Flexible Polymer-Based Electrodes for Detecting Depression-Related Theta Oscillations in the Medial Prefrontal Cortex


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
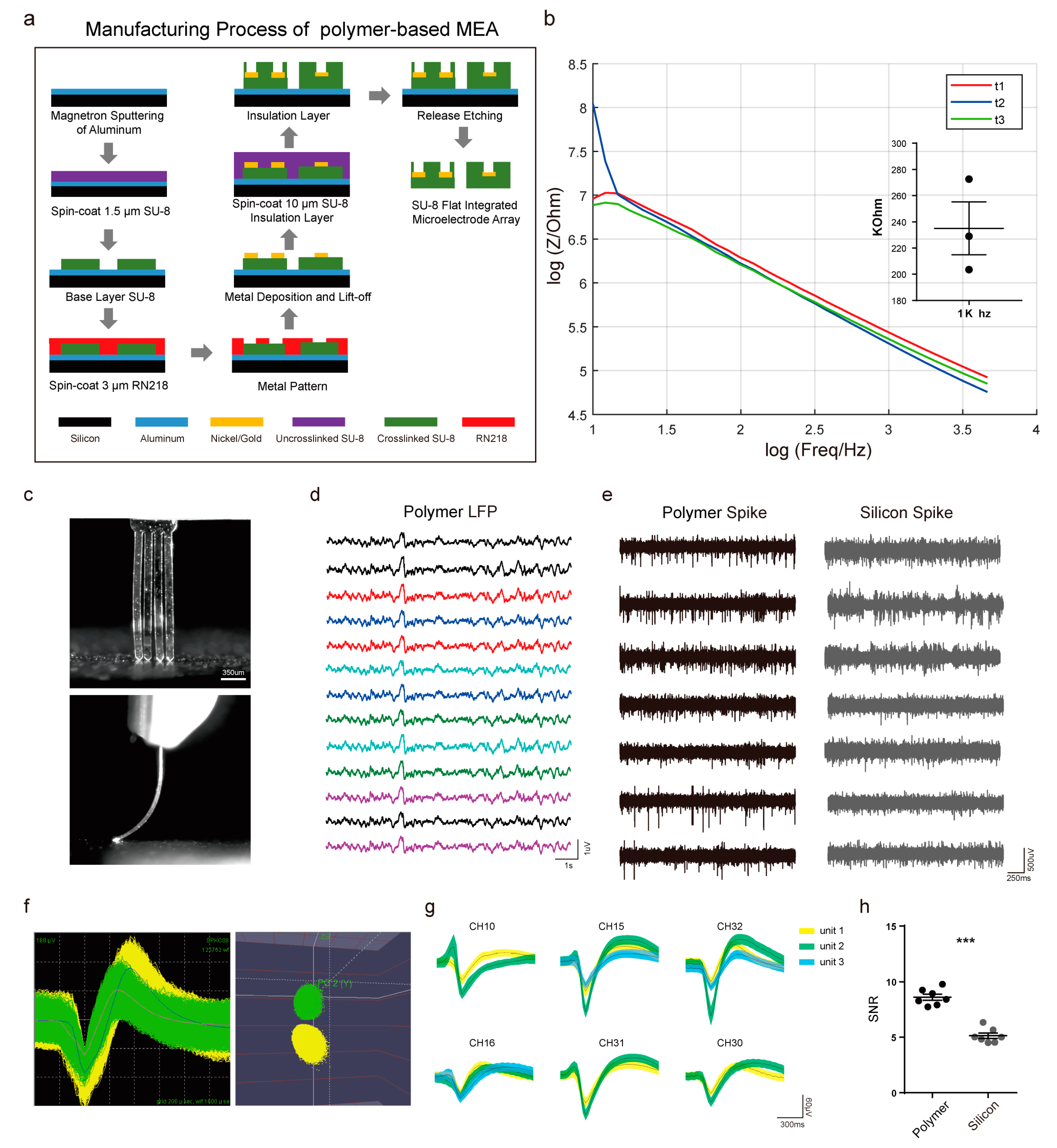
2.2. Electrode Fabrication and EIS Testing
2.3. In Vivo Electrophysiology
2.3.1. Electrode Implantation Surgery
2.3.2. Signal-to-Noise Ratio (SNR) Calculation
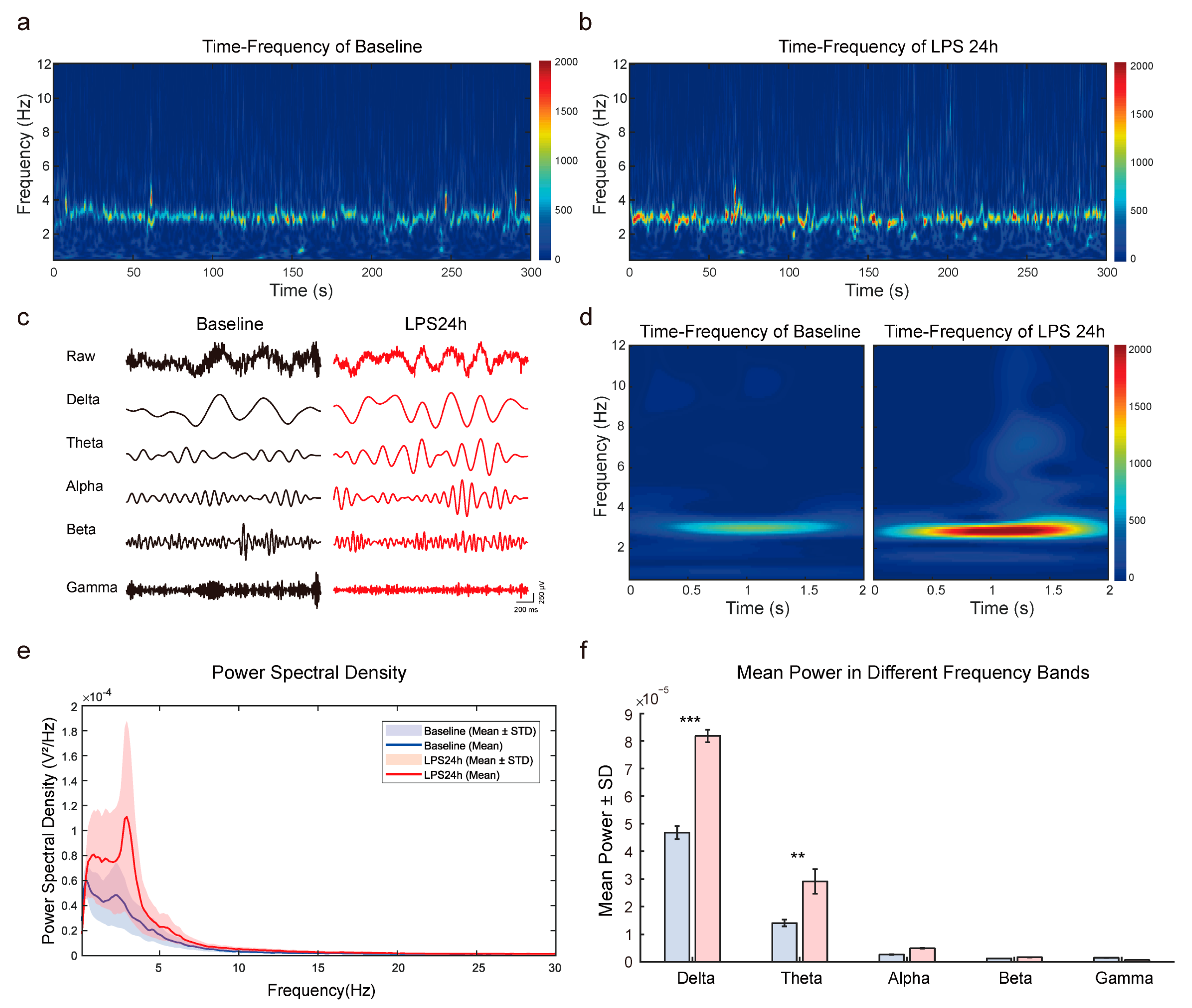
2.3.3. Power Spectral Density (PSD) Analysis
2.3.4. Time–Frequency Analysis
2.4. Behavioral Assays
2.4.1. Open Field Test (OFT)
2.4.2. Elevated Plus Maze Test (EPM)
2.4.3. Tail-Suspension Test (TST)
2.4.4. Forced Swim Test (FST)
2.4.5. Sucrose Preference Test (SPT)
2.5. Statistics and Data Visualization
3. Results
3.1. Behavioral Validation of the LPS-Induced Acute Depression Model in Mice
3.2. Fabrication and Performance Evaluation of Flexible Polymer Multichannel Electrodes
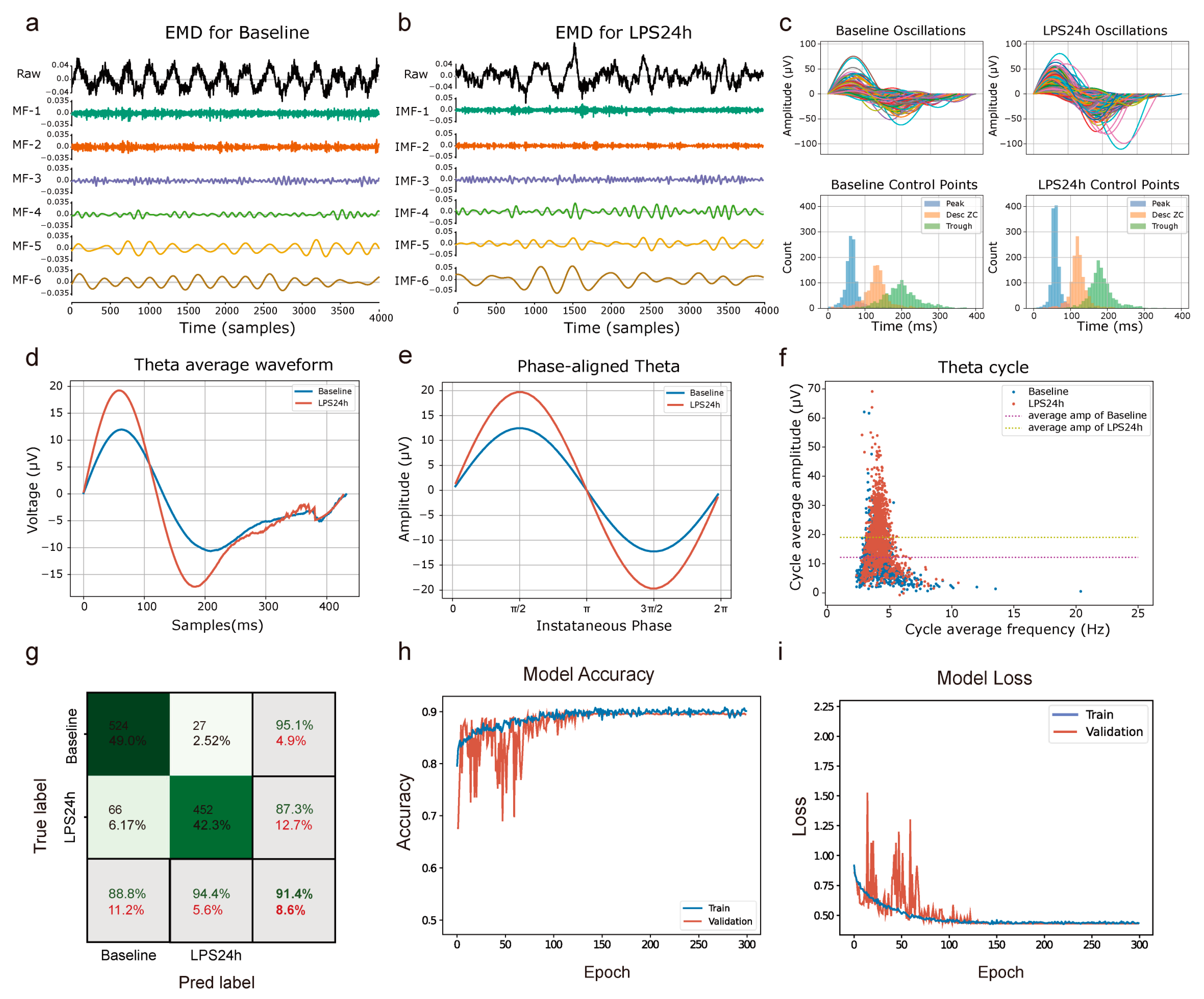
3.3. Enhanced Theta Oscillations in the mPFC of LPS-Induced Depressive Mice
3.4. Depression State Recognition Based on EMD and CNN-LSTM Machine Learning Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef] [PubMed]
- Marwaha, S.; Palmer, E.; Suppes, T.; Cons, E.; Young, A.H.; Upthegrove, R. Novel and emerging treatments for major depression. Lancet 2023, 401, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qiao, Y.; Wang, M.; Liang, X.; Zhang, M.; Li, C.; Cairang, J.; Wang, J.; Bi, H.; Gao, T. The influence of genetic and acquired factors on the vulnerability to develop depression: A review. Biosci. Rep. 2023, 43, BSR20222644. [Google Scholar] [CrossRef] [PubMed]
- Smart, O.L.; Tiruvadi, V.R.; Mayberg, H.S. Multimodal approaches to define network oscillations in depression. Biol. Psychiatry 2015, 77, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zhang, T. Alteration of phase-phase coupling between theta and gamma rhythms in a depression-model of rats. Cogn. Neurodyn. 2013, 7, 167–172. [Google Scholar] [CrossRef]
- Hare, B.D.; Duman, R.S. Prefrontal cortex circuits in depression and anxiety: Contribution of discrete neuronal populations and target regions. Mol. Psychiatry 2020, 25, 2742–2758. [Google Scholar] [CrossRef]
- Csicsvari, J.; Hirase, H.; Czurkó, A.; Mamiya, A.; Buzsáki, G. Oscillatory coupling of hippocampal pyramidal cells and interneurons in the behaving Rat. J. Neurosci. 1999, 19, 274–287. [Google Scholar] [CrossRef]
- Kumar, S.; Black, S.J.; Hultman, R.; Szabo, S.T.; DeMaio, K.D.; Du, J.; Katz, B.M.; Feng, G.; Herbert, E.C., III; Dzirasa, K. Cortical control of affective networks. J. Neurosci. 2013, 33, 1116–1129. [Google Scholar] [CrossRef]
- Sun, Y.; Giacobbe, P.; Tang, C.W.; Barr, M.S.; Rajji, T.; Kennedy, S.H.; Fitzgerald, P.B.; Lozano, A.M.; Wong, W.; Daskalakis, Z.J. Deep Brain Stimulation Modulates Gamma Oscillations and Theta-Gamma Coupling in Treatment Resistant Depression. Brain Stimul. 2015, 8, 1033–1042. [Google Scholar] [CrossRef]
- Dantzer, R.; O’connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Lawson, M.A.; Andre, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol. Psychiatry 2009, 14, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Normann, R.A.; Fernandez, E. Clinical applications of penetrating neural interfaces and Utah Electrode Array technologies. J. Neural Eng. 2016, 13, 061003. [Google Scholar] [CrossRef] [PubMed]
- Kipke, D.R.; Shain, W.; Buzsáki, G.; Fetz, E.; Henderson, J.M.; Hetke, J.F.; Schalk, G. Advanced Neurotechnologies for Chronic Neural Interfaces: New Horizons and Clinical Opportunities. J. Neurosci. 2008, 28, 11830–11838. [Google Scholar] [CrossRef] [PubMed]
- Zorzos, A.N.; Boyden, E.S.; Fonstad, C.G. Multiwaveguide implantable probe for light delivery to sets of distributed brain targets. Opt. Lett. 2010, 35, 4133–4135. [Google Scholar] [CrossRef]
- Jeong, J.W.; Yeo, W.H.; Akhtar, A.; Norton, J.J.; Kwack, Y.J.; Li, S.; Jung, S.Y.; Su, Y.; Lee, W.; Xia, J.; et al. Materials and optimized designs for human-machine interfaces via epidermal electronics. Adv. Mater. 2013, 25, 6839–6846. [Google Scholar] [CrossRef]
- Kim, D.H.; Viventi, J.; Amsden, J.J.; Xiao, J.; Vigeland, L.; Kim, Y.S.; Blanco, J.A.; Panilaitis, B.; Frechette, E.S.; Contreras, D.; et al. Dissolvable films of silk fibroin for ultrathin conformal bio-integrated electronics. Nat. Mater. 2010, 9, 511–517. [Google Scholar] [CrossRef]
- Luan, L.; Wei, X.; Zhao, Z.; Siegel, J.J.; Potnis, O.; Tuppen, C.A.; Lin, S.; Kazmi, S.; Fowler, R.A.; Holloway, S.; et al. Ultraflexible nanoelectronic probes form reliable, glial scar-free neural integration. Sci. Adv. 2017, 3, e1601966. [Google Scholar] [CrossRef]
- Kozai TD, Y.; Langhals, N.B.; Patel, P.R.; Deng, X.; Zhang, H.; Smith, K.L.; Lahann, J.; Kotov, N.A.; Kipke, D. Ultrasmall implantable composite microelectrodes with bioactive surfaces for chronic neural interfaces. Nat. Mater 2012, 11, 1065–1073. [Google Scholar] [CrossRef]
- Quinn, A.J.; Lopes-dos-Santos, V.; Dupret, D.; Nobre, A.C.; Woolrich, M.W. EMD: Empirical Mode Decomposition and Hilbert-Huang Spectral Analyses in Python. J. Open Source Softw. 2021, 6, 2977. [Google Scholar] [CrossRef]
- Okonogi, T.; Sasaki, T. Theta-Range Oscillations in Stress-Induced Mental Disorders as an Oscillotherapeutic Target. Front. Behav. Neurosci. 2021, 15, 698753. [Google Scholar] [CrossRef]
- Ward, M.P.; Rajdev, P.; Ellison, C.; Irazoqui, P.P. Toward a comparison of microelectrodes for acute and chronic recordings. Brain Res. 2009, 1282, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Guo, L. (Ed.) Recording Electrodes. In Principles of Electrical Neural Interfacing: A Quantitative Approach to Cellular Recording and Stimulation; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; pp. 17–31. [Google Scholar]
- Neto, J.P.; Baião, P.; Lopes, G.; Frazão, J.; Nogueira, J.; Fortunato, E.; Barquinha, P.; Kampff, A.R. Does Impedance Matter When Recording Spikes With Polytrodes? Front. Neurosci. 2018, 12, 715. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y. Material considerations for in vitro neural interface technology. MRS Bull. 2012, 37, 566–572. [Google Scholar] [CrossRef]
- Sun, R.; Tang, M.Y.; Yang, D.; Zhang, Y.Y.; Xu, Y.H.; Qiao, Y.; Yu, B.; Cao, S.X.; Wang, H.; Huang, H.Q. C3aR in the medial prefrontal cortex modulates the susceptibility to LPS-induced depressive-like behaviors through glutamatergic neuronal excitability. Prog. Neurobiol. 2024, 236, 102614. [Google Scholar] [CrossRef] [PubMed]
- Rho, Y.-A.; Sherfey, J.; Vijayan, S. Emotional Memory Processing during REM Sleep with Implications for Post-Traumatic Stress Disorder. J. Neurosci. 2023, 43, 433–446. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, W.; Liu, S.; Su, F.; Kang, X.; Ke, Y.; Ming, D. Altered fronto-central theta-gamma coupling in major depressive disorder during auditory steady-state responses. Clin. Neurophysiol. 2023, 146, 65–76. [Google Scholar] [CrossRef]
- Porsolt, R.D. Behavioral Despair Revisited—A Citation-Classic Commentary on Depression—A New Animal-Model Sensitive to Antidepressant Treatments; Porsolt, R.D., Lepichon, M., Jalfre, M., Eds.; Institute For Scientific Information: Philadelphia, PA, USA, 1993; Volume 20, p. 9. [Google Scholar]
- Khodagholy, D.; Gelinas, J.N.; Thesen, T.; Doyle, W.; Devinsky, O.; Malliaras, G.G.; Buzsáki, G. NeuroGrid: Recording action potentials from the surface of the brain. Nat. Neurosci. 2015, 18, 310–315. [Google Scholar] [CrossRef]
- Mitchell, D.J.; McNaughton, N.; Flanagan, D.; Kirk, I.J. Frontal-midline theta from the perspective of hippocampal “theta”. Prog. Neurobiol. 2008, 86, 156–185. [Google Scholar] [CrossRef]
- Knyazev, G.G. Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci. Biobehav. Rev. 2007, 31, 377–395. [Google Scholar] [CrossRef]
- Olbrich, S.; Arns, M. EEG biomarkers in major depressive disorder: Discriminative power and prediction of treatment response. Int. Rev. Psychiatry 2013, 25, 604–618. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Redmond, S.J.; Bertoux, M.; Hodges, J.R.; Hornberger, M. A Comparison of Magnetic Resonance Imaging and Neuropsychological Examination in the Diagnostic Distinction of Alzheimer’s Disease and Behavioral Variant Frontotemporal Dementia. Front. Aging Neurosci. 2016, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Canolty, R.T.; Knight, R.T. The functional role of cross-frequency coupling. Trends Cogn. Sci. 2010, 14, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Bland, B.H.; Oddie, S.D. Theta band oscillation and synchrony in the hippocampal formation and associated structures: The case for its role in sensorimotor integration. Behav. Brain Res. 2001, 127, 119–136. [Google Scholar] [CrossRef]
- Jacobs, J.; Hwang, G.; Curran, T.; Kahana, M.J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. Neuroimage 2006, 32, 978–987. [Google Scholar] [CrossRef]
- Fell, J.; Axmacher, N. The role of phase synchronization in memory processes. Nat. Rev. Neurosci. 2011, 12, 105–118. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, R.; Shang, S.; Yuan, Q.; Wang, P.; Zhuang, L. Flexible Polymer-Based Electrodes for Detecting Depression-Related Theta Oscillations in the Medial Prefrontal Cortex. Chemosensors 2024, 12, 258. https://doi.org/10.3390/chemosensors12120258
Sun R, Shang S, Yuan Q, Wang P, Zhuang L. Flexible Polymer-Based Electrodes for Detecting Depression-Related Theta Oscillations in the Medial Prefrontal Cortex. Chemosensors. 2024; 12(12):258. https://doi.org/10.3390/chemosensors12120258
Chicago/Turabian StyleSun, Rui, Shunuo Shang, Qunchen Yuan, Ping Wang, and Liujing Zhuang. 2024. "Flexible Polymer-Based Electrodes for Detecting Depression-Related Theta Oscillations in the Medial Prefrontal Cortex" Chemosensors 12, no. 12: 258. https://doi.org/10.3390/chemosensors12120258
APA StyleSun, R., Shang, S., Yuan, Q., Wang, P., & Zhuang, L. (2024). Flexible Polymer-Based Electrodes for Detecting Depression-Related Theta Oscillations in the Medial Prefrontal Cortex. Chemosensors, 12(12), 258. https://doi.org/10.3390/chemosensors12120258