A., Tavares, L., Barreto, A. S., & Henriques, A. R. (2021). Listeria monocytogenes Assessment in a Ready-to-Eat Salad Shelf-Life Study Using Conventional Culture-Based Methods, Genetic Profiling, and Propidium Monoazide Quantitative PCR. Foods, 10(2), 235. https://doi.org/10.3390/foods10020235