Recent Advances of In Vitro Culture for the Application of New Breeding Techniques in Citrus

 ,
, 
 ,
,  ,
, 
Abstract
1. Introduction
2. Regeneration of Citrus for Genetic Transformation
2.1. Genotype Influence
2.2. Source of Explant Type
2.3. Basal Media and Other Factors Influencing Organogenic Response
3. Citrus Transformation Protocols
3.1. Selectable Marker Strategy
3.2. Role of the Promoter
4. Attempts to Reduce the Long Juvenility
5. Success in Transgenesis Applied to Citrus
6. Genome Editing
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT Database Results. 2018. Available online: http://www.fao.org/faostat/ (accessed on 5 April 2020).
- Eriksson, D.; Harwood, W.; Hofvander, P.; Jones, H.; Rogowsky, P.; Stöger, E.; Visser, R.G.F. A Welcome Proposal to Amend the GMO Legislation of the EU. Trends Biotechnol. 2018, 36, 1100–1103. [Google Scholar] [CrossRef]
- Limera, C.; Sabbadini, S.; Sweet, J.B.; Mezzetti, B. New Biotechnological Tools for the Genetic Improvement of Major Woody Fruit Species. Front. Plant Sci. 2017, 8, 1418. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xie, C.; Si, H.; Yang, J. CRISPR/Cas9-mediated genome editing in plants. Methods 2017, 121, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Cisgenic plants are similar to traditionally bred plants: International regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep. 2006, 7, 750–753. [Google Scholar] [CrossRef]
- Lusser, M.; Parisi, C.; Plan, D.; Rodríguez-Cerezo, E. Deployment of new biotechnologies in plant breeding. Nat. Biotechnol. 2012, 30, 231–239. [Google Scholar] [CrossRef]
- De Alba, A.E.M.; Elvira-Matelot, E.; Vaucheret, H. Gene silencing in plants: A diversity of pathways. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 1300–1308. [Google Scholar] [CrossRef]
- Chhetri, A.; Hazarika, B.N.; Angami, T.; Devi, H.; Alice, A.; Khonglah, L. Overview and Applications of RNA Silencing in Fruit Crops. Curr. J. Appl. Sci. Technol. 2019, 33, 1–7. [Google Scholar] [CrossRef]
- Grosser, J.W.; Ollitrault, P.; Olivares-Fuster, O. Invited review: Somatic hybridization in citrus: An effective tool to facilitate variety improvement. Vitr. Cell. Dev. Biol. Plant 2000, 36, 434–449. [Google Scholar] [CrossRef]
- Dutt, M.; Erpen, L.; Grosser, J.W. Genetic transformation of the ‘W Murcott’ tangor: Comparison between different techniques. Sci. Hortic. 2018, 242, 90–94. [Google Scholar] [CrossRef]
- Li, D.D.; Shi, W.; Deng, X.X. Agrobacterium-mediated transformation of embryogenic calluses of Ponkan mandarin and the regeneration of plants containing the chimeric ribonuclease gene. Plant Cell Rep. 2002, 21, 153–156. [Google Scholar] [CrossRef]
- Dutt, M.; Vasconcellos, M.; Song, K.J.; Gmitter, F.G.; Grosser, J.W. In vitro production of autotetraploid Ponkan mandarin (Citrus reticulata Blanco) using cell suspension cultures. Euphytica 2010, 173, 235–242. [Google Scholar] [CrossRef]
- Omar, A.A.; Dutt, M.; Gmitter, F.G.; Grosser, J.W. Chapter 13 Somatic Embryogenesis: Still a Relevant Technique in Citrus Improvement. In In Vitro Embryogenesis in Higher Plants, Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; Volume 1359, ISBN 9781493930616. [Google Scholar]
- Orbović, V.; Grosser, J.W. Citrus Transformation Using Juvenile Tissue Explants. In Agrobacterium Protocols: Vol. 2, Methods in Molecular Biology (Clifton, N.J.); Wang, K., Ed.; Springer: New York, NY, USA, 2015; pp. 245–258. ISBN 9781493916580. [Google Scholar]
- Pena, L.; Perez, R.M.; Cervera, M.; Juarez, J.A.; Navarro, L. Agrobacterium-mediated Transformation of Citrus. In Transgenic Crops of the World; Curtis, I.S., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 145–157. [Google Scholar]
- Dutt, M.; Grosser, J.W. Evaluation of parameters affecting Agrobacterium-mediated transformation of citrus. Plant Cell Tissue Organ Cult. 2009, 98, 331–340. [Google Scholar] [CrossRef]
- Peña, L.; Cervera, M.; Juárez, J.; Ortega, C.; Pina, J.A.; Durán-Vila, N.; Navarro, L. High efficiency Agrobacterium-mediated transformation and regeneration of citrus. Plant Sci. 1995, 104, 183–191. [Google Scholar] [CrossRef]
- Cervera, M.; Navarro, A.; Navarro, L.; Peña, L. Production of transgenic adult plants from clementine mandarin by enhancing cell competence for transformation and regeneration. Tree Physiol. 2008, 28, 55–66. [Google Scholar] [CrossRef]
- Ghorbel, R.; Dominguez, A.; Navarro, L.; Pena, L. High efficiency genetic transformation of sour orange (Citrus aurantium) and production of transgenic trees containing the coat protein gene of citrus tristeza virus. Tree Physiol. 2000, 20, 1183–1189. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Boscariol, R.L.; Almeida, W.A.B.; Derbyshire, M.T.V.C.; Mourão Filho, F.A.A.; Mendes, B.M.J. The use of the PMI/mannose selection system to recover transgenic sweet orange plants (Citrus sinensis L. Osbeck). Plant Cell Rep. 2003, 22, 122–128. [Google Scholar] [CrossRef]
- Marutani-Hert, M.; Bowman, K.D.; McCollum, G.T.; Mirkov, T.E.; Evens, T.J.; Niedz, R.P. A Dark Incubation Period Is Important for Agrobacterium-Mediated Transformation of Mature Internode Explants of Sweet Orange, Grapefruit, Citron, and a Citrange Rootstock. PLoS ONE 2012, 7, e47426. [Google Scholar] [CrossRef]
- Peng, A.; Zou, X.; Xu, L.; He, Y.; Lei, T.; Yao, L.; Li, Q.; Chen, S. Improved protocol for the transformation of adult Citrus sinensis Osbeck ‘Tarocco’ blood orange tissues. Vitr. Cell. Dev. Biol. Plant 2019, 55, 659–667. [Google Scholar] [CrossRef]
- Cervera, M.; Juárez, J.; Navarro, A.; Pina, J.A.; Durán-Vila, N.; Navarro, L.; Peña, L. Genetic transformation and regeneration of mature tissues of woody fruit plants bypassing the juvenile stage. Transgenic Res. 1998, 7, 51–59. [Google Scholar] [CrossRef]
- Zou, X.; Peng, A.; Xu, L.; Liu, X.; Lei, T.; Yao, L.; He, Y.; Chen, S. Efficient auto-excision of a selectable marker gene from transgenic citrus by combining the Cre/loxP system and ipt selection. Plant Cell Rep. 2013, 32, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xu, C.J.; Hu, G.B.; Chen, K.S. Establishment of an Agrobacterium-mediated transformation system for Fortunella crassifolia. Biol. Plant. 2007, 51, 541–545. [Google Scholar] [CrossRef]
- Tong, Z.; Tan, B.; Zhang, J.; Hu, Z.; Guo, W.; Deng, X. Using precocious trifoliate orange (Poncirus trifoliata [L.] Raf.) to establish a short juvenile transformation platform for citrus. Sci. Hortic. 2009, 119, 335–338. [Google Scholar] [CrossRef]
- Singh, S.; Rajam, M.V. Highly Efficient and Rapid Plant Regeneration in Citrus sinensis. J. Plant Biochem. Biotechnol. 2010, 19, 195–202. [Google Scholar] [CrossRef]
- Almeida, W.A.B.; Mourão Filho, F.A.A.; Pino, L.E.; Boscariol, R.L.; Rodriguez, A.P.M.; Mendes, B.M.J. Genetic transformation and plant recovery from mature tissues of Citrus sinensis L. Osbeck. Plant Sci. 2003, 164, 203–211. [Google Scholar] [CrossRef]
- Cervera, M.; Juárez, J.; Navarro, L.; Peña, L. Genetic Transformation of Mature Citrus Plants. In Transgenic Plants. Methods and Protocols; Humana Press: Totowa, NJ, USA, 2005; Volume 286, pp. 177–188. [Google Scholar]
- Kobayashi, A.K.; Vieira, L.G.E.; Bespalhok Filho, J.C.; Leite, R.P.; Pereira, L.F.P.; Molinari, H.B.C.; Marques, V.V. Enhanced resistance to citrus canker in transgenic sweet orange expressing the sarcotoxin IA gene. Eur. J. Plant Pathol. 2017, 149, 865–873. [Google Scholar] [CrossRef]
- Kobayashi, A.K.; Bespalhok, J.C.; Pereira, L.F.P.; Vieira, L.G.E. Plant regeneration of sweet orange (Citrus sinensis) from thin sections of mature stem segments. Plant Cell Tissue Organ Cult. 2003, 74, 99–102. [Google Scholar] [CrossRef]
- Esmaeilnia, E.; Dehestani, A. In vitro plant regeneration from mature tissues of Thomson navel sweet orange (Citrus sinensis L. Osbeck.). Biharean Biol. 2015, 9, 9–14. [Google Scholar]
- Khan, E.U.; Fu, X.; Wang, J.; Fan, Q.; Huang, X.; Zhang, G.; Shi, J.; Liu, J. Regeneration and characterization of plants derived from leaf in vitro culture of two sweet orange (Citrus sinensis (L.) Osbeck) cultivars. Sci. Hortic. 2009, 120, 70–76. [Google Scholar] [CrossRef]
- Khan, E.U.; Fu, X.Z.; Liu, J.H. Agrobacterium-mediated genetic transformation and regeneration of transgenic plants using leaf segments as explants in Valencia sweet orange. Plant Cell Tissue Organ Cult. 2012, 109, 383–390. [Google Scholar] [CrossRef]
- Dutt, M.; Grosser, J.W. An embryogenic suspension cell culture system for Agrobacterium-mediated transformation of citrus. Plant Cell Rep. 2010, 29, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Grosser, J.W.; Gmitter, F.G. Protoplast fusion and citrus improvement. Plant Breed Rev. 1990, 8, 339–374. [Google Scholar]
- Omar, A.A.; Song, W.Y.; Grosser, J.W. Introduction of Xa21, a Xanthomonas-resistance, gene from rice, into “Hamlin” sweet orange [Citrus sinensis (L.) Osbeck] using protoplast-GFP co-transformation or single plasmid transformation. J. Hortic. Sci. Biotechnol. 2007, 82, 914–923. [Google Scholar] [CrossRef]
- Guo, W.; Duan, Y.; Olivares-Fuster, O.; Wu, Z.; Arias, C.R.; Burns, J.K.; Grosser, J.W. Protoplast transformation and regeneration of transgenic Valencia sweet orange plants containing a juice quality-related pectin methylesterase gene. Plant Cell Rep. 2005, 24, 482–486. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Murashige, T.; Tucker, D.P.H. Growth factor requirements of citrus tissue culture. Proc. First Intl. Citrus Symp. 1969, 3, 1155–1161. [Google Scholar]
- de Oliveira, M.P.; Cardoso Costa, M.; da Silva, C.; Campos Otoni, W. Growth regulators, culture media and antibiotics in the in vitro shoot regeneration from mature tissue of citrus cultivars. Pesquisa Agropecuária Brasileira 2010, 45, 654–660. [Google Scholar] [CrossRef]
- Cervera, M.; Pina, J.A.; Juárez, J.; Navarro, L.; Peña, L. Agrobacterium-mediated transformation of citrange: Factors affecting transformation and regeneration. Plant Cell Rep. 1998, 18, 271–278. [Google Scholar] [CrossRef]
- Boscariol, R.L.; Monteiro, M.; Takahashi, E.K.; Chabregas, S.M.; Vieira, M.L.C.; Vieira, L.G.E.; Pereira, L.F.P.; Mouräo Filho, F.D.A.A.; Cardoso, S.C.; Christiano, R.S.C.; et al. Attacin A gene from Tricloplusia ni reduces susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinesis “Hamlin”. J. Am. Soc. Hortic. Sci. 2006, 131, 530–536. [Google Scholar] [CrossRef]
- Cardoso, S.C.; Barbosa-Mendes, J.M.; Boscariol-Camargo, R.L.; Christiano, R.S.C.; Filho, A.B.; Vieira, M.L.C.; Mendes, B.M.J.; Mourão Filho, F.D.A.A. Transgenic sweet orange (Citrus sinensis L. Osbeck) expressing the attacin a gene for resistance to Xanthomonas citri subsp. citri. Plant Mol. Biol. Rep. 2010, 28, 185–192. [Google Scholar] [CrossRef]
- Lyoyd, G.B.; McCown, B.H. Commercially feasible micropropagation of mountain laurel (Kalmia latifolia) by use of shoot tip culture. Proc. Int. Plant Propag. Soc. 1980, 30, 421–437. [Google Scholar]
- Rodríguez, A.; Cervera, M.; Peris, J.E.; Peña, L. The same treatment for transgenic shoot regeneration elicits the opposite effect in mature explants from two closely related sweet orange (Citrus sinensis (L.) Osb.) genotypes. Plant Cell Tissue Organ Cult. 2008, 93, 97–106. [Google Scholar] [CrossRef]
- Peña, L.; Cervera, M.; Fagoaga, C.; Perez, R.; Romero, J.; Juarez, J.; Pina, J.A.; Navarro, L. Agrobacterium-Mediated Transformation of Citrus. In Transgenic Crops of the World; Springer: Dordrecht, The Netherlands, 2004; pp. 145–158. ISBN 9789401570213. [Google Scholar]
- Navarro-García, N.; Morte, M.; Pérez-Tornero, O. In vitro adventitious organogenesis and histological characterization from mature nodal explants of Citrus limon. Vitr. Cell. Dev. Biol. Plant 2016, 52, 161–173. [Google Scholar] [CrossRef]
- Dutt, M.; Vasconcellos, M.; Grosser, J.W. Effects of antioxidants on Agrobacterium-mediated transformation and accelerated production of transgenic plants of Mexican lime (Citrus aurantifolia Swingle). Plant Cell Tissue Organ Cult. 2011, 107, 79–89. [Google Scholar] [CrossRef]
- Duran-Vila, N.; Gogorcena, Y.; Ortega, V.; Ortiz, J.; Navarro, L. Morphogenesis and tissue culture of sweet orange (Citrus sinensis (L.) Osb.): Effect of temperature and photosynthetic radiation. Plant Cell Tissue Organ Cult. 1992, 29, 11–18. [Google Scholar] [CrossRef]
- Kobayashi, S.; Uchimiya, H. Expression and integration of a foreign gene in orange (Citrus sinensis Osb.) protoplasts by direct DNA transfer. Jpn. J. Genet. 1989, 64, 91–97. [Google Scholar] [CrossRef]
- Gong, X.Q.; Liu, J.H. Genetic transformation and genes for resistance to abiotic and biotic stresses in Citrus and its related genera. Plant Cell Tissue Organ Cult. 2013, 113, 137–147. [Google Scholar] [CrossRef]
- Orbović, V.; Shankar, A.; Peeples, M.; Hubbard, C.; Zale, J. Citrus Transformation Using Mature Tissue Explants. In Agrobacterium Protocols: Vol. 2, Methods in Molecular Biology (Clifton, N.J.); Wang, K., Ed.; Springer: New York, NY, USA, 2015; Volume 1224, pp. 259–274. ISBN 9781493916580. [Google Scholar]
- Sendin, L.N.; Filippone, M.P. The Genetic Transformation of Sweet Orange (Citrus sinensis L. Osbeck) for Enhanced Resistance to Citrus Canker. In Transgenic Plants; Humana Press: New York, NY, USA, 2019. [Google Scholar]
- Durán-Vila, N.; Ortega, V.; Navarro, L. Morphogenesis and tissue cultures of three citrus species. Plant Cell Tissue Organ Cult. 1989, 133, 123–133. [Google Scholar] [CrossRef]
- Peña, L.; Cervera, M.; Juárez, J.; Navarro, A.; Pina, J.A.; Durán-Vila, N.; Navarro, L. Agrobacterium-mediated transformation of sweet orange and regeneration of transgenic plants. Plant Cell Rep. 1995, 14, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Marques, N.T.; Nolasco, G.B.; Leitão, J.P. Factors affecting in vitro adventitious shoot formation on internode explants of Citrus aurantium L. cv. Brazilian. Sci. Hortic. 2011, 129, 176–182. [Google Scholar] [CrossRef]
- Birch, R.G. Plant Transformation: Problems and Strategies for Practical Application. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 297–326. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Cervera, M.; Juarez, J.; Navarro, A.; Ortega, C.; Pina, J.; Duran-Vila, N.; Navarro, L. Procedure for the genetic transformation of adult plants of woody species. Office 1998, 42, 1–47. [Google Scholar]
- Ghorbel, R.; la Malfa, S.; López, M.M.; Petit, A.; Navarro, L.; Peña, L. Additional copies of virG from pTiBo542 provide a super-transformation ability to Agrobacterium tumefaciens in citrus. Physiol. Mol. Plant Pathol. 2001, 58, 103–110. [Google Scholar] [CrossRef]
- Wu, H.; Acanda, Y.; Jia, H.; Wang, N.; Zale, J. Biolistic transformation of Carrizo citrange (Citrus sinensis Osb. × Poncirus trifoliata L. Raf.). Plant Cell Rep. 2016, 35, 1955–1962. [Google Scholar] [CrossRef]
- Wu, H.; Acanda, Y.; Canton, M.; Zale, J. Efficient Biolistic Transformation of Immature Citrus Rootstock Using Phosphomanose- isomerase Selection. Plants 2019, 8, 390. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhang, D.M.; Zhong, Y.; Chang, X.J.; Hu, M.L.; Cheng, C.Z. A simple and efficient in planta transformation method for pommelo (Citrus maxima) using Agrobacterium tumefaciens. Sci. Hortic. 2017, 214, 174–179. [Google Scholar] [CrossRef]
- Yong, H.; Yongrui, H.; Shanchun, C.; Yixiong, T.; Jinren, Z.; Shirong, J. New gene transformation technique forcommercial citrus cultivars. Chin. J. Trop. Crops 2000, 15, 37–41. [Google Scholar]
- Figueiredo, J.F.L.; Römer, P.; Lahaye, T.; Graham, J.H.; White, F.F.; Jones, J.B. Agrobacterium-mediated transient expression in citrus leaves: A rapid tool for gene expression and functional gene assay. Plant Cell Rep. 2011, 30, 1339–1345. [Google Scholar] [CrossRef]
- Sendín, L.N.; Filippone, M.P.; Orce, I.G.; Rigano, L.; Enrique, R.; Peña, L.; Vojnov, A.A.; Marano, M.R.; Castagnaro, A.P. Transient expression of pepper Bs2 gene in Citrus limon as an approach to evaluate its utility for management of citrus canker disease. Plant Pathol. 2012, 61, 648–657. [Google Scholar] [CrossRef]
- Enrique, R.; Siciliano, F.; Favaro, M.A.; Gerhardt, N.; Roeschlin, R.; Rigano, L.; Sendin, L.; Castagnaro, A.; Vojnov, A.; Marano, M.R. Novel demonstration of RNAi in citrus reveals importance of citrus callose synthase in defence against Xanthomonas citri subsp. citri. Plant Biotechnol. J. 2011, 9, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.I.; Dai, S.M.; Deng, Z.N.; Li, D.Z.; Long, G.Y.; Na, L.I.; Li, Y.; Gentile, A. Evaluation of parameters affecting Agrobacterium-mediated transient expression in citrus. J. Integr. Agric. 2017, 16, 572–579. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9, e93806. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, N. Xcc-facilitated agroinfiltration of citrus leaves: A tool for rapid functional analysis of transgenes in citrus leaves. Plant Cell Rep. 2014, 33, 1993–2001. [Google Scholar] [CrossRef]
- Jia, H.; Orbović, V.; Jones, J.B.; Wang, N. Modification of the PthA4 effector binding elements in Type I CsLOB1 promoter using Cas9/sgRNA to produce transgenic Duncan grapefruit alleviating XccΔpthA4: DCsLOB1.3 infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef]
- Jia, H.; Orbovi, V.; Wang, N. CRISPR-LbCas12a-mediated modification of citrus. Plant Biotechnol. J. 2019, 17, 1928–1937. [Google Scholar] [CrossRef]
- Peña, L.; Cervera, M.; Fagoaga, C.; Romero, J.; Ballester, A.; Soler, N.; Pons, E.; Rodriguez, A.; Peris, J.; Juarez, J.; et al. Citrus. In Compendium of Transgenic Crop Plants: Tropical and Subtropical Fruits and Nuts; Kole, C., Hall, T.C., Eds.; Blackwell Publishing: Oxford, UK, 2008; pp. 1–62. [Google Scholar]
- Ballester, A.; Cervera, M.; Peña, L. Selectable marker-free transgenic orange plants recovered under non-selective conditions and through PCR analysis of all regenerants. Plant Cell Tissue Organ Cult. 2010, 102, 329–336. [Google Scholar] [CrossRef]
- Ballester, A.; Cervera, M.; Peña, L. Evaluation of selection strategies alternative to nptII in genetic transformation of citrus. Plant Cell Rep. 2008, 27, 1005–1015. [Google Scholar] [CrossRef]
- Ballester, A.; Cervera, M.; Peña, L. Efficient production of transgenic citrus plants using isopentenyltransferase positive selection and removal of the marker gene by site-specific recombination. Plant Cell Rep. 2007, 738, 191–200. [Google Scholar] [CrossRef]
- Domínguez, A.; Fagoaga, C.; Navarro, L.; Moreno, P.; Peña, L. Regeneration of transgenic citrus plants under non selective conditions results in high-frequency recovery of plants with silenced transgenes. Mol. Genet. Genomics 2002, 267, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, A.; Cervera, M.; Pérez, R.M.; Romero, J.; Fagoaga, C.; Cubero, J.; López, M.M.; Juárez, J.A.; Navarro, L.; Peña, L. Characterisation of regenerants obtained under selective conditions after Agrobacterium-mediated transformation of citrus explants reveals production of silenced and chimeric plants at unexpected high frequencies. Mol. Breed. 2004, 14, 171–183. [Google Scholar] [CrossRef]
- Ghorbel, R.; Juárez, J.; Navarro, L.; Peña, L. Green fluorescent protein as a screenable marker to increase the efficiency of generating transgenic woody fruit plants. Theor. Appl. Genet. 1999, 99, 350–358. [Google Scholar] [CrossRef]
- Miles, J.S.; Guest, J.R. Nucleotide sequence and transcriptional start point of the phosphomannose isomerase gene (manA) of Escherichia coli. Gene 1984, 32, 41–48. [Google Scholar] [CrossRef]
- Joersbo, M.; Donaldson, L.; Kreiberg, J.; Petersen, S.G.; Brunstedt, J.; Okkels, F.T. Analysis of mannose selection used for transformation of sugar beet. Mol. Breed. 1998, 4, 111–117. [Google Scholar] [CrossRef]
- Ebinuma, H.; Sugita, K.; Matsunaga, E.; Yamakado, M. Selection of marker-free transgenic plants using the isopentenyl transferase gene. Proc. Natl. Acad. Sci. USA 1997, 94, 2117–2121. [Google Scholar] [CrossRef]
- Sugita, K.; Matsunaga, E.; Ebinuma, H. Effective selection system for generating marker-free transgenic plants independent of sexual crossing. Plant Cell Rep. 1999, 18, 941–947. [Google Scholar] [CrossRef]
- Endo, S.; Kasahara, T.; Sugita, K.; Ebinuma, H. A new GST-MAT vector containing both ipt and iaaM/H genes can produce marker-free transgenic tobacco plants with high frequency. Plant Cell Rep. 2002, 20, 923–928. [Google Scholar] [CrossRef]
- Lyznik, L.A.; Gordon-Kamm, W.J.; Tao, Y. Site-specific recombination for genetic engineering in plants. Plant Cell Rep. 2003, 21, 925–932. [Google Scholar] [CrossRef]
- Dale, E.C.; Ow, D.W. Gene transfer with subsequent removal of the selection gene from the host genome. Proc. Natl. Acad. Sci. USA 1991, 88, 10558–10562. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh, S.; Motallebi, M.; Zamani, M.R.; Jahromi, Z.M. Selectable marker gene removal and expression of transgene by inducible promoter containing FFDD Cis-acting elements in transgenic plants. Iran. J. Biotechnol. 2015, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moravčíková, J.; Vaculková, E.; Bauer, M.; Libantová, J. Feasibility of the seed specific cruciferin C promoter in the self excision Cre/loxP strategy focused on generation of marker-free transgenic plants. Theor. Appl. Genet. 2008, 117, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Boszorádová, E.; Libantová, J.; Matušíková, I.; Moravčíková, J. Application of Arabidopsis tissue-specific CRUC promoter in the Cre/loxP self-excision strategy for generation of marker-free oilseed rape: Potential advantages and drawbacks. Acta Physiol. Plant. 2014, 36, 1399–1409. [Google Scholar] [CrossRef]
- Kopertekh, L.; Schulze, K.; Frolov, A.; Strack, D.; Broer, I.; Schiemann, J. Cre-mediated seed-specific transgene excision in tobacco. Plant Mol. Biol. 2010, 72, 597–605. [Google Scholar] [CrossRef]
- van Ex, F.; Verweire, D.; Claeys, M.; Depicker, A.; Angenon, G. Evaluation of seven promoters to achieve germline directed Cre-lox recombination in Arabidopsis thaliana. Plant Cell Rep. 2009, 28, 1509–1520. [Google Scholar] [CrossRef]
- Dalla Costa, L.; Piazza, S.; Campa, M.; Flachowsky, H.; Hanke, M.V.; Malnoy, M. Efficient heat-shock removal of the selectable marker gene in genetically modified grapevine. Plant Cell Tissue Organ Cult. 2016, 124, 471–481. [Google Scholar] [CrossRef]
- Cuellar, W.; Gaudin, A.; Solórzano, D.; Casas, A.; Ñopo, L.; Chudalayandi, P.; Medrano, G.; Kreuze, J.; Ghislain, M. Self-excision of the antibiotic resistance gene nptII using a heat inducible Cre-loxP system from transgenic potato. Plant Mol. Biol. 2006, 62, 71–82. [Google Scholar] [CrossRef]
- Chong-Pérez, B.; Kosky, R.G.; Reyes, M.; Rojas, L.; Ocaña, B.; Tejeda, M.; Pérez, B.; Angenon, G. Heat shock induced excision of selectable marker genes in transgenic banana by the Cre-lox site-specific recombination system. J. Biotechnol. 2012, 159, 265–273. [Google Scholar] [CrossRef]
- Odell, J.T.; Nagy, F.; Chua, N.H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 1985, 313, 810–812. [Google Scholar] [CrossRef]
- Meyer, P.; Saedler, H. Homology-Dependent Gene Silencing in Plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 23–48. [Google Scholar] [CrossRef] [PubMed]
- Erpen, L.; Tavano, E.C.R.; Harakava, R.; Dutt, M.; Grosser, J.W.; Piedade, S.M.S.; Mendes, B.M.J.; Mourão Filho, F.A.A. Isolation, characterization, and evaluation of three Citrus sinensis-derived constitutive gene promoters. Plant Cell Rep. 2018, 37, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Erpen Dalla Corte, L.; Mendes, B.M.J.; Mourão Filho, F.A.A.; Grosser, J.W.; Dutt, M. Functional characterization of full-length and 5′ deletion fragments of Citrus sinensis-derived constitutive promoters in Nicotiana benthamiana. Vitr. Cell. Dev. Biol. Plant 2020. [Google Scholar] [CrossRef]
- Endo, T.; Shimada, T.; Fujii, H.; Moriguchi, T.; Omura, M. Promoter analysis of a type 3 metallothionein-like gene abundant in Satsuma mandarin (Citrus unshiu Marc.) fruit. Sci. Hortic. 2007, 112, 207–214. [Google Scholar] [CrossRef]
- Sorkina, A.; Bardosh, G.; Liu, Y.Z.; Fridman, I.; Schlizerman, L.; Zur, N.; Or, E.; Goldschmidt, E.E.; Blumwald, E.; Sadka, A. Isolation of a citrus promoter specific for reproductive organs and its functional analysis in isolated juice sacs and tomato. Plant Cell Rep. 2011, 30, 1627–1640. [Google Scholar] [CrossRef]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Omura, M. Isolation and characterization of a Citrus FT/TFL1 homologue (CuMFT1), which shows quantitatively preferential expression in Citrus seeds. J. Jpn. Soc. Hortic. Sci. 2008, 77, 38–46. [Google Scholar] [CrossRef]
- de Azevedo, F.A.; Mourão Filho, F.D.A.A.; Schinor, E.H.; de Paoli, L.G.; Mendes, B.M.J.; Harakava, R.; Gabriel, D.W.; Lee, R.F. GUS gene expression driven by a citrus promoter in transgenic tobacco and “Valencia” sweet orange. Pesqui. Agropecu. Bras. 2006, 41, 1623–1628. [Google Scholar] [CrossRef]
- Dutt, M.; Ananthakrishnan, G.; Jaromin, M.K.; Brlansky, R.H.; Grosser, J.W. Evaluation of four phloem-specific promoters in vegetative tissues of transgenic citrus plants. Tree Physiol. 2012, 32, 83–93. [Google Scholar] [CrossRef]
- Bhattacharyya-Pakrasi, M.; Peng, J.; Elmer, J.S.; Laco, G.; Shen, P.; Kaniewska, M.B.; Kononowicz, H.; Wen, F.; Hodges, T.K.; Beachy, R.N. Specificity of a promoter from the rice tungro bacilliform virus for expression in phloem tissues. Plant J. 1993, 4, 71–79. [Google Scholar] [CrossRef]
- Schmulling, T.; Schell, J.; Spena, A. Promoters of the rolA, B and C Genes of Agrobacterium rhizogenes Are Differentially Regulated in Transgenic Plants. Plant Cell 1989, 1, 665–670. [Google Scholar] [CrossRef]
- Sauer, N.; Stolz, J. SUC1 and SUC2: Two sucrose transporters from Arabidopsis thaliana; expression and characterization in baker’s yeast and identification of the histidine-tagged protein. Plant J. 1994, 6, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, M.B.; Powell, K.S.; van Damme, E.; Hilder, V.A.; Gatehouse, A.M.R.; Boulter, D.; Gatehouse, J.A. Use of the rice sucrose synthase-1 promoter to direct phloem-specific expression of β-glucuronidase and snowdrop lectin genes in transgenic tobacco plants. J. Exp. Bot. 1994, 45, 623–631. [Google Scholar] [CrossRef]
- Miyata, L.Y.; Harakava, R.; Stipp, L.C.L.; Mendes, B.M.J.; Appezzato-da-Glória, B.; de Assis Alves Mourão Filho, F. GUS expression in sweet oranges (Citrus sinensis L. Osbeck) driven by three different phloem-specific promoters. Plant Cell Rep. 2012, 31, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Tavano, E.C.D.R.; Erpen, L.; Aluisi, B.; Harakava, R.; Lopes, J.R.S.; Vieira, M.L.C.; Piedade, S.M.D.S.; Mendes, B.M.J.; Mourão Filho, F.D.A.A. Sweet orange genetic transformation with the attacin A gene under the control of phloem-specific promoters and inoculation with Candidatus Liberibacter asiaticus. J. Hortic. Sci. Biotechnol. 2019, 94, 210–219. [Google Scholar] [CrossRef]
- Zou, X.; Song, E.; Peng, A.; He, Y.; Xu, L.; Lei, T.; Yao, L.; Chen, S. Activation of three pathogen-inducible promoters in transgenic citrus (Citrus sinensis Osbeck) after Xanthomonas axonopodis pv. citri infection and wounding. Plant Cell Tissue Organ Cult. 2014, 117, 85–98. [Google Scholar] [CrossRef]
- Peng, J.L.; Bao, Z.L.; Li, P.; Chen, G.Y.; Wang, J.S.; Dong, H.S. HarpinX00 and its functional domains activate pathogen-inducible plant promoters in Arabidopsis. Acta Bot. Sin. 2004, 46, 1083–1090. [Google Scholar]
- Pontier, D.; Godiard, L.; Marco, Y.; Roby, D. hsr203J, a tobacco gene whose activation is rapid, highly localized and specific for incompatible plant/pathogen interactions. Plant J. 1994, 5, 507–521. [Google Scholar] [CrossRef]
- Malnoy, M.; Reynoird, J.P.; Borejsza-Wysocka, E.E.; Aldwinckle, H.S. Activation of the pathogen-inducible Gst1 promoter of potato after elicitation by Venturia inaequalis and Erwinia amylovora in transgenic apple (Malus × domestica). Transgenic Res. 2006, 15, 83–93. [Google Scholar] [CrossRef]
- Barbosa-Mendes, J.M.; Mourão Filho, F.D.A.A.; Filho, A.B.; Harakava, R.; Beer, S.V.; Mendes, B.M.J. Genetic transformation of Citrus sinensis cv. Hamlin with hrpN gene from Erwinia amylovora and evaluation of the transgenic lines for resistance to citrus canker. Sci. Hortic. 2009, 122, 109–115. [Google Scholar] [CrossRef]
- Sendin, L.N.; Orce, I.G.; Gómez, R.L.; Enrique, R.; Grellet Bournonville, C.F.; Noguera, A.S.; Vojnov, A.A.; Marano, M.R.; Castagnaro, A.P.; Filippone, M.P. Inducible expression of Bs2 R gene from Capsicum chacoense in sweet orange (Citrus sinensis L. Osbeck) confers enhanced resistance to citrus canker disease. Plant Mol. Biol. 2017, 93, 607–621. [Google Scholar] [CrossRef]
- Cernadas, R.A.; Camillo, L.R.; Benedetti, C.E. Transcriptional analysis of the sweet orange interaction with the citrus canker pathogens Xanthomonas axonopodis pv. citri and Xanthomonas axonopodis pv. aurantifolii. Mol. Plant Pathol. 2008, 9, 609–631. [Google Scholar] [CrossRef]
- de Oliveira, M.L.; de Lima Silva, C.C.; Abe, V.Y.; Costa, M.G.C.; Cernaddas, R.A.; Benedetti, C.E. Increased resistance against citrus canker mediated by a citrus mitogen-activated protein kinase. Mol. Plant-Microbe Interact. 2013, 26, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Dutt, M.; Dhekney, S.A.; Soriano, L.; Kandel, R.; Grosser, J.W. Temporal and spatial control of gene expression in horticultural crops. Hortic. Res. 2014, 1, 1–17. [Google Scholar]
- Duan, Y.X.; Fan, J.; Guo, W.W. Regeneration and characterization of transgenic kumquat plants containing the Arabidopsis APETALA1 gene. Plant Cell Tissue Organ Cult. 2010, 100, 273–281. [Google Scholar] [CrossRef]
- Tan, B.; Li, D.L.; Xu, S.X.; Fan, G.E.; Fan, J.; Guo, W.W. Highly efficient transformation of the GFP and MAC12.2 genes into precocious trifoliate orange (Poncirus trifoliata [L.] Raf), a potential model genotype for functional genomics studies in Citrus. Tree Genet. Genomes 2009, 5, 529–537. [Google Scholar] [CrossRef]
- Yang, L.; Hu, W.; Xie, Y.; Li, Y.; Deng, Z. Factors affecting Agrobacterium-mediated transformation efficiency of kumquat seedling internodal stem segments. Sci. Hortic. 2016, 209, 105–112. [Google Scholar] [CrossRef]
- Zhu, C.; Zheng, X.; Huang, Y.; Ye, J.; Chen, P.; Zhang, C.; Zhao, F.; Xie, Z.; Zhang, S.; Wang, N.; et al. Genome sequencing and CRISPR/Cas9 gene editing of an early flowering Mini-Citrus (Fortunella hindsii). Plant Biotechnol. J. 2019, 17, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Martín-Trillo, M.; Juárez, J.; Pina, J.A.; Navarro, L.; Martínez-Zapater, J.M. Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time. Nat. Biotechnol. 2001, 19, 263–267. [Google Scholar] [CrossRef]
- Cervera, M.; Navarro, L.; Peña, L. Gene stacking in 1-year-cycling APETALA1 citrus plants for a rapid evaluation of transgenic traits in reproductive tissues. J. Biotechnol. 2009, 140, 278–282. [Google Scholar] [CrossRef]
- Endo, T.; Shimada, T.; Fujii, H.; Kobayashi, Y.; Araki, T.; Omura, M. Ectopic expression of an FT homolog from Citrus confers an early flowering phenotype on trifoliate orange (Poncirus trifoliata L. Raf.). Transgenic Res. 2005, 14, 703–712. [Google Scholar] [CrossRef]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Kobayashi, Y.; Araki, T.; Omura, M. Transcriptional changes in CiFT-introduced transgenic trifoliate orange (Poncirus trifoliata L. Raf.). Tree Physiol. 2010, 30, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Pons, E.; Alquézar, B.; Rodríguez, A.; Martorell, P.; Genovés, S.; Ramón, D.; Rodrigo, M.J.; Zacarías, L.; Peña, L. Metabolic engineering of β-carotene in orange fruit increases its in vivo antioxidant properties. Plant Biotechnol. J. 2014, 12, 17–27. [Google Scholar] [CrossRef]
- Velázquez, K.; Agüero, J.; Vives, M.C.; Aleza, P.; Pina, J.A.; Moreno, P.; Navarro, L.; Guerri, J. Precocious flowering of juvenile citrus induced by a viral vector based on Citrus leaf blotch virus: A new tool for genetics and breeding. Plant Biotechnol. J. 2016, 14, 1976–1985. [Google Scholar] [CrossRef] [PubMed]
- von Aderkas, P.; Bonga, J.M. Influencing micropropagation and somatic embryogenesis in mature trees by manipulation of phase change, stress and culture environment. Tree Physiol. 2000, 20, 921–928. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, S.; Peng, A.; Zou, X.; Xu, L.; Lei, T.; Liu, X.; Yao, L. Production and evaluation of transgenic sweet orange (Citrus sinensis Osbeck) containing bivalent antibacterial peptide genes (Shiva A and Cecropin B) via a novel Agrobacterium-mediated transformation of mature axillary buds. Sci. Hortic. 2011, 128, 99–107. [Google Scholar] [CrossRef]
- Gentile, A.; Deng, Z.N.; la Malfa, S.; Domina, F.; Germanà, C.; Tribulato, E. Morphological and physiological effects of rolABC genes into citrus genome. Acta Hortic. 2004, 632, 235–242. [Google Scholar] [CrossRef]
- Distefano, G.; Cirilli, M.; las Casas, G.; la Malfa, S.; Continella, A.; Rugini, E.; Thomas, B.; Long, G.; Gentile, A.; Muleo, R. Ectopic expression of Arabidopsis phytochrome B in Troyer citrange affects photosynthesis and plant morphology. Sci. Hortic. 2013, 159, 1–7. [Google Scholar] [CrossRef]
- Fagoaga, C.; Tadeo, F.R.; Iglesias, D.J.; Huerta, L.; Lliso, I.; Vidal, A.M.; Talon, M.; Navarro, L.; García-Martínez, J.L.; Peña, L. Engineering of gibberellin levels in citrus by sense and antisense overexpression of a GA 20-oxidase gene modifies plant architecture. J. Exp. Bot. 2007, 58, 1407–1420. [Google Scholar] [CrossRef]
- Cervera, M.; Ortega, C.; Navarro, A.; Navarro, L.; Peña, L. Generation of transgenic citrus plants with the tolerance-to-salinity gene HAL2 from yeast. J. Hortic. Sci. Biotechnol. 2000, 75, 26–30. [Google Scholar] [CrossRef]
- Molinari, H.B.C.; Marur, C.J.; Filho, J.C.B.; Kobayashi, A.K.; Pileggi, M.; Júnior, R.P.L.; Pereira, L.F.P.; Vieira, L.G.E. Osmotic adjustment in transgenic citrus rootstock Carrizo citrange (Citrus sinensis Osb. × Poncirus trifoliata L. Raf.) overproducing proline. Plant Sci. 2004, 167, 1375–1381. [Google Scholar] [CrossRef]
- Orbović, V.; Ćalović, M.; Dutt, M.; Grosser, J.W.; Barthe, G. Production and characterization of transgenic Citrus plants carrying p35 anti-apoptotic gene. Sci. Hortic. 2015, 197, 203–211. [Google Scholar] [CrossRef]
- Orbović, V.; Fields, J.S.; Syvertsen, J.P. Transgenic citrus plants expressing the p35 anti-apoptotic gene have altered response to abiotic stress. Hortic. Environ. Biotechnol. 2017, 58, 303–309. [Google Scholar] [CrossRef]
- Dutt, M.; Stanton, D.; Grosser, J.W. Ornacitrus: Development of genetically modified anthocyanin-expressing citrus with both ornamental and fresh fruit potential. J. Am. Soc. Hortic. Sci. 2016, 141, 54–61. [Google Scholar] [CrossRef]
- Hijaz, F.; Nehela, Y.; Jones, S.E.; Dutt, M.; Grosser, J.W.; Manthey, J.A.; Killiny, N. Metabolically engineered anthocyanin-producing lime provides additional nutritional value and antioxidant potential to juice. Plant Biotechnol. Rep. 2018, 12, 329–346. [Google Scholar] [CrossRef]
- Alquézar, B.; Rodrigo, M.J.; Zacarías, L. Regulation of carotenoid biosynthesis during fruit maturation in the red-fleshed orange mutant Cara Cara. Phytochemistry 2008, 69, 1997–2007. [Google Scholar] [CrossRef]
- Li, D.D.; Shi, W.; Deng, X.X. Factors influencing Agrobacterium-mediated embryogenic callus transformation of Valencia sweet orange (Citrus sinensis) containing the pTA29-barnase gene. Tree Physiol. 2003, 23, 1209–1215. [Google Scholar] [CrossRef]
- Koltunow, A.M.; Brennan, P.; Protopsaltis, S. Regeneration of West Indian Limes (Citrus aurantifolia) containing genes for decreased seed set. Acta Hortic. 2000, 535, 81–91. [Google Scholar] [CrossRef]
- Hudson, M.E. Florida Citrus Statistics 2017–2018; Florida Department of Agricultural and Consume Services; United States Department of Agriculture: Washington, DC, USA, 2019; pp. 1–114.
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- Coletta-filho, H.D.; Daugherty, M.P.; Ferreira, C.; Lopes, J.R.S. Temporal Progression of ‘Candidatus Liberibacter asiaticus’ Infection in Citrus and Acquisition Efficiency by Diaphorina citri Helvecio. Phytopathology 2014, 104, 416–421. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Bassanezi, R.B.; Paulo, S. Citrus Huanglongbing: The Pathogen and Its Impact Plant Health Progress Plant Health Progress. Plant Health Prog. 2007, 8, 31. [Google Scholar] [CrossRef]
- Boman, H.G. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Schaad, N.W.; Postnikova, E.; Lacy, G.; Sechler, A.; Agarkova, I.; Stromberg, P.E.; Stromberg, V.K.; Vidaver, A.K. Emended classification of xanthomonad pathogens on citrus. Syst. Appl. Microbiol. 2006, 29, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Soriano, L.; Tavano, E.C.D.R.; Correa, M.F.; Harakava, R.; Mendes, B.M.J.; Mourão Filho, F.D.A.A. In vitro organogenesis and genetic transformation of mandarin cultivars. Revista Brasileira de Fruticultura 2019, 41, 1–11. [Google Scholar] [CrossRef]
- Zou, X.; Jiang, X.; Xu, L.; Lei, T.; Peng, A.; He, Y.; Yao, L.; Chen, S. Transgenic citrus expressing synthesized cecropin B genes in the phloem exhibits decreased susceptibility to Huanglongbing. Plant Mol. Biol. 2017, 93, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Stover, E.; Gupta, G. Overexpression of a modified plant Thionin enhances disease resistance to Citrus canker and Huanglongbing (HLB). Front. Plant Sci. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Furman, N.; Kobayashi, K.; Zanek, M.C.; Calcagno, J.; García, M.L.; Mentaberry, A. Transgenic sweet orange plants expressing a dermaseptin coding sequence show reduced symptoms of citrus canker disease. J. Biotechnol. 2013, 167, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. Current status of the gene-for-gene concept. Ann. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Mendes, B.M.J.; Cardoso, S.C.; Boscariol-Camargo, R.L.; Cruz, R.B.; Mourão Filho, F.A.A.; Bergamin Filho, A. Reduction in susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis expressing the rice Xa21 gene. Plant Pathol. 2010, 59, 68–75. [Google Scholar] [CrossRef]
- Omar, A.A.; Murata, M.M.; El-Shamy, H.A.; Graham, J.H.; Grosser, J.W. Enhanced resistance to citrus canker in transgenic mandarin expressing Xa21 from rice. Transgenic Res. 2018, 27, 179–191. [Google Scholar] [CrossRef]
- Hao, G.; Pitino, M.; Duan, Y.; Stover, E. Reduced susceptibility to xanthomonas citri in transgenic citrus expressing the FLS2 receptor from nicotiana benthamiana. Mol. Plant-Microbe Interact. 2016, 29, 132–142. [Google Scholar] [CrossRef]
- Kuc, J. Immunity to Plant Disease. BioScience 1982, 32, 854–860. [Google Scholar]
- Chen, X.; Barnaby, J.Y.; Sreedharan, A.; Huang, X.; Orbović, V.; Grosser, J.W.; Wang, N.; Dong, X.; Song, W.Y. Over-expression of the citrus gene CtNH1 confers resistance to bacterial canker disease. Physiol. Mol. Plant Pathol. 2013, 84, 115–122. [Google Scholar] [CrossRef]
- Boscariol-Camargo, R.L.; Takita, M.A.; Machado, M.A. Bacterial resistance in AtNPR1 transgenic sweet orange is mediated by priming and involves EDS1 and PR2. Trop. Plant Pathol. 2016, 41, 341–349. [Google Scholar] [CrossRef]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. Eur. J. Plant Pathol. 2010, 128, 91–100. [Google Scholar] [CrossRef]
- Dutt, M.; Barthe, G.; Irey, M.; Grosser, J. Transgenic citrus expressing an arabidopsis NPR1 gene exhibit enhanced resistance against Huanglongbing (HLB.; Citrus greening). PLoS ONE 2015, 10, e0137134. [Google Scholar] [CrossRef]
- Robertson, C.J.; Zhang, X.; Gowda, S.; Orbović, V.; Dawson, W.O.; Mou, Z. Overexpression of the Arabidopsis NPR1 protein in citrus confers tolerance to Huanglongbing. J. Citrus Pathol. 2018, 5, 1–8. [Google Scholar]
- Qiu, W.; Soares, J.; Pang, Z.; Huang, Y.; Sun, Z.; Wang, N.; Grosser, J.; Dutt, M. Potential Mechanisms of AtNPR1 Mediated Resistance against Huanglongbing (HLB) in Citrus. Int. J. Mol. Sci. 2020, 21, 2009. [Google Scholar] [CrossRef]
- Gentile, A.; Deng, Z.; la Malfa, S.; Distefano, G.; Domina, F.; Vitale, A.; Polizzi, G.; Lorito, M.; Tribulato, E. Enhanced resistance to Phoma tracheiphila and Botrytis cinerea in transgenic lemon plants expressing a Trichoderma harzianum chitinase gene. Plant Breed. 2007, 126, 146–151. [Google Scholar] [CrossRef]
- Distefano, G.; la Malfa, S.; Vitale, A.; Lorito, M.; Deng, Z.; Gentile, A. Defence-related gene expression in transgenic lemon plants producing an antimicrobial Trichoderma harzianum endochitinase during fungal infection. Transgenic Res. 2008, 17, 873–879. [Google Scholar] [CrossRef]
- Muccilli, V.; Vitale, A.; Sheng, L.; Gentile, A.; Cardullo, N.; Tringali, C.; Oliveri, C.; la Rosa, R.; di Guardo, M.; la Malfa, S.; et al. Substantial Equivalence of a Transgenic Lemon Fruit Showing Postharvest Fungal Pathogens Resistance. J. Agric. Food Chem. 2020, 68, 3806–3816. [Google Scholar] [CrossRef]
- Domínguez, A.; de Mendoza, A.H.; Guerri, J.; Cambra, M.; Navarro, L.; Moreno, P.; Peña, L. Pathogen-derived resistance to Citrus tristeza virus (CTV) in transgenic mexican lime (Citrus arrurantifolia (Christ.) Swing.) plants expressing its p25 coat protein gene. Mol. Breed. 2002, 10, 1–10. [Google Scholar] [CrossRef]
- Fagoaga, C.; López, C.; de Mendoza, A.H.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Post-transcriptional gene silencing of the p23 silencing suppressor of Citrus tristeza virus confers resistance to the virus in transgenic Mexican lime. Plant Mol. Biol. 2006, 60, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Soler, N.; Plomer, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnol. J. 2012, 10, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Zhang, Y.; Yang, J.; Zhong, Y. Expression of hairpin RNA (hpRNA) targeting the three CTV-silencing suppressor genes confers sweet orange with stem-pitting CTV tolerance. J. Hortic. Sci. Biotechnol. 2017, 92, 465–474. [Google Scholar] [CrossRef]
- Reyes, C.A.; de Francesco, A.; Peña, E.J.; Costa, N.; Plata, M.I.; Sendin, L.; Castagnaro, A.P.; García, M.L. Resistance to Citrus psorosis virus in transgenic sweet orange plants is triggered by coat protein-RNA silencing. J. Biotechnol. 2011, 151, 151–158. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Masunaka, A.; Tsuge, T.; Yamamoto, M.; Ohtani, K.; Fukumoto, T.; Gomi, K.; Peever, T.L.; Akimitsu, K. Functional analysis of a multicopy host-selective ACT-toxin biosynthesis gene in the tangerine pathotype of Alternaria alternata using RNA silencing. Mol. Plant-Microbe Interact. 2008, 21, 1591–1599. [Google Scholar] [CrossRef]
- Narayan, R.D.; Blackman, L.M.; Shan, W.; Hardham, A.R. Phytophthora nicotianae transformants lacking dynein light chain 1 produce non-flagellate zoospores. Fungal Genet. Biol. 2010, 47, 663–671. [Google Scholar] [CrossRef]
- Singerman, A.; Rogers, M.E. The Economic Challenges of Dealing with Citrus Greening: The Case of Florida. J. Integr. Pest Manag. 2020, 11. [Google Scholar] [CrossRef]
- Paiva, P.E.B.; Cota, T.; Neto, L.; Soares, C.; Tomás, J.C.; Duarte, A. Water Vapor Pressure Deficit in Portugal and Implications for the Development of the Invasive African Citrus Psyllid Trioza erytreae. Insects 2020, 11, 229. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for genome editing, regulation and targeting. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Malzahn, A.A.; Sretenovic, S.; Qi, Y. The emerging and uncultivated potential of CRISPR technology in plant science. Nat. Plants 2019, 5, 778–794. [Google Scholar] [CrossRef] [PubMed]
- Agüero, J.; del Carmen Vives, M.; Velázquez, K.; Pina, J.A.; Navarro, L.; Moreno, P.; Guerri, J. Effectiveness of gene silencing induced by viral vectors based on Citrus leaf blotch virus is different in Nicotiana benthamiana and citrus plants. Virology 2014, 460–461, 154–164. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.B.; Hao, B.H.; Lyon, M.P.; et al. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, J.; Jia, H.; Sosso, D.; Li, T.; Frommer, W.B.; Yang, B.; White, F.F.; Wang, N.; Jones, J.B. Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc. Natl. Acad. Sci. USA 2014, 111. [Google Scholar] [CrossRef]
- Yang, L.; Hu, C.; Li, N.; Zhang, J.; Yan, J.; Deng, Z. Transformation of sweet orange [Citrus sinensis (L.) Osbeck] with pthA-nls for acquiring resistance to citrus canker disease. Plant Mol. Biol. 2011, 75, 11–23. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef]
- Zaidi, S.S.E.A.; Mahfouz, M.M.; Mansoor, S. CRISPR-Cpf1: A New Tool for Plant Genome Editing. Trends Plant Sci. 2017, 22, 550–553. [Google Scholar] [CrossRef]
- Endo, A.; Masafumi, M.; Kaya, H.; Toki, S. Efficient targeted mutagenesis of rice and tobacco genomes using Cpf1 from Francisella novicida. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, C.; Liu, Q.; Fu, Y.; Wang, K. Targeted mutagenesis in rice using CRISPR-Cpf1 system. J. Genet. Genomics 2017, 44, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Qin, R.; Li, H.; Li, D.; Li, L.; Wei, P.; Yang, J. Generation of targeted mutant rice using a CRISPR-Cpf1 system. Plant Biotechnol. J. 2017, 15, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Biswal, A.K.; Dionora, J.; Perdigon, K.M.; Balahadia, C.P.; Mazumdar, S.; Chater, C.; Lin, H.C.; Coe, R.A.; Kretzschmar, T.; et al. CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep. 2017, 36, 745–757. [Google Scholar] [CrossRef]
- Tang, X.; Lowder, L.G.; Zhang, T.; Malzahn, A.A.; Zheng, X.; Voytas, D.F.; Zhong, Z.; Chen, Y.; Ren, Q.; Li, Q.; et al. A CRISPR-Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 2017, 3, 1–5. [Google Scholar] [CrossRef]
- Jia, H.; Zou, X.; Orbović, V.; Wang, N. Genome Editing in Citrus Tree with CRISPR/Cas9. In Plant Genome Editing with CRISPR Systems. Methods in Molecular Biology; Qi, Y., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1917, pp. 235–241. ISBN 9781493989911. [Google Scholar]
- Wang, L.; Chen, S.; Peng, A.; Xie, Z.; He, Y.; Zou, X. CRISPR/Cas9-mediated editing of CsWRKY22 reduces susceptibility to Xanthomonas citri subsp. citri in Wanjincheng orange (Citrus sinensis (L.) Osbeck). Plant Biotechnol. Rep. 2019, 13, 501–510. [Google Scholar] [CrossRef]
- Zhang, F.; LeBlanc, C.; Irish, V.F.; Jacob, Y. Rapid and efficient CRISPR/Cas9 gene editing in Citrus using the YAO promoter. Plant Cell Rep. 2017, 36, 1883–1887. [Google Scholar] [CrossRef]
- Yan, L.; Wei, S.; Wu, Y.; Hu, R.; Li, H.; Yang, W.; Xie, Q. High-Efficiency Genome Editing in Arabidopsis Using YAO Promoter-Driven CRISPR/Cas9 System. Mol. Plant 2015, 8, 1820–1823. [Google Scholar] [CrossRef]
- LeBlanc, C.; Zhang, F.; Mendez, J.; Lozano, Y.; Chatpar, K.; Irish, V.F.; Jacob, Y. Increased efficiency of targeted mutagenesis by CRISPR/Cas9 in plants using heat stress. Plant J. 2018, 93, 377–386. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Generation of homozygous canker-resistant citrus in the T0 generation using CRISPR-SpCas9p. Plant Biotechnol. J. 2020, 1–3. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.J.; Kim, S.; Kim, J.S.; Velasco, R.; Kanchiswamy, C.N. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Caserta, R.; Teixeira-Silva, N.S.; Granato, L.M.; Dorta, S.O.; Rodrigues, C.M.; Mitre, L.K.; Yochikawa, J.T.H.; Fischer, E.R.; Nascimento, C.A.; Souza-Neto, R.R.; et al. Citrus biotechnology: What has been done to improve disease resistance in such an important crop? Biotechnol. Res. Innov. 2020. [Google Scholar] [CrossRef]
- Kim, J.I.; Kim, J.Y. New era of precision plant breeding using genome editing. Plant Biotechnol. Rep. 2019, 13, 419–421. [Google Scholar] [CrossRef]


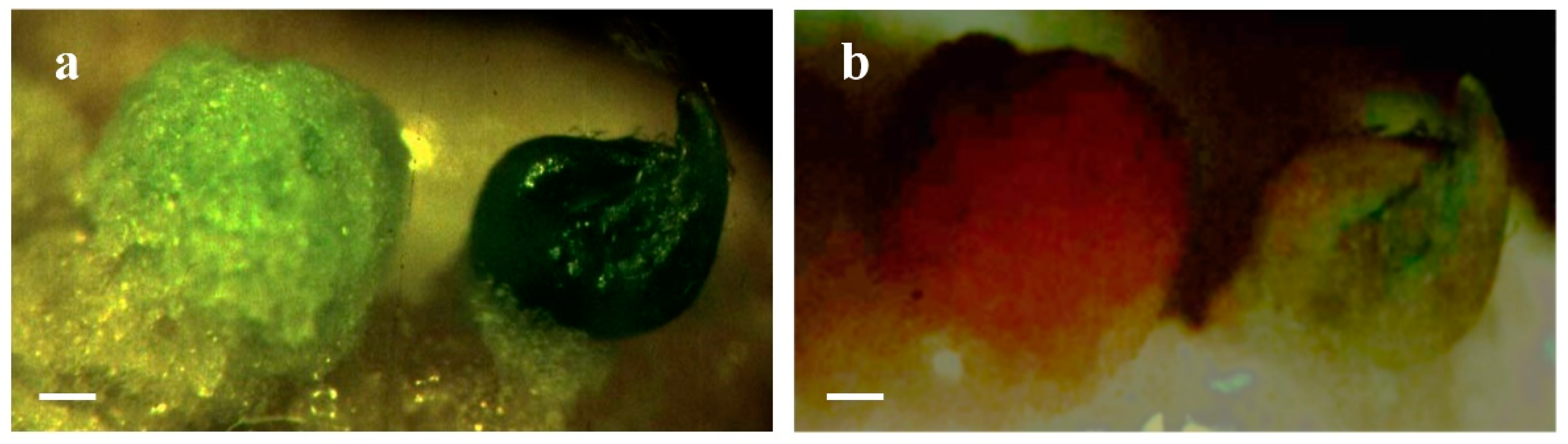{kind=link}
| Species | Cultivar/Genotype | Explant Type | RE (%) | TE (%) | Reference |
|---|---|---|---|---|---|
| C. sinensis L. Osb. × P. trifoliata L. Raf. | Carrizo citrange | ES | 37.5 | 20.6 | [19] |
| C. sinensis L. Osb. | Valencia | ES | 28.8 | 23.8 | [23] |
| C. sinensis L. Osb. | Valencia | MIS | 9.12 | 0.88 | [24] |
| C. sinensis L. Osb. | Tarocco | MNS | 74.7 | 9.1 | [25] |
| C. sinensis L. Osb. | Pineapple | MIS | 23 | 6.1 | [26] |
| C. sinensis L. Osb. | Jincheng | ES | 28.3 | 4.7 | [27] |
| Fortunella crassifolia Swingle | Jindan | ES | 13 | 3.6 | [28] |
| C. reticulata ‘Sunki’ × P. trifoliata ‘Flying Dragon’ | US-942 | MIS | 29.42 | 3.96 | [24] |
| C. clementina | Clemenules | MIS | 1.28 | 0.3-3 | [20] |
| C. paradisi Macf. | Ruby Red | MIS | 10.70 | 1.05 | [24] |
| C. medica L. | Etrog | MIS | 9.49 | 1.49 | [24] |
| P. trifoliata L. Raf. | Precocious trifoliate orange | ES | 66.1 | 57.4 | [29] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poles, L.; Licciardello, C.; Distefano, G.; Nicolosi, E.; Gentile, A.; La Malfa, S. Recent Advances of In Vitro Culture for the Application of New Breeding Techniques in Citrus. Plants 2020, 9, 938. https://doi.org/10.3390/plants9080938
Poles L, Licciardello C, Distefano G, Nicolosi E, Gentile A, La Malfa S. Recent Advances of In Vitro Culture for the Application of New Breeding Techniques in Citrus. Plants. 2020; 9(8):938. https://doi.org/10.3390/plants9080938
Chicago/Turabian StylePoles, Lara, Concetta Licciardello, Gaetano Distefano, Elisabetta Nicolosi, Alessandra Gentile, and Stefano La Malfa. 2020. "Recent Advances of In Vitro Culture for the Application of New Breeding Techniques in Citrus" Plants 9, no. 8: 938. https://doi.org/10.3390/plants9080938
APA StylePoles, L., Licciardello, C., Distefano, G., Nicolosi, E., Gentile, A., & La Malfa, S. (2020). Recent Advances of In Vitro Culture for the Application of New Breeding Techniques in Citrus. Plants, 9(8), 938. https://doi.org/10.3390/plants9080938