
Vitamin E Supplementation Enhances Lipid Oxidative Stability via Increasing Vitamin E Retention, Rather Than Gene Expression of MAPK-Nrf2 Signaling Pathway in Muscles of Broilers

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds and Treatment
2.2. Sampling
2.3. Determination of α-Tocopherol (VE) in Feed and Muscles
2.4. Lipid Oxidation and Antioxidant Capacity in Muscles and Serum
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
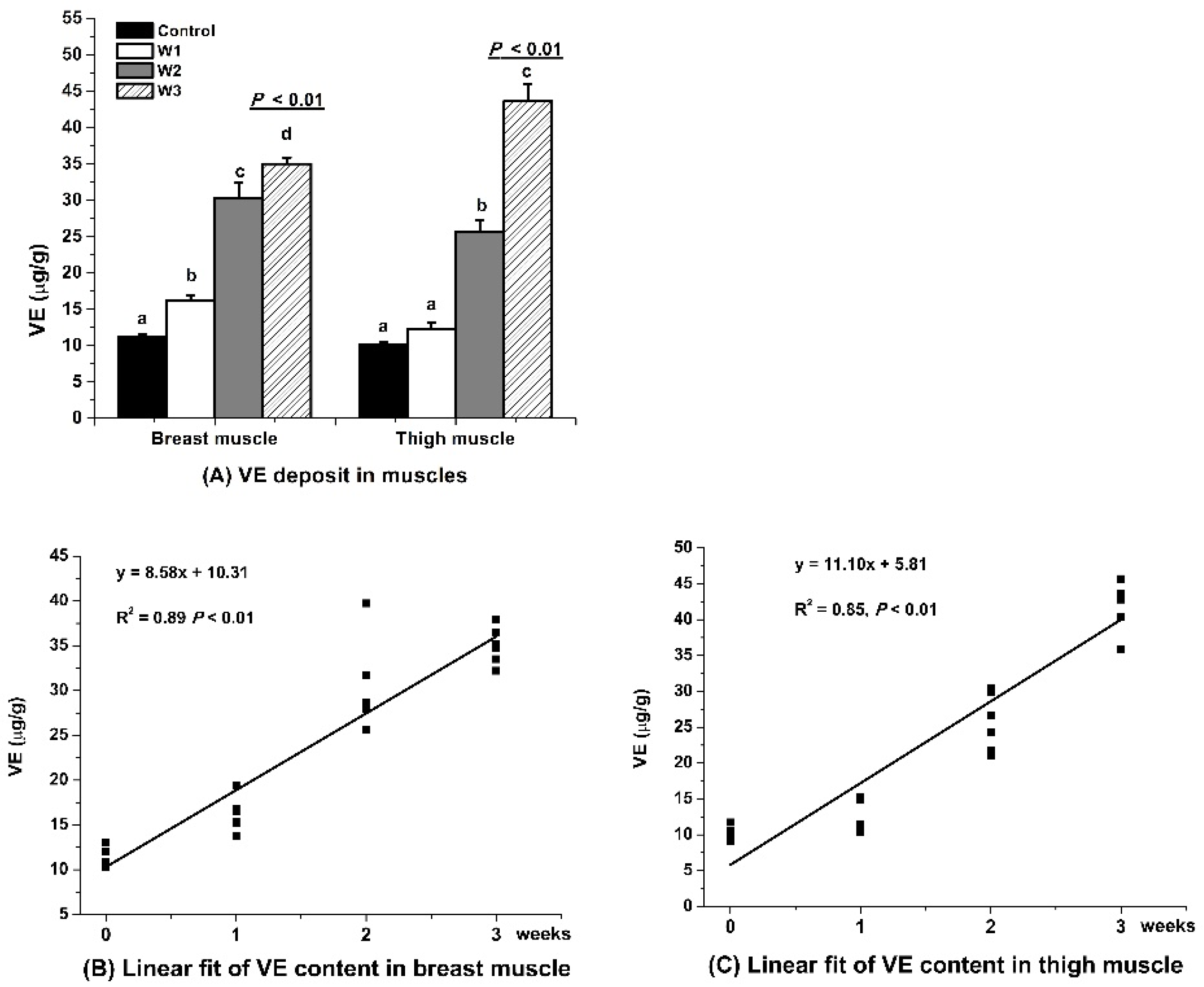
3.1. VE Retention
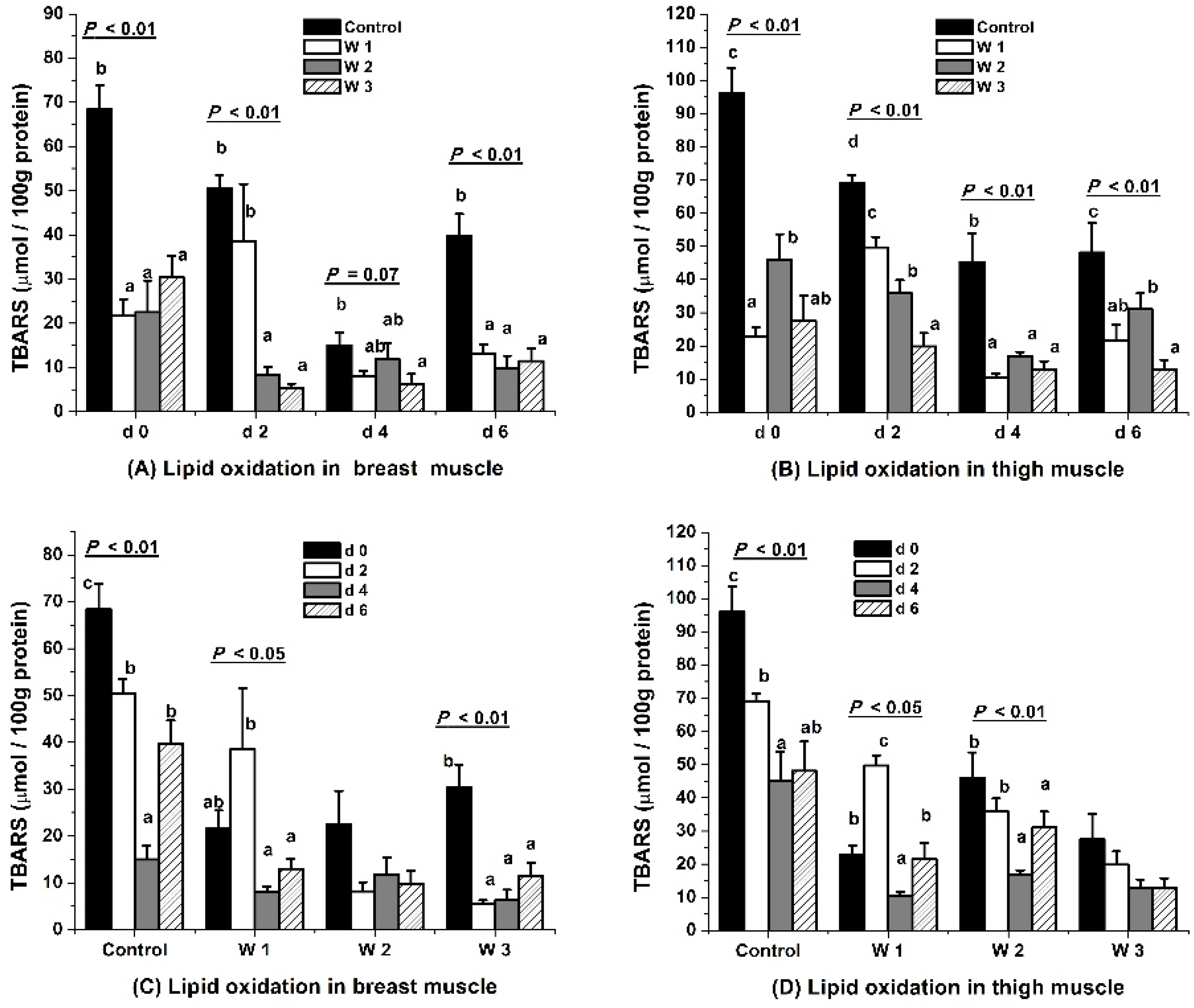
3.2. Lipid Oxidative Stability
3.3. Antioxidant Capacity
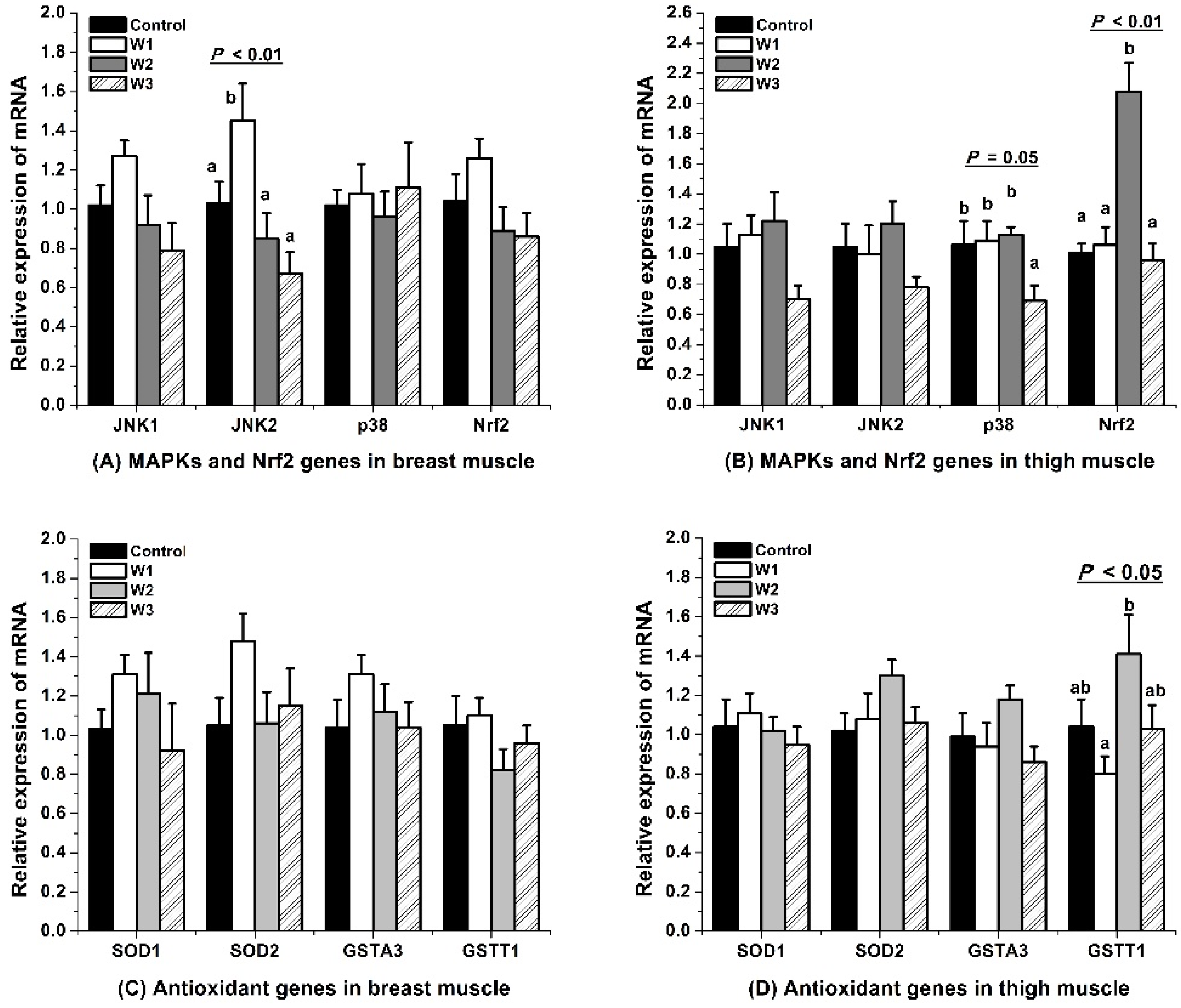
3.4. MAPK-Nrf2 Signaling Pathway
3.5. Correlations among Variables
4. Discussion
4.1. VE Retention
4.2. Lipid Oxidative Stability
4.3. Antioxidant Capacity
4.4. MAPK-Nrf2 Signaling Pathway
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, J.; Ding, X.M.; Bai, S.P.; Wang, J.P.; Zeng, Q.F.; Peng, H.W.; Xuan, Y.; Su, Z.W.; Zhang, K.Y. Effects of dietary vitamin E supplementation on laying performance, hatchability, and antioxidant status in molted broiler breeder hens. J. Appl. Poult. Res. 2021, 30, 100184. [Google Scholar] [CrossRef]
- Zingg, J.M. Vitamin E: Regulatory role on signal transduction. IUBMB Life 2019, 71, 456–478. [Google Scholar] [CrossRef]
- Wojtasik-Kalinowska, I.; Guzek, D.; Gorska-Horczyczak, E.; Głąbska, D.; Brodowska, M.; Sun, D.W.; Wierzbicka, A. Volatile compounds and fatty acids profile in longissimus dorsi muscle from pigs fed with feed containing bioactive components. LWT-Food Sci. Technol. 2016, 67, 112–117. [Google Scholar] [CrossRef]
- Pan, J.H.; Feng, L.; Jiang, W.D.; Wu, P.; Kuang, S.Y.; Tang, L.; Zhang, Y.A.; Zhou, X.Q.; Liu, Y. Vitamin E deficiency depressed fish growth, disease resistance, and the immunity and structural integrity of immune organs in grass carp (Ctenopharyngodon idella): Referring to NF-κB, TOR and Nrf2 signaling. Fish. Shellfish. Immunol. 2017, 60, 219–236. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Lee, E.J.; Ahn, D.U. Consumption of oxidized oil increases oxidative stress in broilers and affects the quality of breast meat. J. Agric. Food Chem. 2011, 59, 969–974. [Google Scholar] [CrossRef] [Green Version]
- Imik, H.; Atasever, M.A.; Urcar, S.; Ozlu, H.; Gumus, R.; Atasever, M. Meat quality of heat stress exposed broilers and effect of protein and vitamin E. Br. Poult. Sci. 2012, 53, 689–698. [Google Scholar] [CrossRef]
- Bartov, I.; Frigg, M. Effect of high concentrations of dietary vitamin E during various age periods on performance, plasma vitamin E and meat stability of broiler chicks at 7 weeks of age. Br. Poult. Sci. 1992, 33, 393–402. [Google Scholar] [CrossRef]
- Morrissey, P.A.; Brandon, S.; Buckley, D.J.; Sheehy, P.J.; Frigg, M. Tissue content of alpha-tocopherol and oxidative stability of broilers receiving dietary alpha-tocopheryl acetate supplement for various periods pre-slaughter. Br. Poult. Sci. 1997, 38, 84–88. [Google Scholar] [CrossRef]
- Rostami, H.; Seidavi, A.; Dadashbeiki, M.; Asadpour, Y.; Simões, J.; Laudadio, V.; Milis, C.; Tufarelli, V. Oxidative stability of chilled broiler breast meat as affected by dietary supplementation with rosemary (Rosmarinus officinalis L.) powder and vitamin E. Food Sci. Nutr. 2017, 5, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Narciso-Gaytán, C.; Shin, D.; Sams, A.R.; Keeton, J.T.; Miller, R.K.; Smith, S.B.; Sánchez-Plata, M.X. Dietary lipid source and vitamin E effect on lipid oxidation stability of refrigerated fresh and cooked chicken meat. Poult. Sci. 2010, 89, 2726–2734. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wu, S.G.; Zhang, H.J.; Zhang, L.; Yue, H.Y.; Ji, F.; Qi, G.H. Comparison of lipid oxidation, messenger ribonucleic acid levels of avian uncoupling protein, avian adenine nucleotide translocator, and avian peroxisome proliferator-activated receptor-γ coactivator-1α in skeletal muscles from electrical- and gas-stunned broilers. Poult. Sci. 2011, 90, 2069–2075. [Google Scholar]
- Xu, L.; Zhang, H.J.; Yue, H.Y.; Wu, S.G.; Yang, H.M.; Qi, G.H.; Wang, Z.Y. Low-current & high-frequency electrical stunning increased oxidative stress, lipid peroxidation, and gene transcription of the mitogen-activated protein kinase/nuclear factor-erythroid 2-related factor 2/antioxidant responsive element (MAPK/Nrf2/ARE) signaling pathway in breast muscle of broilers. Food Chem. 2018, 242, 491–496. [Google Scholar]
- Bostami, A.B.M.R.; Mun, H.S.; Dilawar, M.A.; Baek, K.S.; Yang, C.J. Carcass characteristics, proximate composition, fatty acid profile, and oxidative stability of pectoralis major and flexor cruris medialis muscle of broiler chicken subjected to with or without level of electrical stunning, slaughter, and subsequent bleeding. Animals 2021, 11, 1679. [Google Scholar]
- Peh, H.Y.; Tan, W.S.; Liao, W.; Wong, W.S. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Tian, Y.; Xiao, Y.; Wang, B.; Sun, C.; Tang, K.; Sun, F. Vitamin E and lycopene reduce coal burning fluorosis-induced spermatogenic cell apoptosis via oxidative stress-mediated JNK and ERK signaling pathways. Biosci. Rep. 2018, 38, BSR20171003. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Xu, Y.; Ren, X.; Xiang, D.; Lei, K.; Zhang, C.; Liu, D. Vitamin E ameliorates lipid metabolism in mice with nonalcoholic fatty liver disease via Nrf2/CES1 signaling pathway. Dig. Dis. Sci. 2019, 64, 3182–3191. [Google Scholar] [CrossRef]
- Fang, J.; Yin, H.; Yang, Z.; Tan, M.; Wang, F.; Chen, K.; Zuo, Z.; Shu, G.; Cui, H.; Ouyang, P.; et al. Vitamin E protects against cadmium-induced sub-chronic liver injury associated with the inhibition of oxidative stress and activation of Nrf2 pathway. Ecotoxicol. Environ. Saf. 2021, 208, 111610. [Google Scholar] [CrossRef]
- Shen, L.; Tang, X.; Wei, Y.; Long, C.; Tan, B.; Wu, S.; Sun, M.; Zhou, Y.; Cao, X.; Wei, G. Vitamin E and vitamin C attenuate Di-(2-ethylhexyl) phthalate-induced blood-testis barrier disruption by p38 MAPK in immature SD rats. Reprod. Toxicol. 2018, 81, 17–27. [Google Scholar] [CrossRef]
- Wong, S.K.; Mohamad, N.V.; Ibrahim, N.; Chin, K.Y.; Shuid, A.N.; Ima-Nirwana, S. The molecular mechanism of vitamin E as a bone-protecting agent: A review on current evidence. Int. J. Mol. Sci. 2019, 20, 1453. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhang, H.; Yue, H.; Wu, S.; Yang, H.; Wang, Z.; Qi, G. Gas stunning with CO2 affected meat color, lipid peroxidation, oxidative stress, and gene expression of mitogen-activated protein kinases, glutathione S-transferases, and Cu/Zn-superoxide dismutase in the skeletal muscles of broilers. J. Anim. Sci. Biotechnol. 2018, 9, 37. [Google Scholar] [CrossRef]
- Xu, L.; Wang, J.; Yue, H.; Farnell, M.B.; Yang, H.; Wang, Z.; Qi, G. Evaluation of pre-slaughter low-current/high-frequency electrical stunning on lipid oxidative stability, antioxidant enzyme activity and gene expression of mitogen-activated protein kinase/nuclear factor erythroid 2-related factor 2 (MAPK/Nrf2) signalling pathway in thigh muscle of broilers. Int. J. Food Sci. Technol. 2020, 55, 953–960. [Google Scholar]
- Arbor Acres. Broiler Management Guide; Aviagen Limited: Midlothian, UK, 2009; pp. 1–59. [Google Scholar]
- Zhao, X.; Shi, W.; Yu, Z.; Wu, G.; Li, J.; Li, Y.; Zhang, J. Determination of Vitamin E in Feeds−High-Performance Liquid Chromatography. National Standard of the People’s Republic of China; GB/T 17812-2008; China Standards Press: Beijing, China, 2008. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sheehy, P.J.; Morrissey, P.A.; Flynn, A. Influence of dietary alpha-tocopherol on tocopherol concentrations in chick tissues. Br. Poult. Sci. 1991, 32, 391–397. [Google Scholar] [CrossRef]
- Pitargue, F.M.; Kim, J.H.; Goo, D.; Delos Reyes, J.B.; Kil, D.Y. Effect of vitamin E sources and inclusion levels in diets on growth performance, meat quality, alpha-tocopherol retention, and intestinal inflammatory cytokine expression in broiler chickens. Poult. Sci. 2019, 98, 4584–4594. [Google Scholar] [CrossRef]
- Mazur-Kuśnirek, M.; Antoszkiewicz, Z.; Lipiński, K.; Fijałkowska, M.; Purwin, C.; Kotlarczyk, S. The effect of polyphenols and vitamin E on the antioxidant status and meat quality of broiler chickens fed diets naturally contaminated with ochratoxin A. Arch. Anim. Nutr. 2019, 73, 431–444. [Google Scholar] [CrossRef]
- Voljc, M.; Frankic, T.; Levart, A.; Nemec, M.; Salobir, J. Evaluation of different vitamin E recommendations and bioactivity of α-tocopherol isomers in broiler nutrition by measuring oxidative stress in vivo and the oxidative stability of meat. Poult. Sci. 2011, 90, 1478–1488. [Google Scholar] [CrossRef]
- Vieira, V.; Marx, F.O.; Bassi, L.S.; Santos, M.C.; Oba, A.; de Oliveira, S.G.; Maiorka, A. Effect of age and different doses of dietary vitamin E on breast meat qualitative characteristics of finishing broilers. Anim. Nutr. 2021, 7, 163–167. [Google Scholar] [CrossRef]
- Mazur-Kuśnirek, M.; Antoszkiewicz, Z.; Lipiński, K.; Kaliniewicz, J.; Kotlarczyk, S.; Żukowski, P. The effect of polyphenols and vitamin E on the antioxidant status and meat quality of broiler chickens exposed to high temperature. Arch. Anim. Nutr. 2019, 73, 111–126. [Google Scholar] [CrossRef]
- Avila-Ramos, F.; Pro-Martínez, A.; Sosa-Montes, E.; Cuca-García, J.M.; Becerril-Pérez, C.M.; Figueroa-Velasco, J.L.; Narciso-Gaytán, C. Effects of dietary oregano essential oil and vitamin E on the lipid oxidation stability of cooked chicken breast meat. Poult. Sci. 2012, 91, 505–511. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Hager-Theodorides, A.L.; Massouras, T.; Simitzis, P.E.; Moschou, K.; Zoidis, E.; Sfakianaki, E.; Politi, K.; Charismiadou, M.; Goliomytis, M.; Deligeorgis, S. Hesperidin and naringin improve broiler meat fatty acid profile and modulate the expression of genes involved in fatty acid β-oxidation and antioxidant defense in a dose dependent manner. Foods 2021, 10, 739. [Google Scholar] [CrossRef]
- Khan, A.W.; Van den Berg, L. Some protein changes during post-mortem tenderization in poultry meat. J. Food Sci. 1964, 29, 597–601. [Google Scholar] [CrossRef]
- Obanu, Z.A.; Ledward, D.A.; Lawrie, R.A. The protein of intermediate moisture meat stored at tropical temperature: II. Effect of protein changes on some aspects of meat quality. Int. J. Food Sci. Technol. 1975, 10, 667–674. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Young, J.F.; Stagsted, J.; Jensen, S.K.; Karlsson, A.H.; Henckel, P. Ascorbic acid, alpha-tocopherol, and oregano supplements reduce stress-induced deterioration of chicken meat quality. Poult. Sci. 2003, 82, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Balah, A.; Ezzat, O.; Akool, E.S. Vitamin E inhibits cyclosporin A-induced CTGF and TIMP-1 expression by repressing ROS-mediated activation of TGF-β/Smad signaling pathway in rat liver. Int. Immunopharmacol. 2018, 65, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Oztürk-Urek, R.; Bozkaya, L.A.; Tarhan, L. The effects of some antioxidant vitamin- and trace element-supplemented diets on activities of SOD, CAT, GSH-Px and LPO levels in chicken tissues. Cell Biochem. Funct. 2001, 19, 125–132. [Google Scholar] [CrossRef]
- Alcala, M.; Calderon-Dominguez, M.; Serra, D.; Herrero, L.; Ramos, M.P.; Viana, M. Short-term vitamin E treatment impairs reactive oxygen species signaling required for adipose tissue expansion, resulting in fatty liver and insulin resistance in obese mice. PLoS ONE 2017, 12, e0186579. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Gupta, S. The multifaceted role of glutathione S-transferases in cancer. Cancer Lett. 2018, 433, 33–42. [Google Scholar] [CrossRef]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Fang, Y.; Cao, W.; Zhang, Z.; Pan, S.; Xu, X. Attenuation of tert-butyl hydroperoxide (t-BHP)-induced oxidative damage in HepG2 cells by tangeretin: Relevance of the Nrf2-ARE and MAPK signaling pathways. J. Agric. Food Chem. 2018, 66, 6317–6325. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013, 513, 1–13. [Google Scholar] [CrossRef]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.Á. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.D.; Strange, R.C. Invited commentary potential contribution of the glutathione s-transferase supergene family to resistance to oxidative stress. Free Radic. Res. 1995, 22, 193–207. [Google Scholar] [CrossRef]
- Sozen, E.; Karademir, B.; Yazgan, B.; Bozaykut, P.; Ozer, N.K. Potential role of proteasome on c-jun related signaling in hypercholesterolemia induced atherosclerosis. Redox Biol. 2014, 2, 732–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Abdelmegeed, M.A.; Song, B.J. Preventive effects of dietary walnuts on high-fat-induced hepatic fat accumulation, oxidative stress and apoptosis in mice. J. Nutr. Biochem. 2016, 38, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Minter, B.E.; Lowes, D.A.; Webster, N.R.; Galley, H.F. Differential effects of mitovite, α-tocopherol and trolox on oxidative stress, mitochondrial function and inflammatory signalling pathways in endothelial cells cultured under conditions mimicking sepsis. Antioxidants 2020, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.J.; Ryu, S.W.; Song, B.J. JNK- and p38 kinase-mediated phosphorylation of Bax leads to its activation and mitochondrial translocation and to apoptosis of human hepatoma HepG2 cells. J. Biol. Chem. 2006, 281, 21256–21265. [Google Scholar] [CrossRef] [Green Version]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Feed Composition | |
|---|---|---|
| d 0–17 | d 18–38 | |
| Corn | 63.00 | 55.90 |
| Soybean | 31.74 | 37.93 |
| Soybean oil | 1.82 | 2.23 |
| Limestone | 1.36 | 1.19 |
| Dicalcium phosphate | 1.30 | 1.87 |
| Lysine hydrochloride | 0.05 | 0.05 |
| DL-methionine | 0.07 | 0.17 |
| Salt | 0.35 | 0.35 |
| Choline | 0.23 | 0.23 |
| Vitamin premix 1 | 0.02 | 0.02 |
| Mineral premix 2 | 0.20 | 0.20 |
| Nutrient levels 3 | ||
| Metabolizable energy, MJ/kg | 12.22 | 12.39 |
| Crude protein, % | 22.60 | 20.30 |
| Lysine, % | 1.24 | 1.08 |
| Methionine, % | 0.48 | 0.36 |
| Methionine + Cystine, % | 0.82 | 0.68 |
| Calcium, % | 0.94 | 0.86 |
| Non-phytate phosphorus, % | 0.43 | 0.34 |
| Treatments 1 | Dietary VE Content (IU/kg) | VE Feeding Duration | Electrical Stunning (Alternative Current) | |||
|---|---|---|---|---|---|---|
| d 0–17 | d 18–24 | d 25–31 | d 32–38 | |||
| Control | 20 | 0 | 0 | 0 | None | 130 mA, 50 Hz, 1 s |
| W1 | 20 | 0 | 0 | 200 | One week | 130 mA, 50 Hz, 1 s |
| W2 | 20 | 0 | 200 | 200 | Two weeks | 130 mA, 50 Hz, 1 s |
| W3 | 20 | 200 | 200 | 200 | Three weeks | 130 mA, 50 Hz, 1 s |
| Items 1 | Variables 2 | VE Retention | |
|---|---|---|---|
| Breast Muscle | Thigh Muscle | ||
| VE | Feeding duration | 0.94 ** | 0.92 ** |
| TBARSd0 | −0.53 ** | −0.42 * | |
| TBARS | TBARSd2 | −0.72 ** | −0.85 ** |
| TBARSd4 | −0.33 | −0.44 * | |
| TBARSd6 | −0.60 ** | −0.51 * | |
| SODd0 | −0.04 | −0.14 | |
| Enzyme activity | SDDd2 | −0.07 | −0.07 |
| GSTd0 | 0.22 | −0.72 ** | |
| GSTd2 | 0.40 * | 0.58 ** | |
| MAPKs genes | JNK1 | −0.49 * | −0.43 * |
| JNK2 | −0.52 ** | −0.23 | |
| p38 | 0.01 | −0.44 * | |
| Nrf2 gene | Nrf2 | −0.45 * | 0.03 |
| Antioxidant genes | SOD1 | −0.10 | −0.25 |
| SOD2 | −0.16 | 0.06 | |
| GSTA3 | −0.21 | −0.14 | |
| GSTT1 | −0.30 | 0.11 | |
| Tissues | Items 1 | Variables 2 | TBARS | |||
|---|---|---|---|---|---|---|
| TBARSd0 | TBARSd2 | TBARSd4 | TBARSd6 | |||
| Breast muscle | VE | Feeding duration | −0.56 ** | −0.76 ** | −0.37 | −0.68 ** |
| Enzyme activity | SODd0 | −0.15 | −0.07 | −0.19 | 0.24 | |
| SDDd2 | −0.07 | −0.13 | −0.09 | 0.17 | ||
| GSTd0 | −0.19 | −0.42 * | −0.26 | −0.14 | ||
| GSTd2 | −0.21 | −0.51 ** | 0.12 | −0.20 | ||
| MAPKs genes | JNK1 | 0.02 | 0.29 | 0.35 | 0.09 | |
| JNK2 | 0.01 | 0.65 ** | 0.31 | 0.06 | ||
| p38 | 0.02 | 0.10 | 0.33 | 0.10 | ||
| Nrf2 gene | Nrf2 | 0.11 | 0.45 * | 0.29 | 0.10 | |
| Antioxidant genes | SOD1 | −0.15 | 0.07 | 0.17 | 0.05 | |
| SOD2 | −0.06 | 0.15 | 0.15 | −0.14 | ||
| GSTA3 | 0.03 | −0.06 | 0.25 | −0.09 | ||
| GSTT1 | 0.30 | 0.37 | 0.21 | 0.38 | ||
| Thigh muscle | VE | Feeding duration | −0.62 ** | −0.92 ** | −0.59 ** | −0.57 ** |
| Enzyme activity | SODd0 | 0.33 | 0.22 | 0.23 | 0.09 | |
| SDDd2 | 0.47 * | 0.24 | 0.44 * | 0.05 | ||
| GSTd0 | 0.22 | 0.51 ** | 0.09 | 0.27 | ||
| GSTd2 | −0.16 | −0.78 ** | −0.34 | −0.27 | ||
| JNK1 | 0.11 | 0.25 | 0.22 | 0.49 * | ||
| MAPKs genes | JNK2 | 0.11 | 0.12 | 0.34 | 0.28 | |
| p38 | 0.27 | 0.28 | 0.12 | 0.30 | ||
| Nrf2 | 0.01 | −0.22 | −0.20 | 0.06 | ||
| Nrf2 gene | SOD1 | 0.03 | 0.05 | 0.36 | 0.30 | |
| Antioxidant genes | SOD2 | 0.02 | −0.34 | −0.13 | 0.05 | |
| GSTA3 | 0.14 | 0.01 | 0.31 | 0.45 * | ||
| GSTT1 | 0.27 | −0.18 | 0.15 | 0.25 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Wang, J.; Zhang, H.; Wu, S.; Yue, H.; Wan, X.; Yang, H.; Wang, Z.; Qi, G. Vitamin E Supplementation Enhances Lipid Oxidative Stability via Increasing Vitamin E Retention, Rather Than Gene Expression of MAPK-Nrf2 Signaling Pathway in Muscles of Broilers. Foods 2021, 10, 2555. https://doi.org/10.3390/foods10112555
Xu L, Wang J, Zhang H, Wu S, Yue H, Wan X, Yang H, Wang Z, Qi G. Vitamin E Supplementation Enhances Lipid Oxidative Stability via Increasing Vitamin E Retention, Rather Than Gene Expression of MAPK-Nrf2 Signaling Pathway in Muscles of Broilers. Foods. 2021; 10(11):2555. https://doi.org/10.3390/foods10112555
Chicago/Turabian StyleXu, Lei, Jing Wang, Haijun Zhang, Shugeng Wu, Hongyuan Yue, Xiaoli Wan, Haiming Yang, Zhiyue Wang, and Guanghai Qi. 2021. "Vitamin E Supplementation Enhances Lipid Oxidative Stability via Increasing Vitamin E Retention, Rather Than Gene Expression of MAPK-Nrf2 Signaling Pathway in Muscles of Broilers" Foods 10, no. 11: 2555. https://doi.org/10.3390/foods10112555
APA StyleXu, L., Wang, J., Zhang, H., Wu, S., Yue, H., Wan, X., Yang, H., Wang, Z., & Qi, G. (2021). Vitamin E Supplementation Enhances Lipid Oxidative Stability via Increasing Vitamin E Retention, Rather Than Gene Expression of MAPK-Nrf2 Signaling Pathway in Muscles of Broilers. Foods, 10(11), 2555. https://doi.org/10.3390/foods10112555